
Abstract
Glial scars formed after brain injuries provide permissive cues for endogenous neural precursor/stem cells (eNP/SCs) to undergo astrogenesis rather than neurogenesis. Following brain injury, eNP/SCs from the subventricular zone leave their niche, migrate to the injured cortex, and differentiate into reactive astrocytes that contribute to glial scar formation. In vivo neuronal reprogramming, directly converting non-neuronal cells such as reactive astrocytes or NG2 glia into neurons, has greatly improved brain injury repair strategies. However, reprogramming carries a high risk of future clinical applications such as tumorigenicity, involving virus. In this study, we constructed a neural matrix to alter the adverse niche at the injured cortex, enabling eNP/SCs to differentiate into functional neurons. We found that the neural matrix functioned as a “glial trap” that largely concentrated and limited reactive astrocytes to the core of the lesion area, thus altering the adverse niche. The eNP/SCs migrated toward the injured cortex and differentiated into functional neurons. In addition, regenerated neurites extended across the boundary of the injured cortex. Mice treated with the neural matrix demonstrated significant behavioral recovery. For the first time, we induced eNP/SC-derived functional neurons in the cortex after brain injury without the use of viruses, microRNAs, or small molecules. Our novel strategy of applying this “glial trap” to obtain functional neurons in the injured cortex may provide a safer and more natural therapeutic alternative to reprogramming in future clinical applications.
Introduction
Brain injury is often accompanied by neuron loss and scar formation [1 –3]. Neural precursor/stem cells (NP/SCs), administered by either direct intracerebral injection or systemic injection after brain injury, have been used to reduce neuron loss and improve neurological functional recovery [4 –6]. However, the effects of exogenous NP/SCs hinder the restoration of brain function, primarily due to the low survival rate of grafted cells in host tissues [7,8], which may be caused by mechanical damage, acute inflammation, immunological rejection, or a lack of trophic signals [9].
After brain injury, glial cells are activated and proliferate rapidly to become reactive glial cells in the injured cortex, forming a glial scar that prevents neuronal growth inside the brain [10,11]. In recent years, neuronal reprogramming by the conversion of non-neuronal cells into functional neurons has represented a substantial improvement in the field of repairing injured or diseased brains [12,13]. Previous studies have shown that postnatal and adult endogenous NP/SCs (eNP/SCs) located in the mammalian subventricular zone (SVZ) and subgranular zone can become mature neurons under suitable conditions [14,15]. However, SVZ-derived NP/SCs home in on the injured cortex to become reactive astrocytes, which contribute to astrogenesis after brain injury [16,17]. We constructed a neural matrix that functions as a “glial trap” to restrict the reactive astrocyte area, providing neurotrophic factors and inducing eNP/SCs to differentiate into functional neurons in the injured cortex for brain injury repair. Unlike reprogramming methods using additional neurogenic transcription factors (TFs), microRNAs, or small molecules, our novel approach restored functional neurons in the injured cortex in a moderate way without direct cellular interference.
Based on evidence that the SVZ NP/SC-generated neuroblasts move to the olfactory bulb (OB), where they differentiate into new neurons [18,19], we constructed a neural matrix from OB to mimic neural niche. Stromal cell-derived factor 1 (SDF-1) is a key regulator of directed cell migration after injury [20]. Brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF) have been reported to play important roles in promoting neurogenesis, by improving the survival of newly generated neurons, dendritic arborization, or synaptic formation [21,22]. A previous study showed that chondroitinase ABC (ChABC) ablated glial scars and thus augmented the growth of neurites and enhanced synaptic plasticity for injury repair [23,24]. We immobilized SDF-1, BDNF, NGF, and ChABC in OB-derived matrix by chemical conjugation to prevent their rapid diffusion into the cerebrospinal fluid and removal from the injured cortex. The constructed neural matrix functioned as a glial trap and formed a favorable neurogenic milieu for neurogenesis at the injured cortex following brain injury.
Experimental Model and Subject Details
Mice
Nestin-CreERT2 mice and R26R-RFP mice were crossed to obtain offspring to track adult NP/SCs in the SVZ. The procedures for obtaining Nestin-CreERT2 mice [25] and R26R-RFP mice [26] have been described previously. All animal procedures were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978) and approved by the Animal Ethics Committee of Fudan University. Tamoxifen (TAM) was dissolved in corn oil to 20 mg/mL. To track adult NP/SCs, 6–8-week-old mice were selected and TAM (0.15 mg/g) was introduced by oral gavage 14 days before brain injury.
Genomic polymerase chain reaction
Genomic DNA was extracted from the tails of the mice. Tissues were incubated and lysed in proteinase K overnight at 55°C, and then centrifuged at maximum speed (21,130 g) for 5 min to obtain supernatants with genomic DNA. The DNA was precipitated with isopropanol, washed in 70% ethanol, and dissolved in deionized water.
Preparation of neural matrix
After neonatal C57BL/6 mice were sacrificed, OBs were isolated and rinsed with phosphate-buffered saline (PBS). After eliminating blood vessels and meninges from the OB surface, the OBs were cut into sections. The OB sections were treated sequentially with demineralized water for 6 h, 4% sodium deoxycholate (Sigma) for 12 h, PBS for 20 min, 40 kU/mL DNase I (Sigma) for 60 min, PBS for 10 min, demineralized water for 4 h, 3% Triton X-100 for 2 h, PBS for 10 min, 40 kU/mL DNase for 60 min, and PBS for 15 min. The procedure was repeated in four cycles for decellularization. The sections were washed five times with demineralized water, and then homogenized in 0.05% (v/v) acetic acid for 3 days. After neutralization with 4 M NaOH, the matrix was dialyzed in demineralized water, which was then adjusted to a concentration of 20 mg/mL. The matrix was divided into four parts, and then each part was mixed with 3 mg/mL Traut's reagent (2-iminothiolane•HCl; Thermo Fisher Scientific) in PBS with 4 mM ethylenediaminetetraacetic acid (EDTA) (pH = 8) for 1 h. The four elements, 0.1 mg/mL BDNF, 0.1 mg/mL NGF, 0.25 mg/mL SDF-1, and 50 kU/mL ChABC, were then dissolved in PBS with 4 mM EDTA, and mixed with a 50-fold molar excess of sulfosuccinimidyl 4-[N-maleimidomethyl]cyclohexane-1-carboxylate (Sulfo-SMCC) (Thermo Fisher Scientific) for 30 min. Matrices that had been reacted with Traut's reagent were mixed with the sulfo-SMCC-reacted elements for 30 min. The four matrix parts were mixed in a blender to obtain the neural matrix.
