
Abstract
Bisphosphonate-related osteonecrosis of the jaws (BRONJ) is a severe adverse reaction, which results in progressive bone destruction in the maxillofacial region of patients. To date, the pathological mechanisms remain largely unclear. Recently, we found that BRONJ patient had significantly deep periodontal pockets and severe periodontal bone defects before the exposed necrotic bone. Human periodontal ligament stem cells (hPDLSCs) play key roles in physiological maintenance and regeneration of periodontal tissues. However, the activities of hPDLSCs derived from BRONJ lesions and the role of hPDLSCs in BRONJ periodontal defect repair remain poorly understood. The aim of the present study was to elucidate the role of hPDLSCs in BRONJ. In this study, we found that the capacities of cell proliferation, adhesion, and migration of hPDLSCs derived from BRONJ lesions (BRONJ-hPDLSCs) were significantly decreased compared with control-hPDLSCs. BRONJ-hPDLSCs underwent early apoptosis compared with control-hPDLSCs. Importantly, we first demonstrated that BRONJ-hPDLSCs exhibited impaired osteogenic differentiation abilities in ectopic osteogenesis of nude mice. The above results suggested that the impaired BRONJ-hPDLSCs may be an important factor in deficient periodontal repair of BRONJ lesions and provide new insight into the underlying mechanism of BRONJ.
Introduction
Bisphosphonate-related osteonecrosis of the jaws (BRONJ), a devastating side effect induced by long-term bisphosphonate (BP) use, is a serious threat to patients [1,2]. Since 2003, Marx first reported osteonecrosis of the jaw in patients treated with high doses of intravenous BPs, an increasing number of BRONJ cases have been documented [3 –5]. Various hypotheses surrounding the etiology of BRONJ have been proposed, among which excessive suppression of the jaw bone turnover, inflammation, angiogenesis inhibition, soft tissue toxicity, and the immune system are the most popular theories [6 –9]; however, the exact pathophysiological mechanisms and the reasons why BRONJ mainly occurs in the jaw remain largely unknown.
Maxilla and mandible are the only bones in the body that hold the teeth in place and are covered by a thin layer of mucosa in a very close proximity to the external environment, where it is susceptible to periodontal disease and procedures such as dental extractions [10,11]. Previous case reports and animal studies have identified periodontal disease as one of the most important risk factors for the development of BRONJ [12 –15]. Clinically, many reports have noted that BRONJ occurs after extraction of teeth due to the severity of dental disease or around teeth with active periodontal or periapical disease [16 –20]. In 2014, Thumbigere-Math et al. conducted a case–control study and showed that BRONJ patients have fewer teeth, greater clinical attachment level, and less alveolar bone support than controls [14].
Numerous researchers have also studied the role of periodontal disease in BRONJ development using animal models [7,21]. Aguirre et al. recently found that oncologic doses of zoledronate exacerbated periodontal tissue damage and induced BRONJ-like lesions in rats [13]. In the clinical examination of patients who received intravenous BP therapy for cancer management, we also found that the BRONJ patient had significantly deep periodontal pockets and severe periodontal bone defects before the exposed necrotic bone. Nevertheless, there were no studies to investigate the underlying processes or mechanisms regarding poor periodontal repair and regeneration in BRONJ lesions.
In a seminal article, He et al. isolated bone marrow stromal cells (BMSCs) from the osteonecrotic BRONJ regions and demonstrated that the decreased activities of BRONJ BMSCs, even the BMSCs derived from the BRONJ lesion boundary, might be an important factor leading to insufficient bone repair of BRONJ lesions [22]. They did not explore the mechanism of osteonecrosis of the jaw and why it only occurred in the jaw. Nevertheless, the findings that the changes of biological characteristics of BMSCs in patients treated with BPs provided us with the insights to further explore the causes of osteonecrosis. Periodontal ligament stem cells (PDLSCs), a newly recognized subpopulation of mesenchymal stem cells, represent key players during periodontal repair and regeneration [23,24]. There was evidence that PDLSCs' dysfunction or apoptosis may lead to poor periodontal repair and reduce periodontal bone regeneration [25,26]. However, there have been no reports evaluating the activity of PDLSCs in BRONJ lesions and their osteogenic capability.
To explore the roles of human periodontal ligament stem cells (hPDLSCs) in BRONJ periodontal defect repair, we isolated hPDLSCs derived from BRONJ lesions and assessed their activities by a variety of methods, including CCK-8 assays, alkaline phosphatase (ALP) assays, Alizarin Red staining, immunofluorescence, flow cytometry, and real-time polymerase chain reaction (RT-PCR) in vitro. To determine their osteogenic capability in vivo, we assessed the ectopic osteogenesis of beta-tricalcium phosphate (β-TCP) combined with hPDLSCs in nude mice.
To our knowledge, this study is the first report of evaluating the activity of PDLSCs in BRONJ lesions and their osteogenic capability, as well as examining periodontal tissue changes in vitro and in vivo. The study provided new insight into the underlying mechanism of BRONJ.
Materials and Methods
Patients
Five cancer patients (two males and three females), ages 54–81 years, underwent surgery at the Department of Oral Surgery, Ninth Peoples Hospital. Debridement of the affected bone and teeth was extended horizontally and inferiorly to reach healthy-appearing tissues [1,27]. Patients were considered eligible for this study if they had a histologically confirmed advanced solid cancer and radiographic confirmation of bone metastases, receiving intravenous BPs and presented with exposed necrotic bone in the maxillofacial region at least eight [28,29]. Patients were considered ineligible when they had received any radiotherapy, chemotherapy, immunotherapy, or hormonotherapy before the study. The control group included five patients, who were older than 50, without bone metabolism diseases, who underwent third molar extraction operation. All patients provided written informed consent to participate in this study. This work was approved by the Ethics Committee of the Ninth Peoples Hospital affiliated to Shanghai Jiao Tong University, School of Medicine. Detailed patient information is listed in Appendix Table A1.
Animals
All animal procedures were approved by the Animal Care and Experiment Committee of the Ninth People's Hospital, which is affiliated with Shanghai Jiao Tong University, School of Medicine, China, and the study was carried out according to the guidelines for the Ethical Conduct in the Care and Use of Nonhuman Animals in Research by the American Psychological Association.
Micro–computed tomographic scanning
The samples were fixed with 4% paraformaldehyde and subjected to micro–computed tomography (μCT) scanning (μCT 80; Scanco Medical, Bassersdorf, Switzerland) at 10-μm resolution. Two-dimensional slices were combined using μCT v6.1 software (Scanco Medical) to form a three-dimensional reconstruction. The volume of periodontal bone volume to total volume ratio was analyzed using Scanco Medical software. We quantified the width of PDL using ImageJ (National Institutes of Health).
Histological assessment of BRONJ and healthy periodontal tissues
After μCT scanning, these samples were decalcified and processed for paraffin embedding. Histological sections (4 μm thick) per sample were cut and stained with Hematoxylin and Eosin (H&E) and Masson staining.
