
Abstract
The neural crest stem cells derived from human induced pluripotent stem cells (iPSC-NCSCs) are a valuable autologous cell source for tissue engineering and regenerative medicine. In this study, we investigated how iPSC-NCSCs could be regulated to regenerate arteries by microenvironmental factors, including the physical factor of matrix stiffness, and the chemical factor of transforming growth factor beta-1 (TGF-β1). We found that, compared to soft substrate, stiff substrate drove iPSC-NCSCs differentiation into smooth muscle cells, which was further enhanced by TGF-β1. To investigate the regulatory role of TGF-β1 in vivo, we fabricated vascular grafts composed of electrospun nanofibrous scaffolds, collagen gel, iPSC-NCSCs, and TGF-β1, and implanted them into athymic rats. The results showed that TGF-β1 significantly promoted extracellular matrix synthesis and increased mechanical strength of vascular grafts. This study presents a proof of concept that iPSC-NCSCs can be used as a promising autologous cell source for vascular regeneration when combined with physical and chemical engineering.
Introduction
The recent prominent discovery of induced pluripotent stem cells (iPSCs), which are reprogrammed from adult somatic cells, has shown promise for use in the field of regenerative medicine [1,2], and there is evidence that iPSCs can serve as an advanced cell source for vascular engineering [3 –7]. The biophysical properties such as the stiffness and micro-/nanotopography play important roles in regulating cell functions and tissue remodeling [8 –10]. Neural crest stem cells (NCSCs) derived from iPSCs are multipotent, and can differentiate into both mesenchymal and neural lineages, which represent a valuable cell source for tissue engineering and blood vessel regeneration in vitro and in vivo [11,12]. However, the in vivo differentiation of NCSCs in the vascular niche has yet to be explored.
Transforming growth factor beta-1 (TGF-β1) has been implicated in the maintenance of self-renewal and pluripotency of both human and mouse stem cells. The addition of TGF-β1 to the defined serum-free medium for human iPSCs supported the robust proliferation and maintained pluripotency of iPSCs [13]. It has been reported that human mesenchymal stem cell (MSC) differentiated toward myogenic lineage by regulating substrate stiffness or combining with TGF-β1 administration [8]. iPSC-derived smooth muscle cells (SMCs) treated with TGF-β1 can be induced into mature SMCs in vitro and the implantation of the SMC-scaffold construct in vivo demonstrated vascular tissue formation [14]. Although matrix stiffness is an important determinant of stem cell differentiation, its effect may not be specific for only one lineage, and biochemical factors such as TGF-β1 are required, together with matrix stiffness, to define a unique differentiation pathway.
In this study, we developed a biodegradable nanofibrous scaffold, integrating physical (stiffness) and chemical (TGF-β1) stimuli, to fabricate a novel cellular vascular graft with iPSC-derived NCSCs. iPSC-NCSCs were used to determine the differentiation commitment in response to various stiffness of collagen gel and TGF-β1 treatment in vitro. In addition, collagen-cellular vascular scaffold with TGF-β1 was designed as a stem cell differentiation platform to investigate the potential of iPSC-NCSCs for vascular engineering in vivo.
Materials and Methods
Cell culture and characterization
Undifferentiated human iPSCs were used to derive NCSCs as described previously [15]. NCSCs derived from iPSCs were maintained in StemPro® NSC serum-free neural induction medium (SFM; Invitrogen) for expansion without differentiation. Expanded NCSCs were characterized by expression of neural crest markers (p75, HNK1, vimentin, and nestin) and transcriptional factors (slug, AP2). NCSCs differentiation into mesenchymal lineages (SMCs, adipocytes, osteoblasts, and chondrocytes) and neural lineages (Schwann cells and peripheral neurons) was carried out and confirmed using the protocol described previously [12].