Surgical procedures and matrix application
The brain injury surgical procedure was performed based on previous studies, with modifications [27]. Briefly, mice were anesthetized by intraperitoneal injection with sodium pentobarbital (40 mg/kg body weight) and randomized to three groups: the treatment group, control group, and sham group (n = 30 mice per group). The mice were mounted in a stereotactic frame, followed by craniotomy on the right parietal cortex. A 2.5-mm-diameter hole was created midway between the bregma and lambda sutures, adjacent to the central suture. A cylindrical polyacetal transducer rod with a 2.5-mm-diameter tip was then placed on the skull hole, and a weight was dropped from a height of 15 cm to vertically strike the transducer rod. In the treatment group, the injury cavity was filled with neural matrix. In the control group, the injury cavity was filled with PBS. In the sham group, mice were maintained under similar environmental conditions, but were not subjected to impact injury. Incisions were sutured with nylon sutures, and the mice were kept in cages under normal conditions with a 12-h light/12-h dark cycle and ad libitum food and water until sacrifice.
Immunofluorescence staining
Mice were perfused with 4% paraformaldehyde (PFA) to collect the right brain, which was then washed in PBS to remove excess blood. The brains were then immersed in 4% PFA for 90 min, washed with PBS, and sequentially cryoprotected in 20% sucrose and 30% sucrose in PBS. The brain was embedded in optimal cutting temperature compound and cut into fifty 15-μm cryosections per mouse on a cryostat. The cryosections were air-dried for 1 h at room temperature and then blocked with blocking buffer (4% donkey serum and 0.1% Triton X-100 in PBS) for 45 min. Primary antibodies were applied and the sections were incubated overnight at 4°C, followed by extensive washing with PBS to remove unbound primary antibodies. Colors were developed with secondary antibodies for 1 h at room temperature. Finally, the sections were washed three times in PBS and counterstained with DAPI (Vector Labs). Five random fields for each section with fluorescence reporters were observed and photographed on Olympus (FV1000 and FV1200) and Nikon (A1+ and C2+) confocal microscopes. Primary and secondary antibodies are listed in Table 1.
Antibodies
GFAP, glial fibrillary acidic protein; MAP2, microtubule-associated protein; Ms, mouse; Rb, rabbit; RFP, red fluorescent protein; VGluT1, vesicular glutamate transporter 1.
Scratch assay for cell migration
Human glial cell suspensions were seeded on 12-well plates at 5 × 105 cells per well. For each group, five replicate wells were set in parallel. At 24 h after seeding, a sterile micropipette tip was used to vertically scratch the 12-well plates. The cells were washed three times with PBS following glial trap coating of the wound in the treatment group. After a 12-h incubation with serum-free solution, the width of the scratch was measured and compared between groups using a microscope (Nikon TE2000). Five fields per group were randomly selected and photographed (10 × ).
Behavioral tests
The open-field test, rotarod test, and beam walking test, used for neurological evaluation, were carried out by two observers who were blinded to surgery conditions. The open-field test was performed 28 days after surgery. The apparatus included a central area (30 × 30 cm) within a square black box (60 × 60 × 25 cm). Mice were placed in the black box for 10 min and their behavior was recorded on video. The open-field test is based on the concept that mice prefer to be near a protective wall rather than expose themselves to the danger of an open field. For the rotarod test, mice were placed on an accelerating rotarod apparatus 1 day before surgery, and 14 and 28 days after surgery. The spin velocity of the apparatus was gradually accelerated to 40 rpm. The time at which the mouse fell from the rotarod apparatus was recorded. The beam walking test was performed at the same time points as the rotarod test. In this test, a horizontal wooden rod (1 cm in length and 1 cm diameter) was placed horizontally on a platform 60 cm high. The number of missteps and the total distance covered were recorded by two blinded observers.
Electrophysiological test
Mice were anesthetized with sodium pentobarbital followed by removal of the brain 28 days after surgery, which was placed in ice-cold artificial cerebrospinal fluid (ACSF) containing 124 mM NaCl, 1.2 mM NaH2PO4, 2.5 mM KCl, 24 mM NaHCO3, 5 mM HEPES, 12.5 mM glucose, 2 mM MgCl2, and 2 mM CaCl2. The target RFP+NeuN+-expressing neurons in the slices were identified under a BX51WI microscope (Olympus). For whole-cell measurements, brain slices were transferred to an interface chamber at 31°C–32°C and continuously superfused in oxygenated recording ACSF with recording electrodes (5–10 MΩ). Data were acquired by a PC running pClamp10 software, and whole-cell current was produced using a Multiclamp 700B amplifier and Digidata 1440A (Molecular Devices) that was interfaced by a converter. Signals were filtered when series resistance (Rs) values varied by 25% during the experiment, or when Rs values were >25 MΩ. All data were analyzed with Mini Analysis (Synaptosoft) and Clampfit (Molecular Devices) software.