Isolation of hPDLSCs from BRONJ and healthy periodontal tissues
hPDLSCs were isolated and cultured according to previously reported protocols with a slight modification [25]. In brief, periodontal ligament (PDL) tissues were gently scraped from the surface of the middle part of the root, minced into 1-mm3 cubes, and placed into six-well culture dishes. Then glass cover slips were placed over the tissues to prevent floating, and PDL tissues were incubated in culture medium (DMEM with 10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 μg/mL streptomycin) in a humidified atmosphere of 95% air and 5% CO2 at 37°C. The tissues were subsequently maintained by replacing the medium every 3–4 days until cell density reached confluence.
Flow cytometry
Cell-surface antigen expression was assessed using flow cytometry analysis. Approximately 1.0 × 106 cells were washed in phosphate-buffered saline (PBS) and incubated with mouse anti-human CD146, STRO-1, CD45, CD90, and CD34 for 30 min at 4°C. Control conjugated immunoglobulin G was used as the isotype control. All antibodies were purchased from BD Biosciences. Cells were washed twice with cold PBS containing 2% FBS. Labeled cells were analyzed using a flow cytometer (Beckman Coulter, Fullerton, CA).
Cell proliferation assay
Third-passage hPDLSCs were seeded at a density of 3 × 103 cells/mL into a 96-well plate. The cell number was assessed on days 1–7 using Cell Counting Kit-8 (Dojindo Laboratories) assay. The optical density was measured at a wavelength of 450 nm using the Spark™ 10M Multimode Microplate Reader (TECAN, Switzerland).
Cell adhesion
The experiment was performed according to a previously reported protocol [30]. Briefly, third-passage hPDLSCs were divided into 5.0 × 104 cells/mL and incubated in Dulbecco's modified Eagle's medium (DMEM) for 30 min. Cells were subsequently seeded onto type I collagen coated six-well plates containing the same media and incubated for 30 min. Then, the wells were rinsed vigorously three times with PBS, and the remaining cells were stained using 0.1% Crystal Violet dye. Data are expressed as adherent cells per field.
Cell scratch assay
Third-passage hPDLSCs were plated at 200,000 cells per well in six-well plates. Once confluent, a scratch wound was performed using a sterile 10 μL pipette tip. The size of the gap was measured microscopically immediately (0 h), 6, and 24 h later.
Determination of apoptotic cells
Third-passage hPDLSCs were plated at 30,000 cells per well in six-well plates. For cell apoptosis, after cells were fixed in 75% ice-cold ethanol, apoptosis was analyzed using FITC Annexin V Apoptosis Detection Kit I (BD Pharmingen™; Biosciences, San Diego, CA).
ALP activity and Alizarin Red staining
Third-passage hPDLSCs were plated at 30,000 cells per well in 24-well plates. For ALP staining, PDLSCs were induced in the osteogenic medium for 14 days, then cells were stained with the BICP/NBT Alkaline Phosphatase Color Development Kit (Beyotime). The quantitative evaluation for ALP activity was performed using the Alkaline Phosphatase Assay Kit (Beyotime). For Alizarin Red staining, PDLSCs were induced in the osteogenic medium for 21 days, and then, cells were stained with 1% Alizarin Red (Sigma-Aldrich). To measure the concentration of calcium, the Alizarin Red dye was extracted with 400 μL of 10% cetylpyridinium chloride for 15 min at room temperature, and then quantified at a wavelength of 562 nm using the Spark 10M Multimode Microplate Reader (TECAN).
Real-time polymerase chain reaction
Third-passage hPDLSCs were plated at 1.0 × 105 cells per well in six-well plates. After 24 h, total RNA was isolated from cells using with TRIzol (Invitrogen; Carlsbad, CA) and reverse transcribed by Superscript II (Invitrogen). Real-time PCR was performed with Light Cycler® 480 II (Roche, Switzerland). β-Actin was used to normalize gene expression. The primers used in this study are shown in Appendix Table A2.
Immunofluorescence
Third-passage hPDLSCs were plated at 3 × 104 cells per well in 12-well plates. After 24 h, cells were fixed in 4% paraformaldehyde for 30 min and permeabilized with 0.3% Triton X-100 for 10 min. After washing with PBS, the cells were incubated for 1 h in a blocking solution (5% BSA in PBS) at room temperature and treated with both antibodies COLIA1 and OCN (1:200; Abcam) at 4°C overnight. Then, cells were incubated with secondary antibodies (1:500; Invitrogen) for 30 min and subsequently incubated with DAPI (1:500; Abcam) for 5 min at room temperature. Fluorescence images were obtained using a fluorescence microscope (Olympus, Tokyo, Japan).
In vivo bone formation assays
Bone formation was analyzed with ectopic osteogenesis in nude mice. Approximately 5 × 107 cells were added to the β-TCP cuboids (Shanghai Bio-Lu Biomaterials Co. Ltd., Shanghai, China) forming the constructs and transplanted into aseptically created subcutaneous pockets in 6-week-old nude male mice (SCID Beige Mouse; Charles River Laboratories International) under anesthesia through 2% sodium pentobarbital. Fifteen constructs were divided into three groups: Group A, β-TCP alone, n = 5; Group B, β-TCP/control-hPDLSCs, n = 5; and Group C, β-TCP/BRONJ-hPDLSCs, n = 5. Transplants were harvested after 12 weeks and assessed by histology.
Statistical analyses
All statistical analyses were performed using GraphPad Prism 7 (GraphPad, Software). The outcome measurements are expressed as the mean ± standard deviation. Differences between two groups were analyzed by t-test. P ≤ 0.05 was considered as the statistically significant difference for all comparisons. All experiments were conducted in triplicate.
Results
Radiographic analysis of BRONJ and healthy periodontal tissues
Representative 3D reconstructed images of extracted teeth from BRONJ patients and control patients are depicted in Fig. 1A. The radiographic image in control patients showed that the surface of normal inherent alveolar bone was smooth and scattered with needle-like pores, which provides attachment for the PDL fiber bundles firmly supporting the root of the tooth (black arrow). In contrast, the alveolar bone in BRONJ patients presented a remarkably different radiographic appearance, which was shown as coarse reticulation and extensive honeycomb destruction in inherent alveolar bone (black arrow). Two-dimensional slices (Fig. 1B) showed expanded PDL spaces, absent lamina dura and irregular absorption in alveolar bone with an eroded edge in BRONJ patients (yellow arrow). In addition, quantitative data (Fig. 1C) based on μCT images showed that alveolar bone loss and expanded PDL in patients administered with zoledronate (ZOL) were statistically significant compared with the control groups.