For all experiments, NCSCs with 70%–80% confluency were used. Unless specified, experiments in vitro were performed in the cell culture medium of Dulbecco's modified Eagle medium (Invitrogen) supplemented with 10% fetal bovine serum (Invitrogen) and 1% penicillin/streptomycin (Invitrogen), in the absence or presence of TGF-β1 (10 ng/mL, PeproTech).
Fabrication of substrates with different stiffness
Collagen-I gel (1 mg/mL; BD BioSciences) was prepared by neutralizing collagen-I solution with NaOH as described previously [8]. The soft substrate had collagen-I gel with about 500 μm thickness. For stiff substrates, collagen-I gel solution (before polymerization) was used to rinse the culture dishes, and excess solution was removed, resulting in a thin layer (microns) coating of polymerized collagen.
Immunohistochemistry analysis
NCSCs were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 15 min, followed by permeabilization with 0.5% Triton X-100 in PBS for 10 min. The specimens were incubated with respective primary antibodies for 2 h, and with appropriate secondary antibodies for 1 h. Nuclei were stained by 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen) in blue. The following antibodies were used: phalloidin (Invitrogen), Vinculin (Sigma-Aldrich), CNN1 (Epitomics), SMA (Sigma-Aldrich), SM22 (Abcam), MHC (Santa Cruz), CD31 (BD Biosciences), and NuMA (Abcam).
The fluorescently stained samples were imaged by using a Leica (Wetzlar, Germany) True Confocal Scanner SL confocal microscopy system, including He/Ne laser sources and a Leica DM IRB microscope, to capture multiple Z-section images (0.3- to 0.5-μm-thick sections) for a given specimen. The optical sections were subsequently projected to a single plane to create an overall image of the specimen. All images in a given group were collected with the same hardware and software settings. Other staining samples were examined using a Zeiss fluorescence microscope. The setting of microscopy system such as exposure time was kept constant for each experiment.
Scaffold fabrication
Poly
Upon completion of electrospinning, the vascular scaffolds were air-dried on the mandrel collector for two nights to remove residual HFIP. The scaffolds were cut into 7-mm length segments. All scaffolds were sterilized with ethylene oxide gas before characterization and in vivo implantation studies. The alignment of nanofibers and the structure of the nanofibrous vascular scaffolds were examined by scanning electron microscopy. The diameter of nanofibers ranged from 500 to 800 nm.
Tissue-engineered grafts and in vivo implantation
Upon completion of electrospinning, the nanofibrous scaffolds were sterilized by ethylene oxide gas sterilization and divided into three experimental groups: (1) grafts filled with collagen-I gel (diluted with SFM at a 1:2 ratio) without cells (Acellular), (2) grafts seeded with NCSCs (Cellular), and (3) grafts seeded with NCSCs in the presence of TGF-β1 (10 ng/mL) (Cellular+TGF).
To be specific, NCSCs were detached by TrypLE Select (Invitrogen) and resuspended in the SFM (2 × 104 cells/μL). The cell suspension was mixed with a cold collagen-I solution at 2:1 ratio (volume to volume) in the absence or presence of TGF-β1 (10 ng/mL), and then injected into the space around the tubular scaffold in a casting tube (eg, 1 mL for one scaffold 1.2 cm in length). The collagen-cellular scaffolds were kept in the incubator at 37°C for more than an hour, and NCSC maintenance medium was added to cover the scaffolds. The culture was maintained in the incubator overnight before surgery.
All animal procedures were approved by the Institutional Animal Care and Use Committee (IACUC) at UC Berkeley and were carried out according to the institutional guidelines and National Institutes of Health Guide for Care and Use of Laboratory Animals. Eighteen adult athymic rats (National Cancer Institute) weighing 200 ± 20 g were used in experiments, six animals for each group.