Magnetic resonance imaging of injured brains
Magnetic resonance imaging (MRI) was performed using a 7T BioSpec scanner (Bruker). A 59-mm volume coil was used for radio frequency emission and reception. One day after surgery, the mice were anesthetized and immobilized in the magnet by a three-point fixation system. The scan parameters were: slice thickness = 1 mm, field of vision (FOV) = 2 × 2 cm, matrix = 256 × 256, and repetition time (TR) = 1,500 ms. The echo time values were within the range of 4–60 ms. Images were acquired using Paravision software (ver. 5.1; Bruker).
Statistical analysis
The data are presented as mean ± standard error of the mean. Cell counting was performed using Image J. All data were analyzed with independent-sample t-tests using SPSS software (ver. 13.0; SPSS, Inc.). *P < 0.05 and **P < 0.01 were considered to be statistically significant. Related data are shown in Supplementary Table S1.
Results
Neural matrix functions as a glial trap and enhances neurite regeneration
Following brain injury, regenerated neurites are sprouted and attempt to extend across the injured cortex during the process of repairing damaged regions. However, reactive astrocytes form a glial scar that functions as a physical and chemical barrier to prevent the regrowth of neurites [28]. In this work, we developed a neural matrix by chemical conjugation. The schematic diagram of the chemical reaction is shown in Fig. 1. In previous studies, SDF-1 was identified as a chemokine that mediates oriented cell migration, and astrocytes expressed SDF-1 receptor CXCR4. We immobilized SDF-1 in the neural matrix, hypothesizing that the matrix would attract reactive astrocytes [29,30]. To confirm this hypothesis, we performed in vitro migration tests using inclined slide glass (Supplementary Fig. S1A, B) and scratch assay (Supplementary Fig. S1D). In the control group, when the astrocytes cultured on the horizontal slide migrated to the inclined slide, the astrocytes migrated to the bottom level of the inclined slide because of the gravity. However, in the treatment group, the astrocytes migrated to the top level of the inclined slide because of the attraction of the glial trap (Supplementary Fig. S1A, B). The scratch assay (Supplementary Fig. S1D) further demonstrated the glial trap could attract the astrocytes. The glial cells tended to migrate to the neural matrix and, by staining with Ki67, we showed that this result was not due to glial cell proliferation. The biological activities of BDNF and NGF were tested by the survival assay of isolated dorsal root ganglia cells. From the results of Cell Counting Kit-8 (CCK-8) assay, we found that the bioactivity of BDNF and NGF was not affected by the conjugation (Supplementary Fig. S1B, C). Wild-type mice were subjected to brain injury, followed by treatment with the neural matrix for 14 days (Fig. 2A, B). Lesion area presented hyperproliferation of reactive astrocyte and neuronal apoptosis. In mice treated with neural matrix, the injury cavity was replaced by tissue, whereas a significant injury cavity persisted in the control group mice (Fig. 2C). We then stained the sections with antibodies to glial fibrillary acidic protein (GFAP), a marker for reactive astrocytes, CS-56, a marker for the deposition of chondroitin sulfate proteoglycans (CSPGs), and microtubule-associated protein 2 (MAP2) to analyze glial scar tissue and neuronal neurites