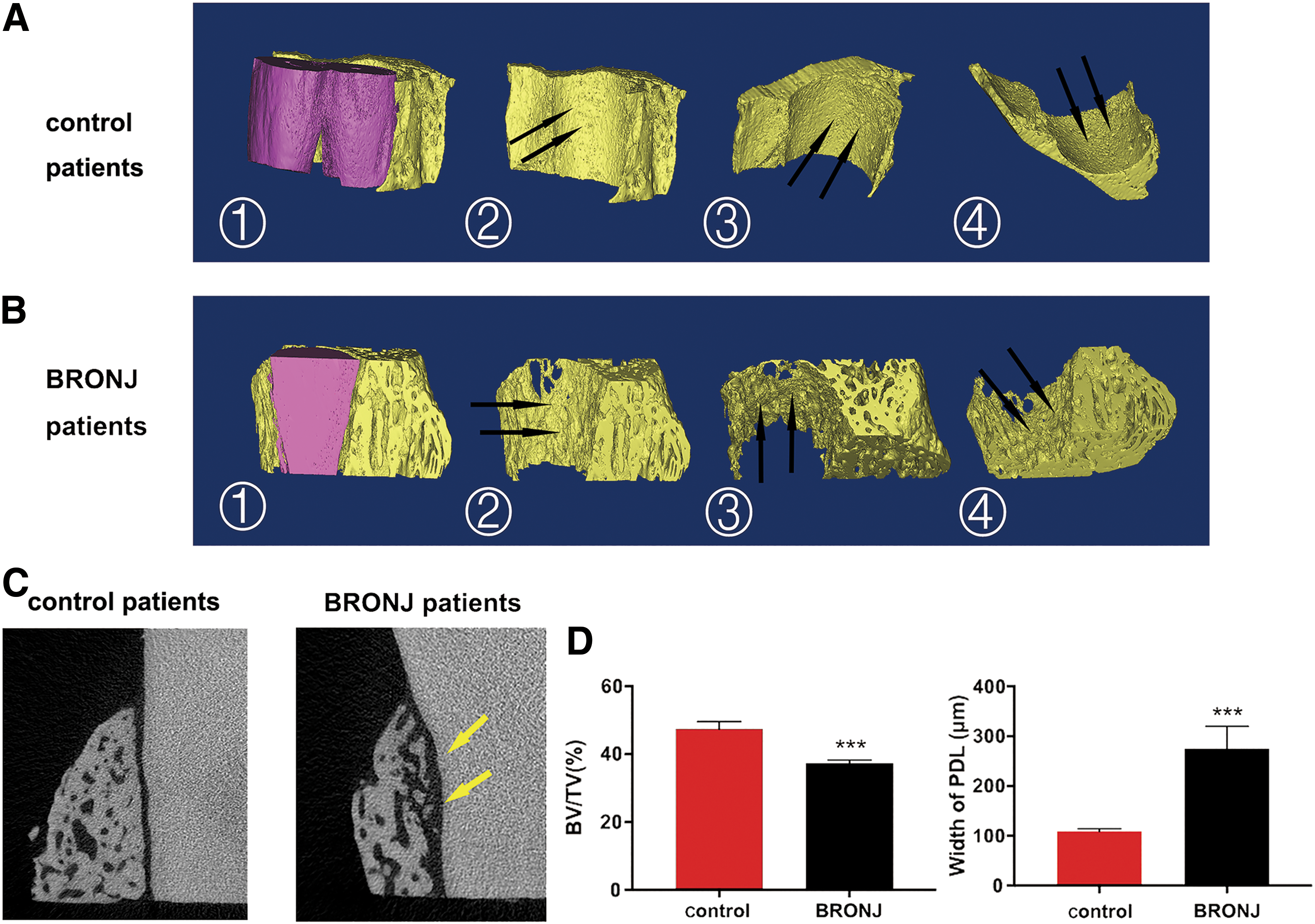
μCT analysis the periodontal microstructure in BRONJ patients and control patients.
Histological analysis of BRONJ and healthy periodontal tissues
To evaluate the histological appearance of PDL and alveolar bone around the root in BRONJ patients and control patients, the periodontal tissues were examined for decalcification, H&E, Masson staining, and immunohistochemical analysis. Figure 2A1 notes the normal histological features of the tooth root, PDL, and the alveolar bone in control patients. However, in sections from the target sites of BRONJ patients, necrotic bone characterized by empty osteocyte lacunae (yellow arrows), severely malformed PDL fibers, and significant destruction in alveolar bone (Fig. 2A) of BRONJ patients were observed. Masson staining assay (Fig. 2B) of nondecalcified tissues displayed a disorganized PDL area showing cracked, edematous fibers and a major reduction of collagen fibers (yellow arrow) in BRONJ patients compared with control patients (Fig. 2B1).
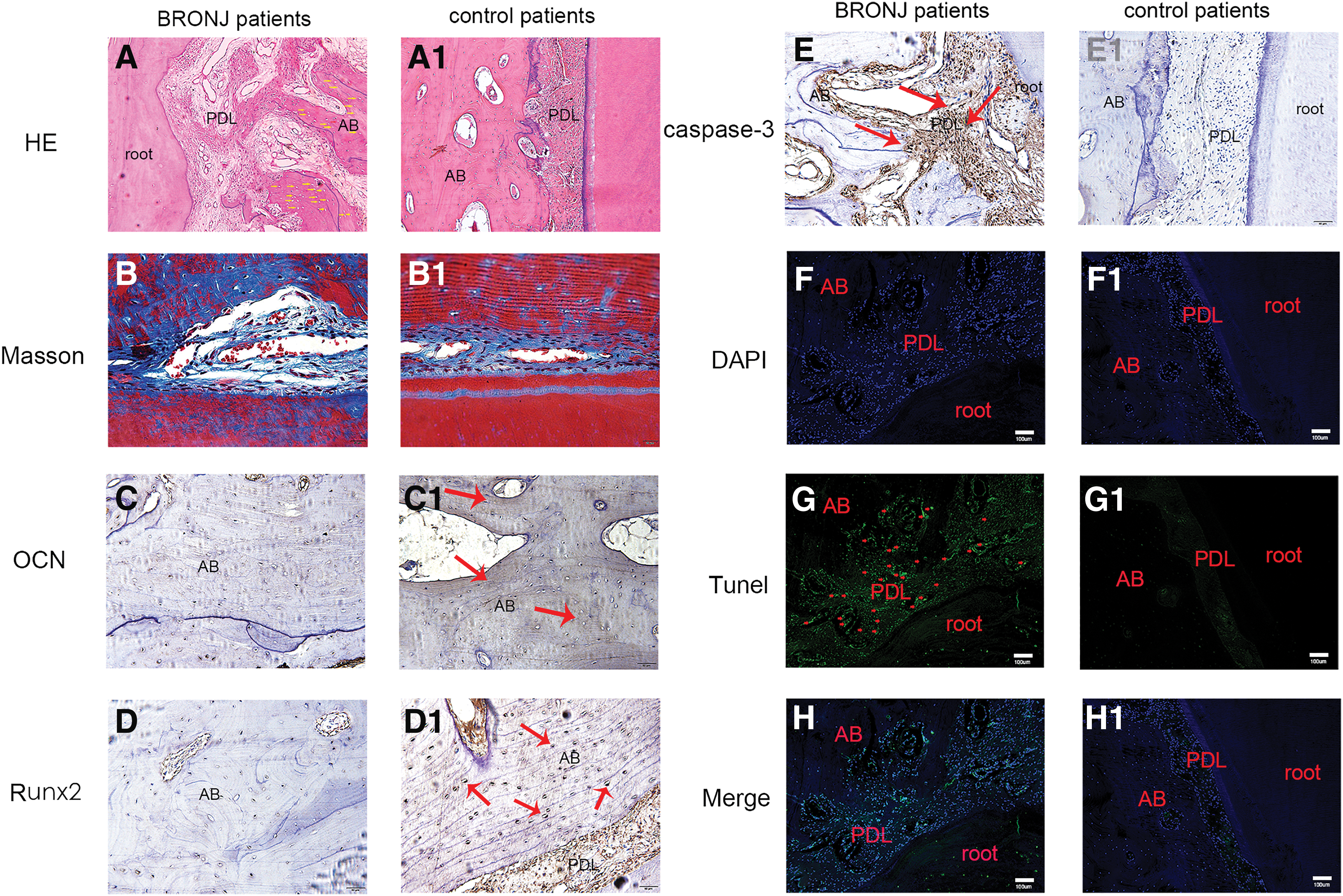
Histological analysis of the periodontal tissues in BRONJ patients and control patients.