The left common carotid artery (CCA) on the ventral side of the neck was exposed and the segment of the artery was clamped temporarily. The CCA was cutoff 5 mm and collagen-cellular scaffold was used to place end-to-end to the CCA, suturing with 10-0 interrupted stitches under the surgical microscope. The muscle layers were approximated with interrupted 4-0 nylon sutures and stainless steel wound clips were used to close the skin wound. Retrieval of the graft involved the same initial steps for implantation. The graft was removed by ligation of native CCA directly adjacent to the suture locations.
Histological analysis
The vascular grafts were explanted and fixed in 4% paraformaldehyde at 4°C. Cross sections in the middle portion of the graft (10 μm in thickness; 5–7 mm from the proximal end of the graft) were cryosectioned for hematoxylin and eosin (H&E) staining, Verhoeff's staining (Sigma-Aldrich), and immunostaining, respectively.
Mechanical testing
Preimplanted grafts and freshly explanted vascular grafts were cut so that 2-mm-long segments could be mounted onto and uniaxially loaded by Instron mechanical testing machine. The stress-strain curve was obtained for each sample using accompanying Bluehill software. The elastic modulus was calculated from the linear portion of the stress-strain curve acquired for each graft.
Atomic force microscopy measurement of matrix stiffness
Animals were sacrificed following 2 weeks, 1 month, and 3 months of vascular graft implantation. Vascular graft samples were removed from the animal and cryopreserved using optimal cutting temperature (OCT). Samples were frozen for at least 24 h before cryosectioning. Samples were then sectioned into 90-μm slices, mounted onto microscope slides, and allowed to dry for 1 h. Sections were rehydrated using PBS and washed three times to dissolve the OCT compound. Samples remained hydrated throughout atomic force microscopy (AFM) testing. Force displacement profiles were created using silicon nitride cantilevers with pyramidal tips in contact mode. Cantilever spring constants were measured using thermal calibration to avoid error due to manufacturing discrepancies. Each tissue area was probed 10 times to avoid inconsistencies due to tissue heterogeneity. A customized MATLAB program was then used to apply the Hertz model to the data and determine the elastic modulus of the samples.
Statistical analysis
Mean and standard deviation values were calculated for each group of data. Analysis of variance was performed to detect whether a significant difference (P < 0.05, n ≥ 3) existed between groups. The Holm t-test was used to identify any differences. Student's t-test was used to analyze experimental groups with two samples. For the data with the normalization to the respective controls, log-transformed t-test was used to determine whether there was significant difference.
Results
Matrix stiffness and cytoskeleton organization in vitro
Expanded iPSC-derived NCSCs were characterized by the expression of NCSC markers (P75, nestin, vimentin, HNK1) and the capability of differentiation into mesenchymal lineages (SMCs, adipogenic cells, osteogenic cells, chondrogenic cells) and neural lineages (peripheral neurons, Schwann cells) in our previous work [9].
Collagen-I solution was applied for fabrication of stiff and soft (1 kPa) substrates [8], and phalloidin staining (Fig. 1a, d) and vinculin staining (Fig. 1e, h) were performed to determine the effects of substrate stiffness on cytoskeleton organization. Phalloidin staining of actin cytoskeleton demonstrated that NCSCs on stiff substrates exhibited extensive stress fibers (Fig. 1a), while cells on collagen gels (as soft as a 1-kPa substrate) had less spreading, fewer stress fibers, but more filopodia (Fig. 1c). TGF-β1 induced thicker stress fibers in NCSCs on stiff substrates (Fig. 1b) but not on gels (Fig. 1d). Large focal adhesions were only detected in NCSCs on stiff substrates in the presence of TGF-β1 (Fig. 1e, h), suggesting that TGF-β1 accelerated cell adhesion and spreading.