in the injured cortex. Brain injury induced the accumulation of reactive astrocytes that were characterized by massive upregulation of GFAP in the injured cortex [31,32]. Figure 2D shows the significant accumulation of prominent reactive astrocytes in the injured cortex in the control group, whereas astrocyte accumulation was strikingly suppressed around the border between the lesion area and perilesional area in the treatment group at 14 days after injury. We also found the CSPGs were significantly decreased in the perilesional area in the glial trap group (Fig. 2E). The glial scar tissue that formed after brain injury was the major obstacle to neuronal regeneration. Notably, the neural matrix functioned as a glial trap that greatly decreased the area of reactive astrocytes in the injured cortex, and the reactive astrocytes were concentrated at the core of the lesion area (Fig. 2D). Thus, the area of reactive astrocyte activity was largely restricted in the treatment group relative to the control group.

The conjugation strategy immobilized the elements in the glial trap.
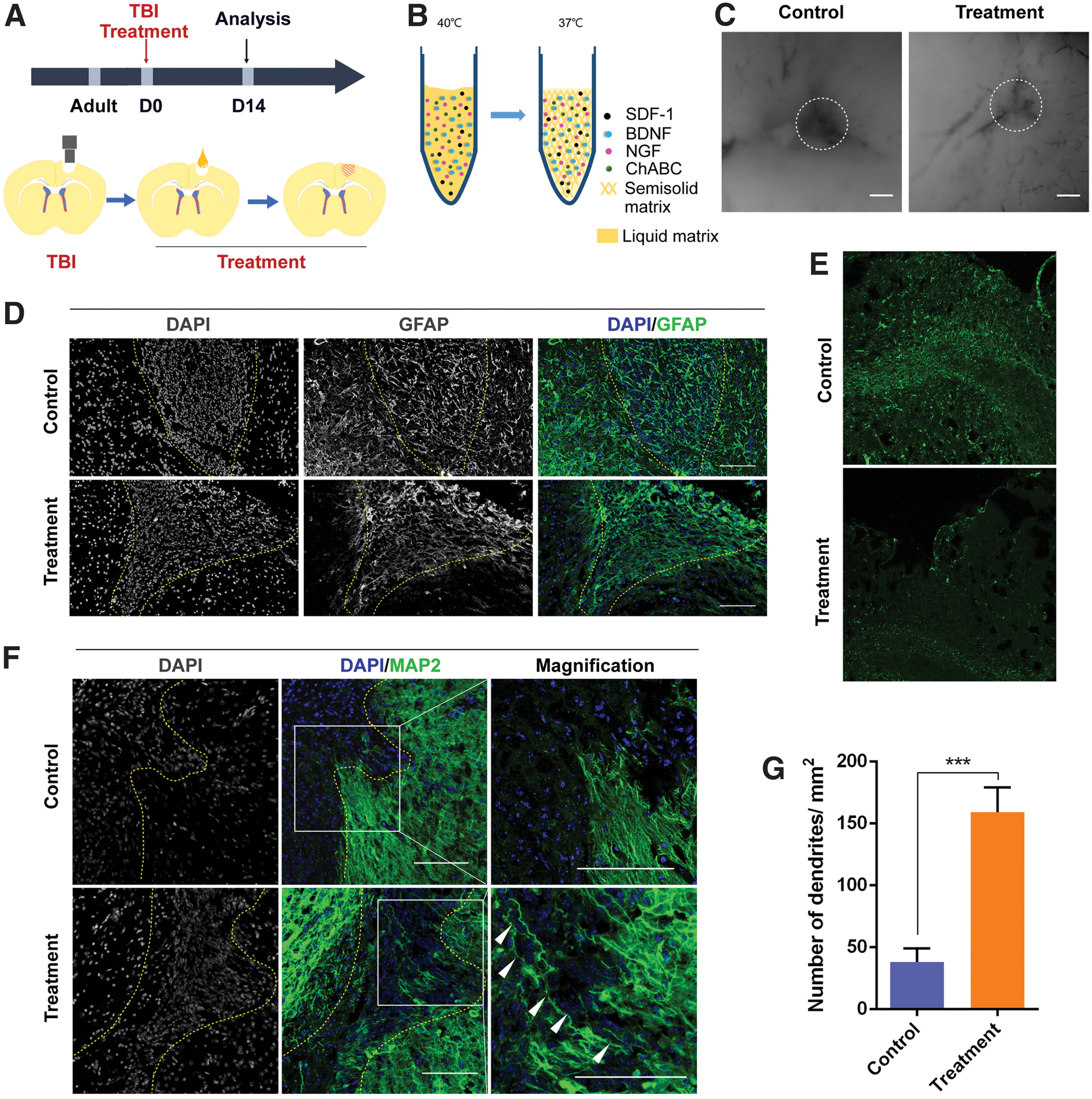
Reactive astrocytes are restricted to the core of the lesion area and neural circuits are regenerated following “glial trap” transplantation.
After staining with cell body and neuronal dendrite marker MAP2 to assess neurite regrowth after injury, we found that almost no neuronal neurites were present in the injured cortex in the control group (within the dotted lines in Fig. 2F and Supplementary Fig. S2C). In contrast, in the treatment group, we observed MAP2+ cells and obvious dendrites beyond the boundaries of the injured cortex and distributed within the injured cortex. In addition, statistical analysis revealed a significant difference in the number of neurites between the two groups (Fig. 2G). These results suggest that the decreased glial scar contributed to the MAP2+ neurons and neurites penetrating and passing through the boundary of the injured cortex (Supplementary Fig. S2).
Lineage tracing of eNP/SCs in adult offspring of Nestin-CreERT2 mice crossed with R26R-RFP mice
An inducible tool nestin-CreERT2 mice was crossed with R26R-RFP mice to trace eNP/SCs. In this way, Cre-mediated recombination only occurred in nestin-expressing cells upon administration of TAM by oral gavage in adult offspring [33]. Two days after TAM treatment, we analyzed the characteristics of eNP/SCs in the SVZ by immunostaining for the NP/SC marker nestin and red fluorescent protein (RFP)-positive cells. Our results showed that RFP only labeled NP/SCs (Fig. 3A, B), which confirmed the efficiency and specificity of transgene expression in the nestin-expressing cells of nestin-CreERT2/+; R26R-RFP/+ mice. Consistent with previous studies showing that NP/SCs resided at the SVZ [34], our results revealed that RFP-positive cells merged with nestin-positive cells localized only at the SVZ. Quantitative statistical analysis showed that 95.64% of the nestin+ cells were co-labeled as RFP-positive cells (Fig. 3B, C). We then further stained the SVZ and normal cortex with NeuN, a marker of mature neurons (Fig. 3D), and found that RFP+ cells were not detected in the normal cortex (Fig. 3E). These results indicated that our methods specifically marked NP/SCs. Therefore, in the following experiment, the RFP-positive cells were derived from svzNP/SCs and unable to be derived from other cells.