In Fig. 2C and D, the images displayed the expression of OCN and Runx2 in the periodontal area of BRONJ patients, respectively. Figure 2C1 and D1 showed the expression of OCN and Runx2 in control patients. The data showed that the expressions of OCN and Runx2 in the periodontal area of BRONJ patients were weaker than the control. The results suggested that the periodontal area of BRONJ patients remarkably exhibited the decline of osteogenic abilities. To further explore the mechanism of the decline of osteogenic ability of periodontal tissues, we added experiments to verify the effect of zoledronic acid on the apoptosis of hPDLSC in BRONJ patients.
The results of caspase 3 immunohistochemical staining are shown in Fig. 2E and 2E1. Caspase 3, as the apoptotic executioner, is a member of the interleukin-converting enzyme-like proteases, which are related to mammalian apoptosis. We observed increased expression of caspase 3 in periodontal tissue of BRONJ patients compared with controls, especially in PDL (Fig. 2E, red arrows). The results showed that there was remarkable apoptosis of PDLSCs in a BRONJ patient, which may be related to the decline of periodontal osteogenesis. The apoptosis of hPDLSC in BRONJ patients (Fig. 2F–H, red arrows) and control patients (Fig. 2F1–H1) was also detected using TUNEL assay, the result further suggests that there is more apoptosis in PDL cells, which may also lead to the decrease of osteogenic ability.
Isolation and characterization of hPDLSCs from BRONJ and healthy PDL tissue
hPDLSCs were successfully isolated from BRONJ and healthy PDL tissue. These cells were found growing around the PDL tissues 5–10 days after the initial incubation, and the passage 0, 1, and 3 of BRONJ-hPDLSCs and control-hPDLSCs are shown in Fig. 3A, in which these cells exhibited typical fibroblastic morphology in vitro. However, there was no significant difference between the two groups. The flow cytometry results (Fig. 3B) showed that these cells were positive for STRO-1, CD90, and CD146, but negative for CD34 and CD45.
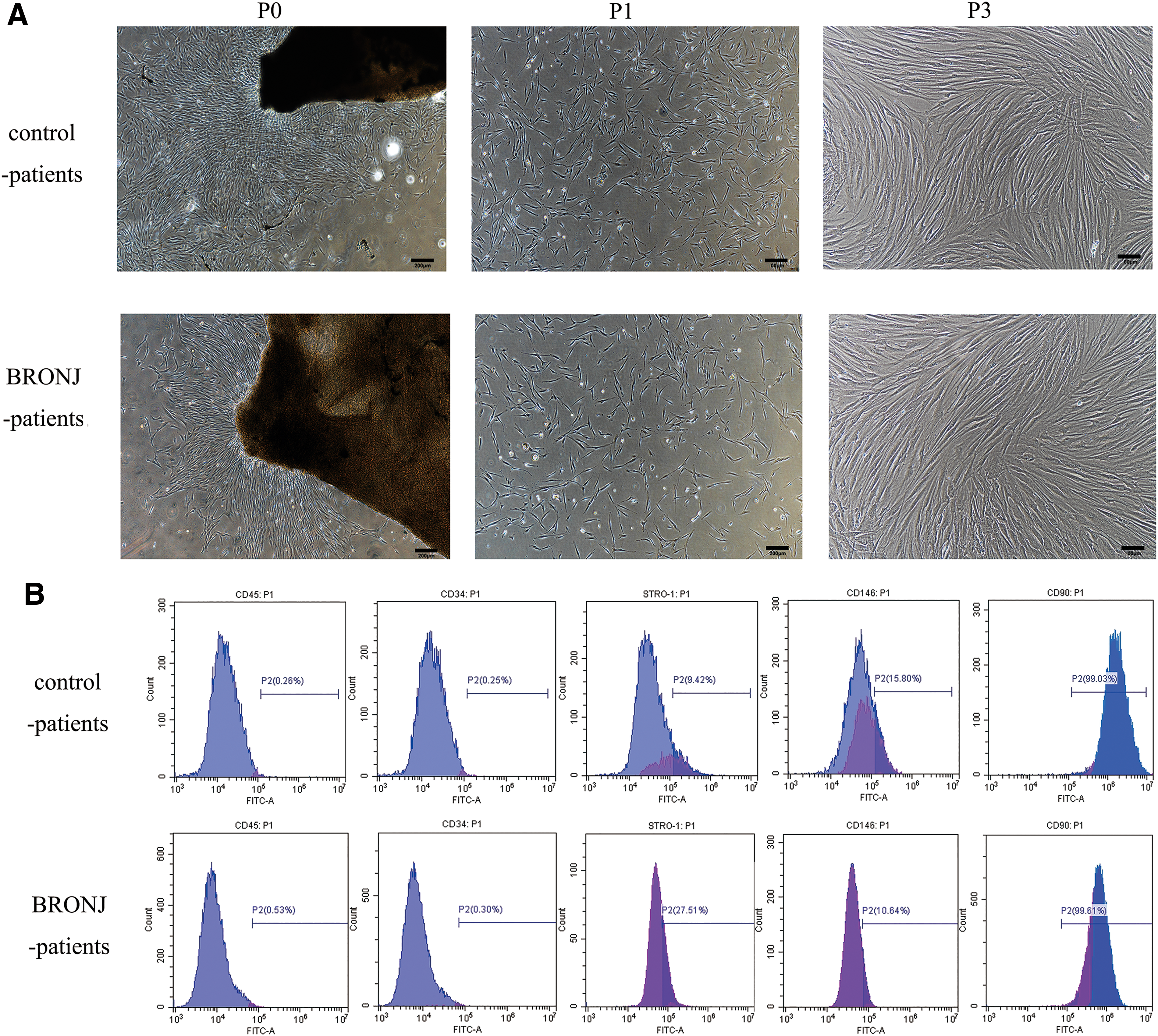
Isolation and identification of hPDLSCs derived from BRONJ patients and control patients.