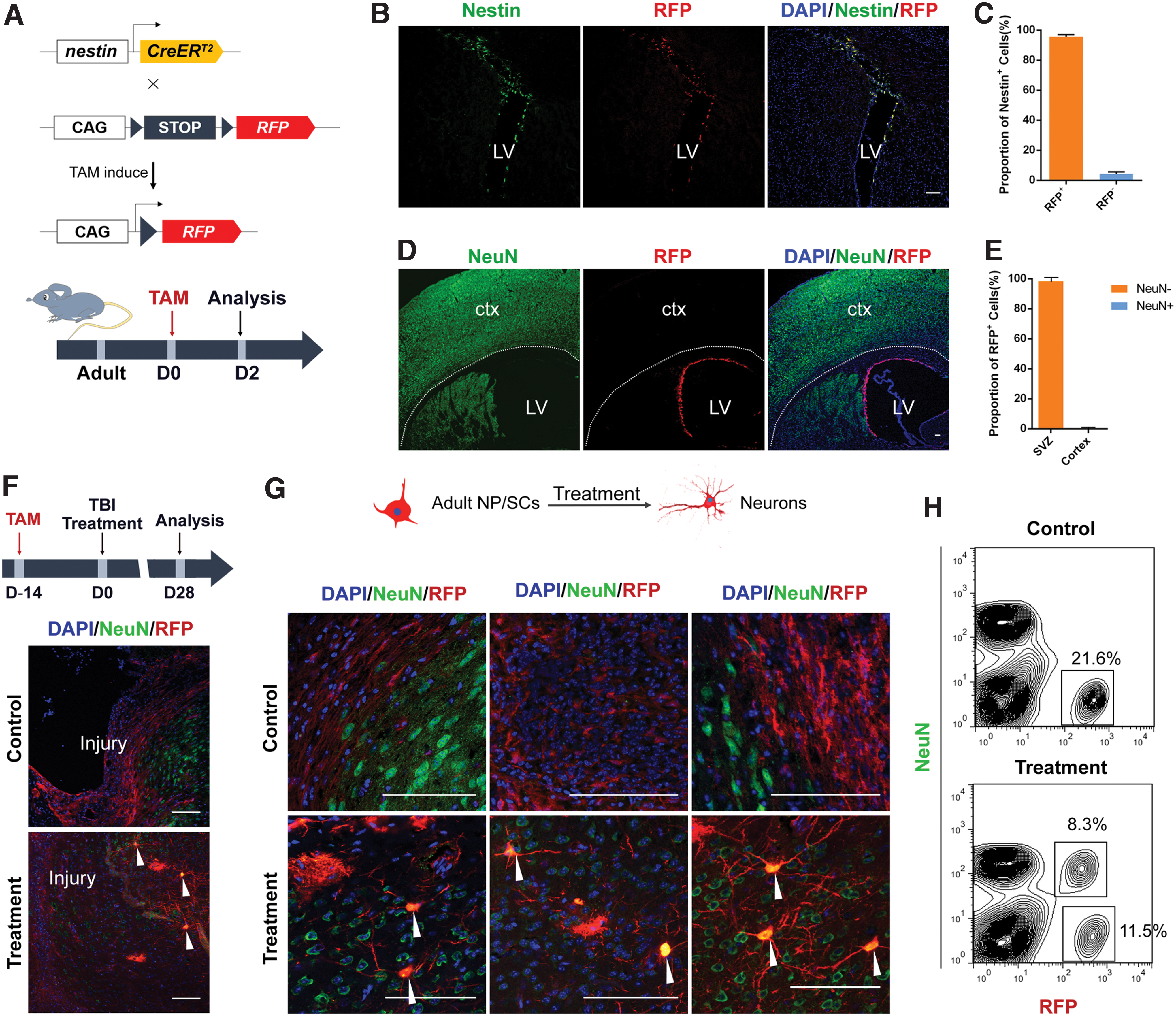
Adult NP/SCs are induced to differentiate into mature neurons in injured cortex under the influence of the glial trap.
Brain injury activates SVZ-derived NP/SCs, facilitating their differentiation into mature neurons
Following brain injury, SVZ-derived NP/SCs home in on the injured cortex to become astrocytes in the adverse milieu that participates in glial scar formation [35]. In our work, the decreased glial scar seen in the injured cortex, and the trophic signals from BDNF and NGF in the neural matrix, may change the milieu from a nonpermissive to a permissive one, thus altering the fate of eNP/SCs. MRIs of mice after brain injury are shown in Supplementary Fig. S3B. Twenty-eight days after brain injury, sections were placed under a fluorescence microscope without immunostaining, and we observed RFP+ eNP/SCs in the injured cortex (Supplementary Fig. S3C), suggesting migration of eNP/SCs to the injured cortex [36 –39]. We next investigated whether the SVZ-derived eNP/SCs could differentiate into neurons. At 28 days after brain injury, we stained brain tissue sections of the injured cortex of the control and treatment groups for NeuN and RFP (Fig. 3F). We were surprised to find that NeuN-positive cells co-localized with RFP-positive cells in the injured cortex, mainly around the border of the lesion area and perilesional area, only in the treatment group and not in the control group (Fig. 3F, G). These results indicated that after eNP/SCs left the SVZ, they migrated toward the injured cortex following brain injury. The glial trap ameliorated the niche of the injured cortex, especially in the perilesional area, and induced the differentiation of the eNP/SCs into mature neurons during the repair process. The percentage of regenerated neurons was 32.45% ± 8.57% in the glial trap group assessed by fluorescent-activated cell sorting (FACS) (Fig. 3H). The acquisition of neurons in the injured cortex has been investigated over the past few years, and neuronal reprogramming represents significant progress in this field [40,41].
We performed further immunostaining to investigate the characteristics of early SVZ-derived cells above the corpus callosum. On day 9 after brain injury, 98.26% of the nestin+ cells co-localized with RFP in the injured cortex (Fig. 4A, B), and RFP+ SVZ-derived NP/SCs began to differentiate. We stained with anti-RFP to track eNP/SCs 14 days after surgery (Fig. 4C). We found that the eNP/SCs migrated to the injured cortex from the ipsilateral side, whereas this phenomenon was not observed at the contralateral side (Fig. 4D). We also stained with immature neuron marker βIII-tubulin (TuJ1) and GFAP to determine the fate of RFP+ SVZ-derived NP/SCs in the injured cortex. Our results showed that almost all of the RFP+ cells were co-labeled with GFAP+ cells, and no TuJ1+ cells were found in the control group. However, substantial numbers of TuJ1+ cells merged with RFP+ cells in the treatment group (Fig. 4E, F). This indicated that the SVZ-derived NP/SCs in the control group differentiated into reactive astrocytes. After application of the glial trap for brain injury, substantial numbers of SVZ-derived NP/SCs differentiated into immature neurons. The results from FACS further showed that the percentage of differentiated reactive astrocytes was lower in the glial trap group when compared with that in the control group (Supplementary Fig. S4). Because the fate of NP/SCs depends on their microenvironment [42], the different fates of SVZ-derived NP/SCs between the control group and the treatment group may be attributable to different microenvironments within the injured cortex.
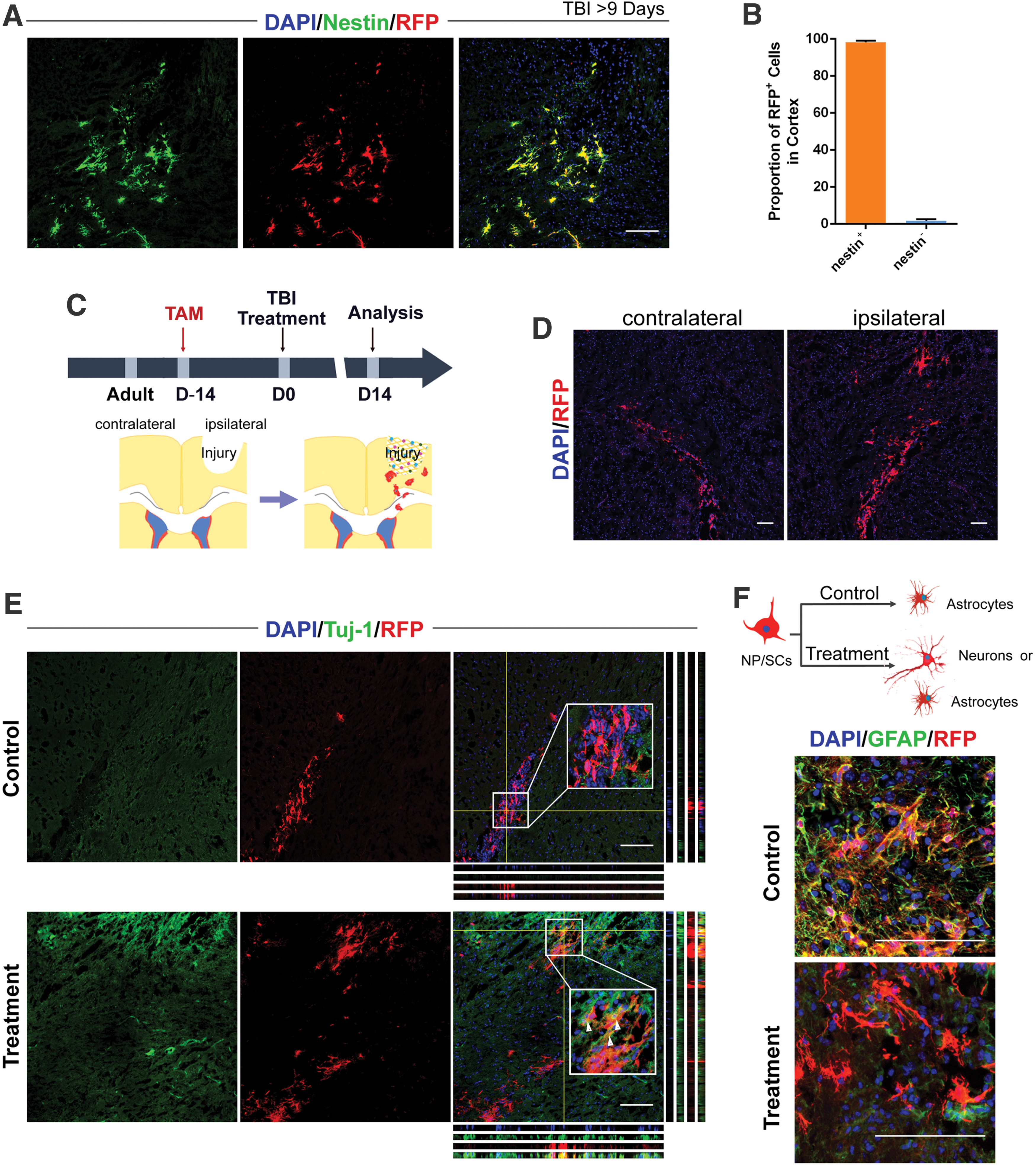
Adult NP/SCs migrate from the SVZ to the injured cortex and differentiate into immature neurons after glial trap treatment.
Regenerated neurons are functional pyramidal neurons
To further characterize the properties of regenerated neurons, we performed immunostaining with different neuronal markers to identify cell types. Figure 5A confirms that the eNP/SC-derived neurons were positive for the pyramidal neuron marker CamKII, and negative for the interneuron markers PV and Som (Fig. 5B, C). Vesicular glutamate transporter 1 (VGluT1) is an important presynaptic marker of glutamatergic neurotransmission that is expressed in cortical and subcortical glutamatergic neurons [43]. The eNP/SC-derived pyramidal neurons were immunopositive for VGluT1, accompanied by mushroom-like mature spines (Fig. 5D). We then performed electrophysiological tests to examine the properties of eNP/SC-derived pyramidal neurons using the patch clamp technique (Fig. 5E). These regenerated neurons fired repetitive action potentials, demonstrating membrane excitability and thus indicating that they were excitatory neurons (Fig. 5E). In a previous study, reprogramming by forced expression of TF Ascl1 successfully reversed SVZ-derived reactive astrocytes to neurons in vivo [17]. In comparison to this reprogramming approach, the functional neurons in our study were obtained before the eNP/SCs differentiated into reactive astrocytes. We then performed an in vitro study to examine the safety of our constructed neural matrix. Human fetal NP/SCs were cultured on neural matrix in multiwell plates for 3 days. There were no significant differences in the number of human fetal NP/SCs between the control and treatment groups.