Proliferation, adhesion, and migration of BRONJ-hPDLSCs and control-hPDLSCs
To determine the proliferation capacity of cells, we performed time/course studies. The growth curves of BRONJ-hPDLSCs and control-hPDLSCs are illustrated in Fig. 4A. Compared with control-hPDLSCs, the proliferation of BRONJ-hPDLSCs showed no significant difference within 72 h. After 72 h, BRONJ-hPDLSCs showed a lower proliferation rate than control-hPDLSCs. We also investigated the capacity of cell adhesion and migration. As evident from this analysis (Fig. 4B), the number of adherent cells was remarkably diminished in BRONJ-hPDLSCs compared with control-hPDLSCs. Figure 4C shows direct microscopic observation of cell migration. As shown in Fig. 4D, the wound healing capacity of BRONJ-hPDLSCs exhibited no significant differences at 6 h compared with control-hPDLSCs (the average migration area in control-hPDLSCs: 17.67% ± 1.08% vs. BRONJ-hPDLSCs: 17.43% ± 0.68%). At 24 h, the migration of BRONJ-hPDLSCs was significantly diminished (the average migration area in control-hPDLSCs: 84.15% ± 1.88% vs. BRONJ-hPDLSCs: 51.42% ± 2.53%). These results suggested that the proliferation, adhesion, and migration of BRONJ-hPDLSCs were remarkably decreased.
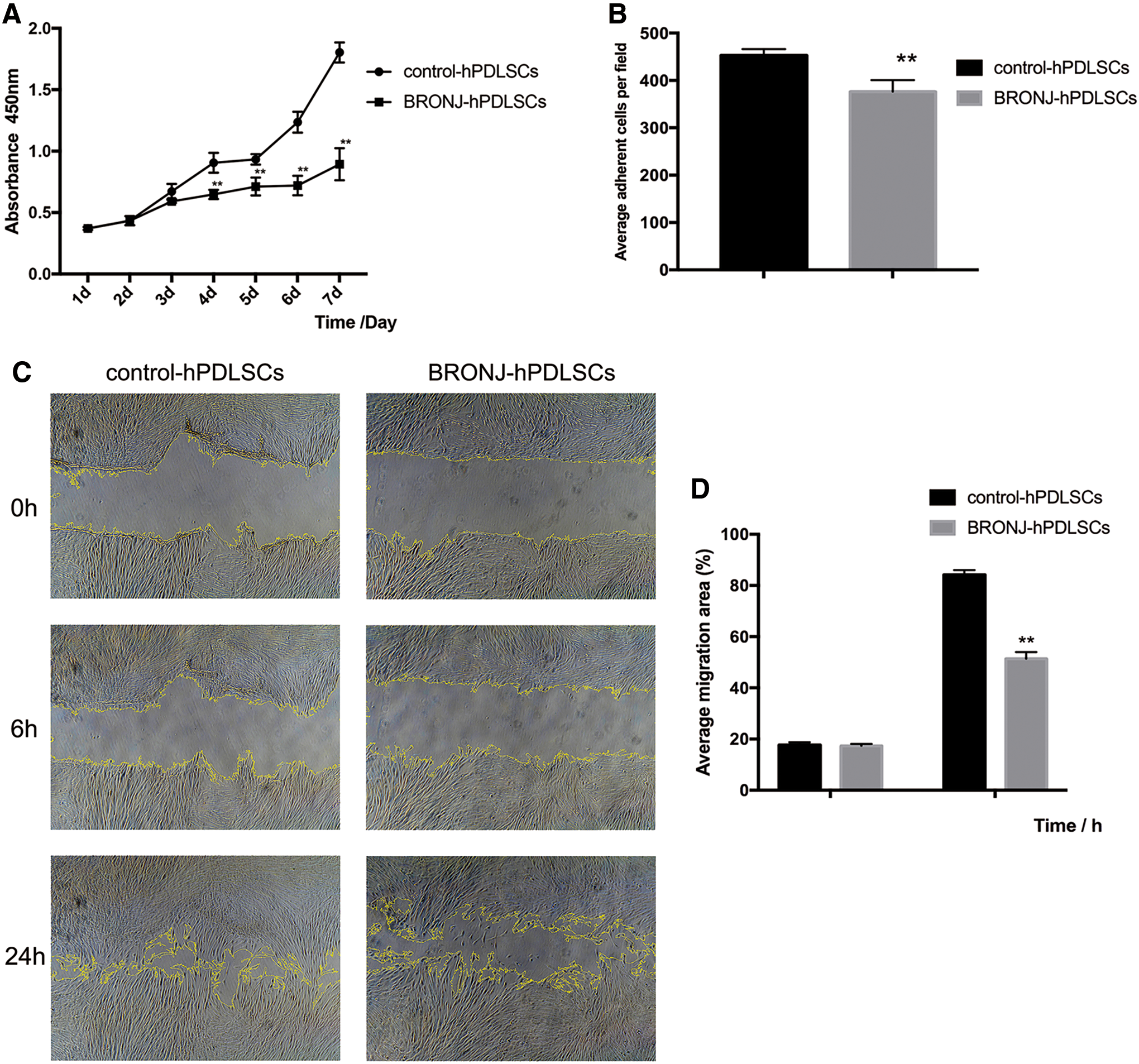
The assessment of the capacities of cell proliferation, adhesion, and migration among BRONJ-hPDLSCs and ZOL-hPDLSCs compared with control-hPDLSCs.
Cell apotosis in BRONJ-hPDLSCs and control-hPDLSCs
Supplementary Fig. S1 (the data are not shown) showed that 14.75% of hPDLSCs from the BRONJ group were positive for the early stages of apoptosis, but fewer apoptotic cells (2.14%) were found in the control group. We demonstrated that BRONJ-hPDLSCs underwent early apoptosis compared with control-hPDLSCs.
In vitro osteogenic differentiation capacity of BRONJ-hPDLSCs and control-hPDLSCs
hPDLSCs can differentiate into osteoblasts. To determine the capacity of BRONJ-hPDLSCs in osteogenic differentiation, we investigated ALP and Alizarin Red staining. Both ALP staining at 14 days and Alizarin Red staining at 21 days (Fig. 5A) showed that BRONJ-hPDLSCs expressed lower ALP levels and formed fewer calcium depositions than control-hPDLSCs in vitro. The quantitative data for ALP activity are shown in Fig. 5B, and Alizarin Red release shown in Fig. 5C also indicated consistent results.