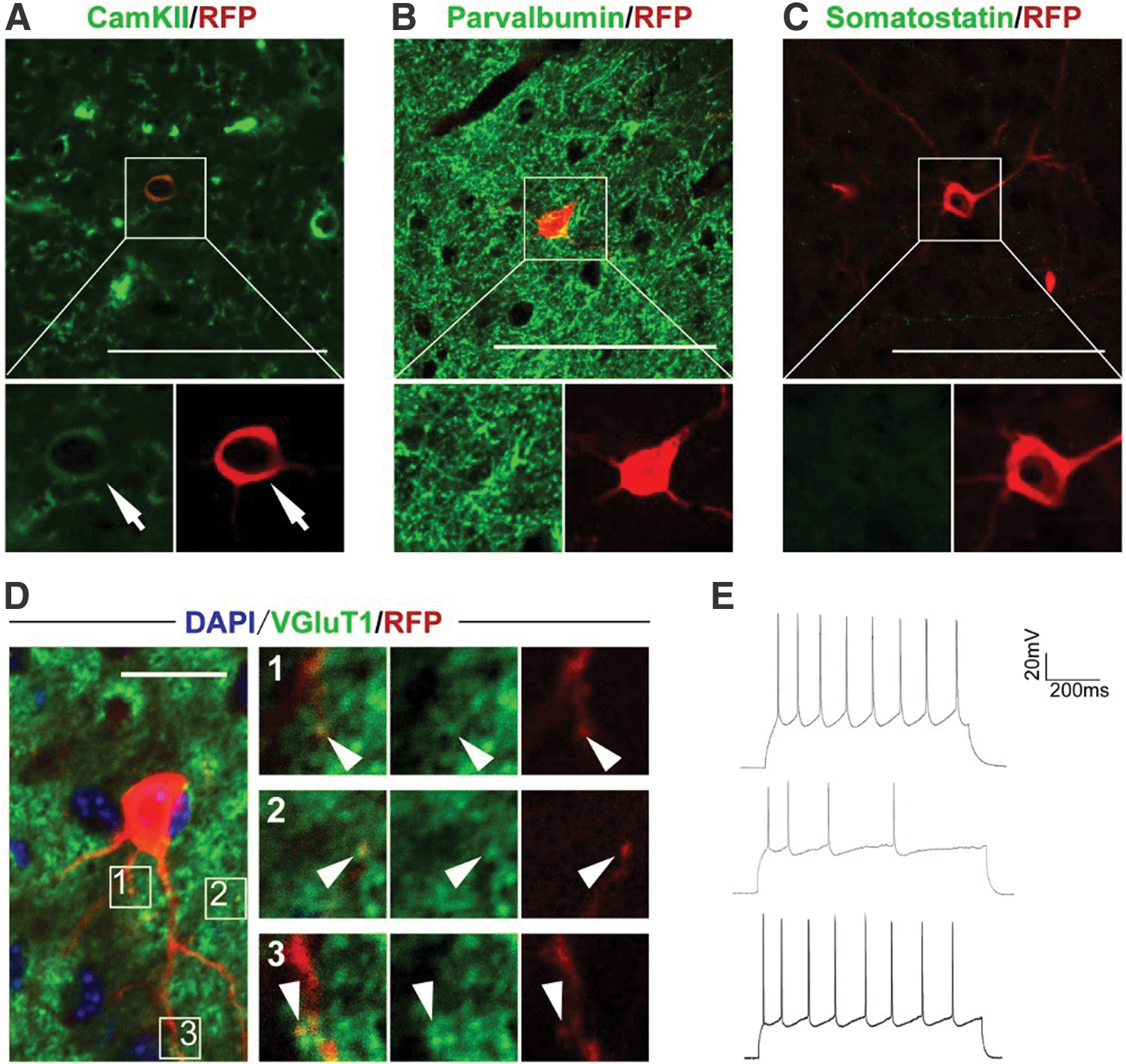
eNP/SCs give rise to pyramidal neurons in the cerebral cortex.
Glial trap-treated mice achieve functional recovery
Whether glial trap treatment could produce functional improvement in injured mice was a major focus of our study. Functional assessments were performed, including the open-field test at 28 days postsurgery, and the rotarod test and beam walking test, performed at −1, 14, and 28 days postsurgery (Fig. 6A). The cumulative distance traveled, distance traveled in the center region, and cumulative vertical activities in the open-field test were significantly higher in the treatment group than in the control group (P < 0.05), indicating motor recovery and exploration recovery in glial trap-treated mice (Fig. 6B–E). The rotarod test and beam walking test were used to evaluate motor function and balance, respectively. The rotarod test results in sham mice did not differ among the three time points. Similar to the results of the open-field test, the latency to falls in the treatment group was significantly higher than in the control group at 14 and 28 days postsurgery (P < 0.05) (Fig. 6F). In the beam walking test, the number of errors/distance was significantly higher in the control group than in the other groups at 14 and 28 days postsurgery (P < 0.001) (Fig. 6G). The results showed that there was no significant significance between the sham group and treatment group for the cumulative traveled distance, distance traveled in the center, cumulative vertical activities, and latency to falls at 28 days following surgery. The functional outcomes were attributed to reduced glial scar formation and strong neurogenesis in mice treated with the glial trap. Because rescuing behavioral deficits remains a therapeutic challenge for in vivo reprogramming [44], our strategy showed significant advantages for functional recovery.

Attenuated injury-induced behavioral deficits in mice treated with the glial trap.
Discussion
SVZ-derived NP/SCs home in on the injured cortex following brain injury; therefore, we crossed Nestin-CreERT2 mice with R26R-RFP mice to lineage trace the fate of adult eNP/SCs after the neurogenic microenvironment was altered by the neural matrix. The SVZ is near the cortex, enabling easy migration of NP/SCs in the SVZ to the injured cortex for repair following brain injury [39]. Targeting these eNP/SCs to achieve repair and regeneration of brain injuries avoids the immune rejection associated with the transplantation of exogenous NP/SCs [45 –48]. Results from the control group in this study accorded with previous studies, although eNP/SCs migrated from the SVZ into the injured cortex and no neurons were seen within the injured cortex 28 days after injury [36].