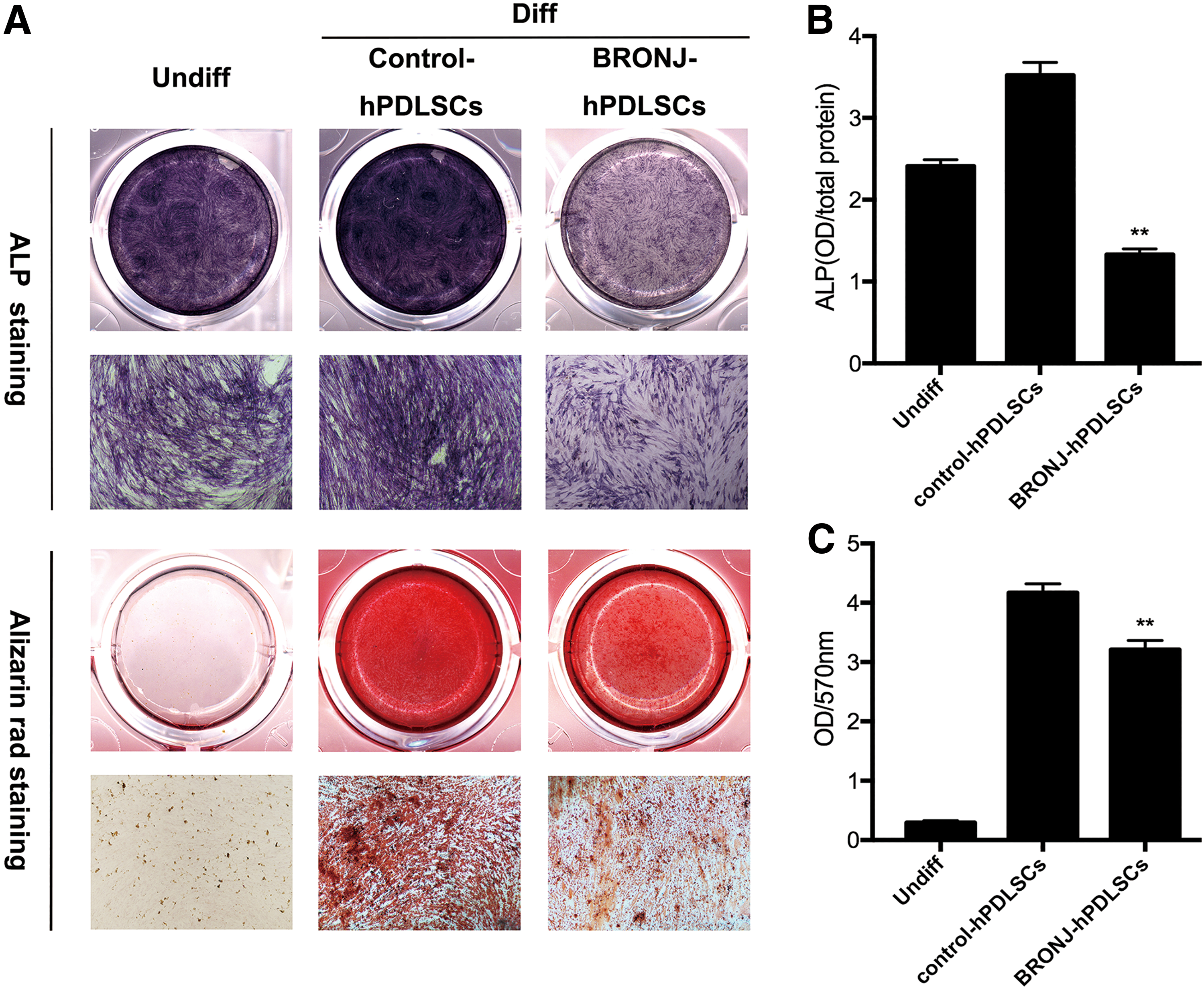
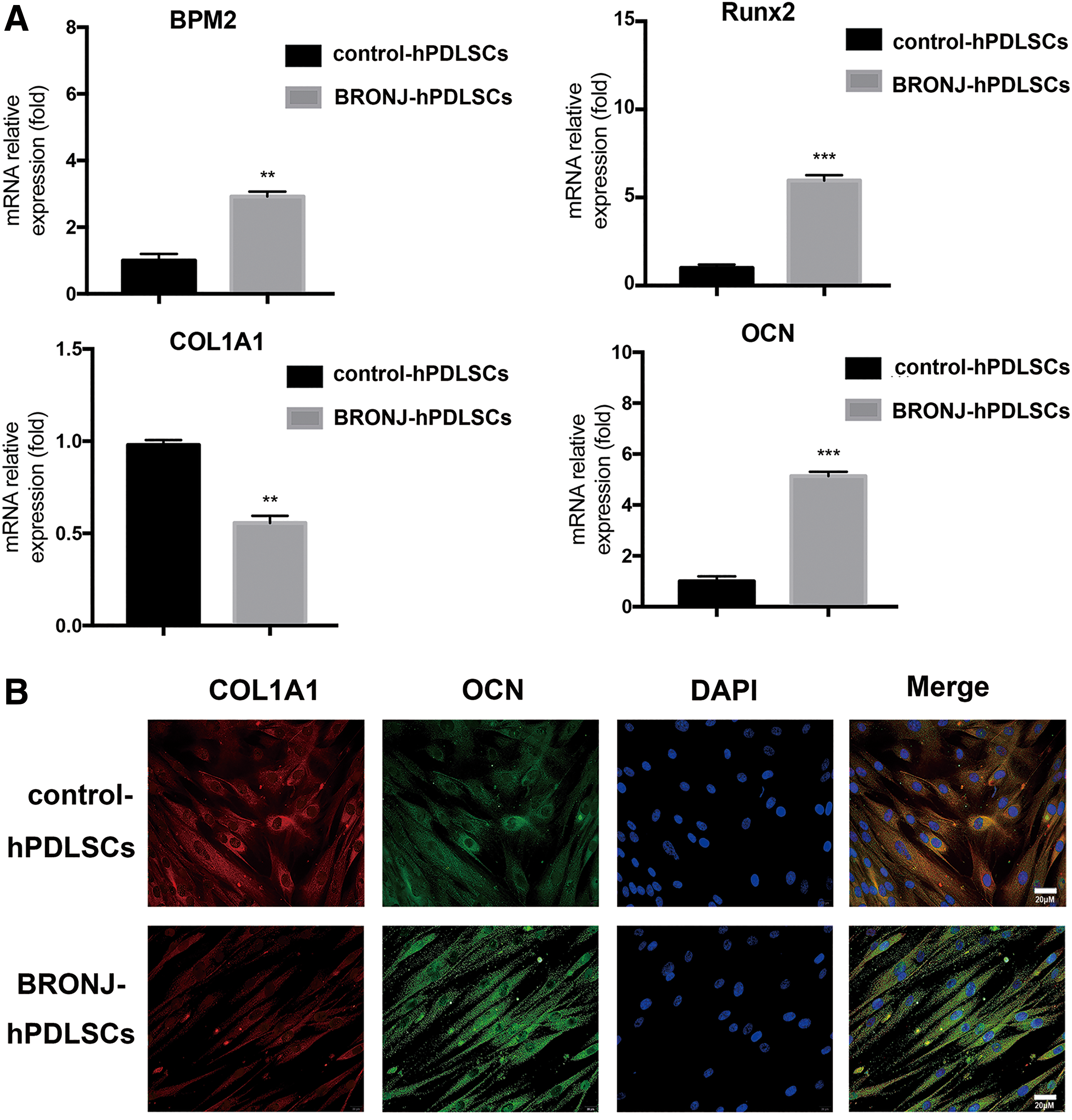
We further investigated the expression of the osteogenesis differentiation-related genes BMP2, OCN, Runx2, and COL1A1 in BRONJ-hPDLSCs. As shown in Fig. 6A, the mRNA expression of BMP2, OCN, and Runx2 was progressively upregulated in BRONJ-hPDLSCs, whereas the level of COL1A1 was remarkably decreased compared with control-hPDLSCs. Fluorescence images (Fig. 6B) showed that BRONJ-hPDLSCs expressed higher OCN protein levels but showed the lower expression of COL1A1 than control-hPDLSCs during osteogenic differentiation.
Comparative ectopic osteogenesis in nude mice
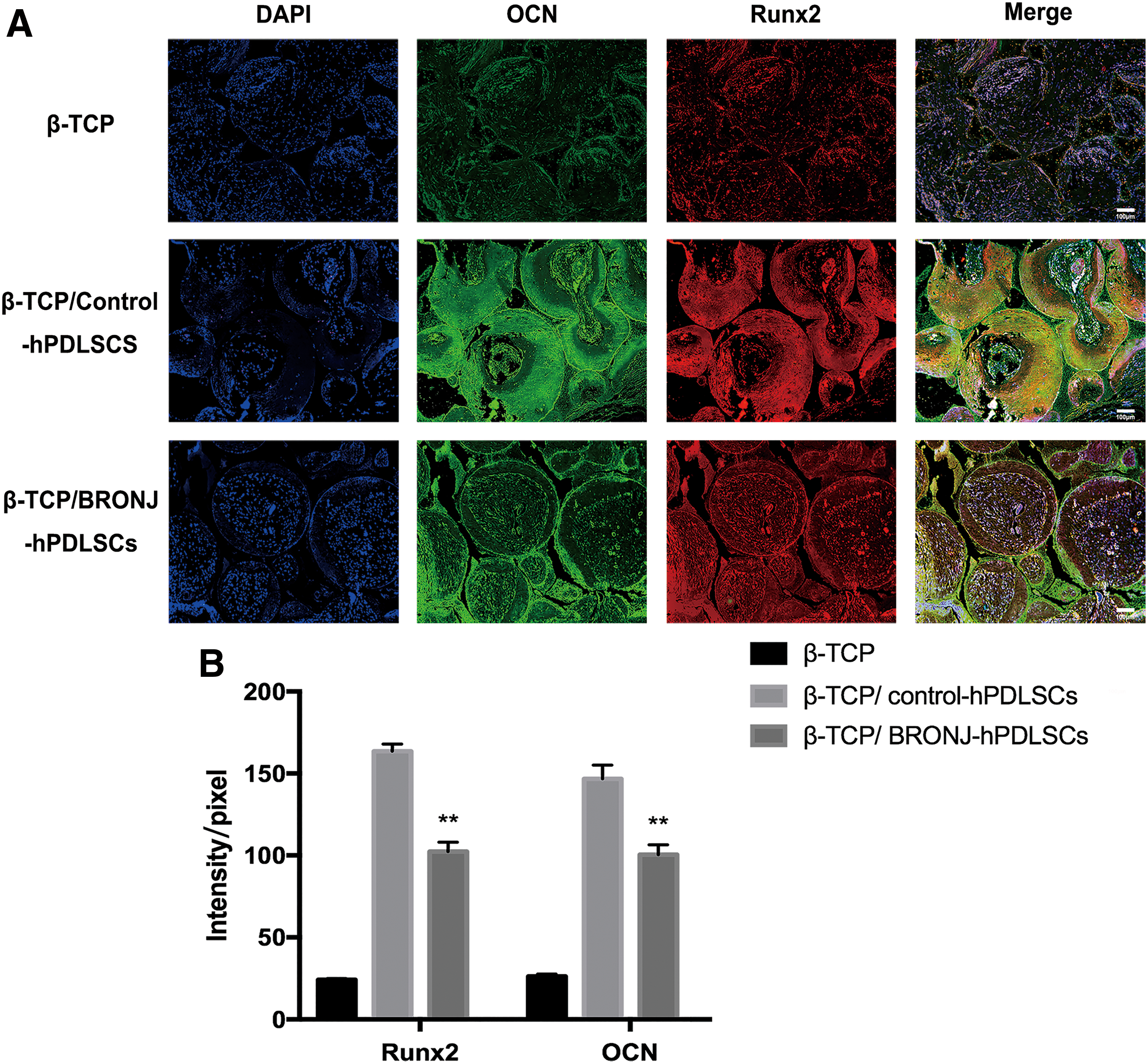
To further investigate cell osteogenesis in vivo, we quantitated new bone formation and the expression of osteogenic marker in three groups. Histological examinations were performed in six groups (Fig. 7A). The relative area of new bone formation was 58.19% ± 1.90% and 16.99% ± 1.62%, for Group B and Group C, respectively (Fig. 7B). There was no bone formation for β-TCP alone (Group A). Immunofluorescence images detected the expression of the osteogenic markers OCN and Runx2 (Fig. 8A). The quantitative data (Fig. 8B) showed that the expressions of OCN and Runx2 in Group C were weaker than Group B. Group A displayed the weakest expression of both proteins. Thus, the results above suggested that BRONJ-hPDLSCs exhibited remarkably impaired osteogenic differentiation abilities in vivo.

Analysis of new bone formation by β-TCP combined with hPDLSCs in nude mice.
Discussion
In this study, we first reported early changes of periodontal tissue in BRONJ patients and evaluated the activity of hPDLSCs in BRONJ lesions and their osteogenic capability, as well as examining periodontal tissue changes in vitro and in vivo. We also first demonstrated the role of impaired BRONJ-hPDLSCs in deficient periodontal repair of BRONJ. We assessed the activities of hPDLSCs derived from BRONJ lesions and found that BRONJ-hPDLSCs had consistently decreased cell proliferation, adhesion, migration, and osteogenic differentiation compared with control-hPDLSCs. BRONJ-hPDLSCs also underwent early apoptosis compared with control-hPDLSCs. In addition, we observed that new bone formation was remarkably decreased in BRONJ-hPDLSCs using an ectopic transplantation model. These results might help explain the poor periodontal defect repair in BRONJ lesions and provided new insights into the underlying mechanism of BRONJ.
To date, a major unanswered question is the reason why BRONJ mainly occurs on the jaw and rarely other bones [31 –33]. As we all know, the oral cavity organ is a special system composed of teeth, oral mucosa, periodontal tissues, and alveolar bone that is susceptible to periodontal disease, pulpal infections, and procedures such as dental extractions [14,34]. The unique structure of the maxillofacial region has been suggested as a contributing factor for the predisposition and progression of BRONJ [35]. Many reports have noted that BRONJ occurs after extraction of teeth due to the severity of periodontal or periapical disease [36,37]. Numerous researchers have also proposed that periodontal disease may predispose patients to developing BRONJ [5,38,39].