Brain injury induced astrocytes to become reactive astrocytes that secreted growth-inhibiting molecules, and together they formed glial scars that resulted in failure of neurite regrowth and reestablished the neuronal network at the lesion area and the perilesional area [28,49]. “Melting” glial scars have been studied for the past few decades, with little success. CSPGs produced by reactive astrocytes are among the most important elements in glial scar formation. CSPGs inhibit neurite regeneration of injured neurons and serve as nonpermissive cues for the eNP/SCs [50,51]. In this study, ChABC is used to ablate chondroitin sulfate glycosaminoglycans, which contribute to the inhibitory function of CSPGs, thus destroying the chemical barrier of glial scars [52,53]. We also found that the neural matrix largely decreased the area of reactive astrocytes, consequently reducing glial scar deposition in the injured cortex, and the neural matrix replaced the injury cavity in the injured cortex with neural tissue (Fig. 2C, D). The ablated glial scar in the injured cortex contributed to the extension of neurites across the boundary of the injured cortex (Supplementary Fig. S2C).
The cell niche at the injured cortex limits the differentiation of migrated eNP/SCs into neurons, as postnatal eNP/SCs generate glial cells rather than producing new neurons [54 –56]. The eNP/SC-derived reactive astrocytes and reactive astrocytes, which are locally generated in response to brain injury, together participate in glial scar formation [55,56]. Previous studies have shown that NP/SCs are self-renewing, multipotent cells with the ability to differentiate into astrocytes or neurons depending on the environmental conditions [57]. Whether the neural matrix formed a suitable microenvironment in the injured cortex for activated eNP/SCs to give rise to neurons was the key to brain injury repair in this study. Our initial experimental showed that MAP2+ neurons scattered at the lesion area and the perilesional area in the matrix-treated group in wild-type mice (Fig. 2F and Supplementary Fig. S2). Reprogramming by turning one neuronal subtype into another is somewhat difficult and has been shown to produce mostly immature cells with poor survival time [58]. We assessed the distribution of NeuN+/RFP+ mature neurons in the injured area at 28 days after treatment with neural matrix (Fig. 3G). The results indicated that eNP/SCs were activated after brain injury and migrated into the injured cortex, differentiating into mature neurons. These mature neurons were primarily located in the perilesional area of the injured cortex, likely because the glial trap concentrated reactive astrocytes in the core of the injured cortex, leaving the surrounding area as a favorable niche. The new neurons were subsequently shown to be functional pyramidal neurons positive for the glutamatergic synaptic marker VGluT1. SDF-1 is also an attractant for SVZ-derived cells following injury, but we find the number of RFP+ cells was not significantly different between the control group and glial trap group in the injured cortex. It may be because the SDF-1 was chemically conjugated to the glial trap and thus its effects were confined to a local area.
Notably, the glial trap concentrated the reactive astrocytes within the core of the lesion area in the treatment group, whereas the reactive astrocytes were distributed within both the lesion area and the perilesional area in the control group. BDNF and NGF, two elements of the neural matrix, are important neurotrophic factors enhancing neural regeneration. The elements in the neural matrix may have provided a favorable milieu for the migrated eNP/SCs to give rise to functional neurons. In addition to their nonpermissive role, CSPGs also play an important role in the formation of the perineuronal net that separates neuronal cells from each other by hyaluronan [59]. ChABC degrades chondroitin sulfate, as well as hyaluronan; therefore, ChABC treatment helped to destroy the perineuronal net and allowed for free interactions between new functional neurons and other neurons in our study. Taken together, these effects may result in functional improvements in mice. Twenty-eight days after brain injury, the functional behavior of mice in the treatment group recovered to an outstanding degree, compared to the level of normal, uninjured mice. Functional behavior in the control group exhibited only limited recovery. The improved functional outcomes were attributed to reduced glial scar formation and strong neurogenesis in mice treated with the glial trap.
To obtain neurons for brain injury repair, eNP/SCs could be expanded and manipulated with gene constructs to control their fate in vitro, and then transplanted into the injured cortex [60]. In another study, Guo et al. showed that reactive glial cells can be directly reprogrammed into functional neurons by retroviral expression of the TF NeuroD1 [44]. In recent years, several reprogramming approaches for turning non-neuronal cells, such as reactive astrocytes, NG2 glia, or fibroblasts, into neurons have been developed [61 –63]. Although significant progress has been made in reprogramming, strategies involving viruses, microRNAs, or small molecules raise serious safety concerns for clinical utilization. A previous study showed that eNP/SCs normally migrate in the confined rostral migratory stream to reach the OB [64,65]. Large numbers of OB interneurons are replaced by new interneurons differentiated from NP/SCs migrating from the SVZ [66,67]. We hypothesized that the OB may contain special proteins or molecules involved in homeostasis for stem cell differentiation. Therefore, we constructed a neural matrix from OB to create a favorable niche at the injured cortex for neurogenesis after brain injury. In addition, SDF-1, BDNF, NGF, and ChABC were anchored in the neural matrix by chemical conjugation to surmount their rapid diffusion from the neural matrix in body fluids, thus avoiding the potential side effects and low efficiency caused by their diffusion. The decreased glial scar formation and trophic signals of BDNF and NGF may convert the adverse niche into a favorable niche for neurogenesis. Our strategy did not involve gene regulation or the use of small molecules to obtain functional neurons, thus constituting a further advancement toward potential clinical applications.
Footnotes
Acknowledgments
We thank Professors Zhengang Yang and Yunli Xie of the Institutes of Brain Science of Fudan University for providing technical support for Nestin-CreERT2 and R26R-RFP mice construction.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from Shanghai Sailing Program (19YF1404600), the National Nature Science Foundation and Ministry of Science and Technology of China (2018YFA0107900, 31771491), and Shanghai Municipal Government (2019CXJQ01).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.