Thumbigere-Math et al. found that in humans treated with BPs, similar subclinical alveolar bone necrosis may be present around teeth with severe periodontal disease, periapical disease, or deep caries when compared with controls [14]. Consistent with Thumbigere-Math et al., in this study, we observed abnormal periodontal changes in BRONJ patients, including expanded PDL space, severely malformed PDL fibers and extreme periodontal bone destruction, which may be the early changes of periodontal tissues in BRONJ. Normally, PDL is soft, specialized connective tissue with high metabolic activity situated between the cementum covering the root of the tooth and the bone forming the socket wall, supporting the teeth, and at the same time permitting them to withstand the considerable forces of mastication [40,41].
hPDLSCs play an important role in the physiological maintenance and regeneration of periodontal tissues due to their ability to differentiate into cementoblasts, osteoblasts, and fibroblast cells [25]. Previous studies demonstrated that adjacent nonbone cells, such as oral keratinocytes and gingival fibroblast cells, contacting with a BP-coated bone surface might experience growth inhibition if contact is prolonged [9,42,43]. It is reasonable that high concentrations of BP in the jaw might produce a similar effect on hPDLSCs. Therefore, we speculated that the poor periodontal repair at BRONJ lesion sites in patients might be related to the impaired BRONJ-hPDLSCs.
In the current study, we first isolated hPDLSCs from BRONJ lesions and demonstrated that the proliferative rate of BRONJ-hPDLSCs dramatically decreased, and the adhesion and migration capacities were lower than those of control-hPDLSCs. Because cell adhesion and migration play pivotal roles in many physiological processes, such as tissue repair and stem cell regulation, cells can lose their normal functions if these processes are disrupted [30,44]. It was reported that hPDLSCs possessed low immunogenicity and have profound immunomodulation functions. Several experimental models [45,46] and clinical studies [47] have demonstrated associations between immune and BRONJ. BPs increase production of acute inflammatory mediators in vitro and in vivo and modify the immunological and cellular profile of BROJN patients [48,49].
However, the immunological properties of hPDLSCs in BRONJ patients have not been reported. Hence, we further explored the important behavior of the important aspect, which is the immunological properties in BRONJ-hPDLSCs, and we found that BRONJ-hPDLSCs underwent early apoptosis compared with control-hPDLSCs. We believe that the suppression of BRONJ-hPDLSC's proliferation, adhesion, migration, and cell apoptosis would be accompanied by serious damage to PDL homeostasis, leading to severe periodontal breakdown. The multipotential capacity of hPDLSCs, particularly osteogenic differentiation, plays an important role in periodontal tissue repair and regeneration.
To further examine whether the extreme periodontal bone destruction was relevant to the impaired BRONJ-hPDLSCs, we explored their osteogenic differentiation ability by ALP assays, Alizarin Red staining, immunofluorescence, and RT-PCR. BRONJ-hPDLSCs showed a dramatic inhibitory effect on osteogenic differentiation compared with control-hPDLSCs, as evidenced by the decrease in cellular ALP levels and the formation of calcium nodules. Surprisingly, the mRNA expression of BMP2, OCN, and Runx2 was upregulated in BRONJ-hPDLSCs, and the protein level of OCN was increased, whereas COL1A1 showed strongly decreased mRNA and protein compared with control-hPDLSCs. Consistent with our results, Kim et al. noted that COL1A1 associated with gingival tissue was significantly downregulated in BRONJ-like lesions [50].
However, some researchers reported that ZOL positively affected osteoblast differentiation and increased COL1A1 expression, which is inconsistent with our results [51 –53]. Intriguingly, the high expression of BMP2, OCN, and Runx2 did not produce continuous enhancement of ALP activity and Alizarin Red staining, as we expected. Conversely, we found that the levels of ALP and Alizarin Red staining in BRONJ-hPDLSCs were significantly decreased, which was in line with the lower expression level of COL1A1.
To our knowledge, ALP is a critical early participant in osseous mineralization, COL1 is a necessary initiator of osteogenic differentiation in PDLSCs and a required substrate for mineralization to occur, and OCN is expressed only in mature osteoblasts [54]. Collectively, we have shown that the osteogenic gene expression, and ALP activity in early stage osteogenic differentiation of BRONJ-hPDLSCs, but the exact mechanisms underlying the gene expression and ALP activity changes were unclear; we speculated that an increase in OCN, Runx2, and BMP2 expression may reflect a positive feedback response to the effects of BPs, COL1 expression reflect longer-term extracellular matrix changes, and ALP level represent early osteogenic differentiation that support decreased osteogenic differentiation.
For the in vivo osteogenesis evaluation, we transplanted hPDLSCs into nude mice, which is the most commonly used animal model for osteogenesis studies [55]. According to our present results, the new bone formation rate and the expression of the osteogenic markers, OCN and Runx2 in BRONJ-hPDLSCs, were significantly decreased compared with control-hPDLSCs. These results suggested that the osteogenic capacity in vivo was significantly impaired in BRONJ-hPDLSCs. We believe that most hPDLSCs within the jaw might also be impaired in BRONJ patients, which might indirectly result in poor periodontal repair of BRONJ lesions.
For the underlying mechanism of BRONJ, based on our results, we infer that most hPDLSCs might be impaired because of the high BP concentrations in the jaw, which might contribute to the malformed PDL and significant periodontal bone destruction in BRONJ patients. In this process, several factors must be emphasized. First, high doses of BPs remarkably inhibited hPDLSCs vitality and osteogenic ability. Second, the impaired hPDLSCs further caused serious damage to PDL homeostasis, leading to progressive periodontal damage. Third, inflammation resulting from periodontal defects may lead to more serious periodontal destruction.
In conclusion, our results above demonstrated that BRONJ-hPDLSCs showed decreased activities, with inhibited proliferative ability, reduced adhesion and migration capacities, and significantly impaired osteogenic differentiation ability, which can explain the severe periodontal destruction observed in BRONJ patients. We demonstrated that the negative effect of BPs on hPDLSCs plays an etiological role in BRONJ development.
Footnotes
Author Disclosure Statement
The authors declare no potential conflicts of interest with respect to the authorship and publication of this article.
Funding Information
This work was supported by the National Natural Science Foundation of China (no. 81271114) and Science and Technology Commission of Shanghai Municipality (19411962000 and 18441903000).
Supplementary Material
Supplementary Figure S1
Appendix
Polymerase Chain Reaction Primers and Conditions for the Specific Amplification of Human mRNA
| Genes | Primers sequences | |
|---|---|---|
| β-Actin | Forward | 5′-GGCACCACACCTTCTACAATGAGC-3′ |
| Reverse | 5′-GATAGCACAGCCTGGATAGCAACG-3′ | |
| OCN | Forward | 5′-CAGGCGCTACCTGTATCAATGGC-3′ |
| Reverse | 5′-GCCGATGTGGTCAGCCAACTC-3′ | |
| Runx-2 | Forward | 5′-AACAGCAGCAGCAGCAGCAG-3′ |
| Reverse | 5′-GCACCGAGCACAGGAAGTTGG-3′ | |
| COL1A1 | Forward | 5′-GGCGAGAGAGGTGAACAAGG-3′ |
| Reverse | 5′-GCCAAGGTCTCCAGGAACAC-3′ | |
| BMP2 | Forward | 5′-GACGTTGGTCAACTCTGTTAAC-3′ |
| Reverse | 5′-GTCAAGGTACAGCATCGAGATA-3′ | |
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.