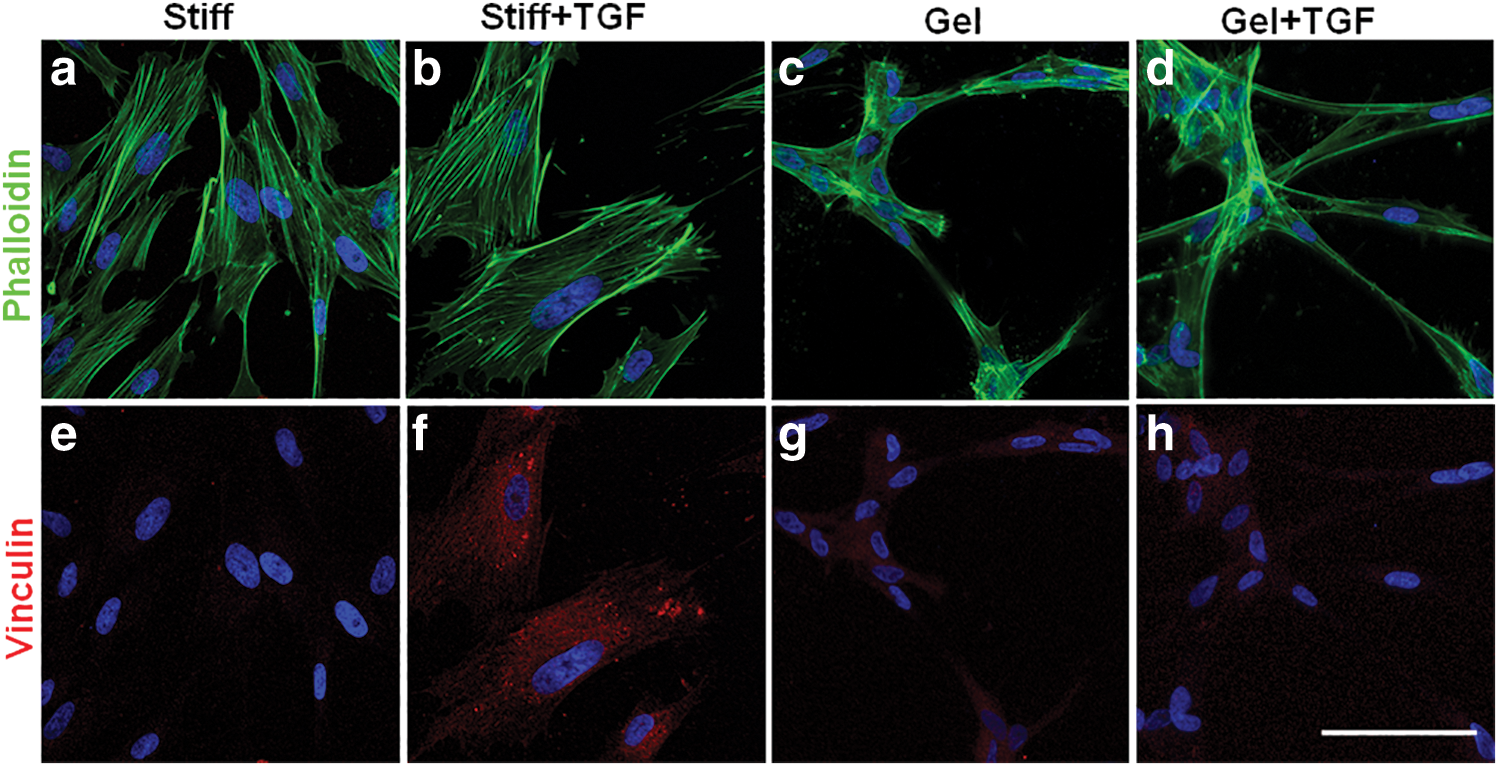
Effects of collagen gel and TGF-β1 on NCSC morphology. NCSCs were cultured in medium containing 10% fetal bovine serum in the absence
Effect of matrix stiffness and TGF-β1 on lineage specifications in vitro
To determine the differentiation commitment, NCSCs were cultured on collagen-coated culture dishes or collagen gels in the absence or presence of TGF-β1 (10 ng/mL) for 2 weeks. According to the immunostaining results shown in Fig. 2, without TGF-β1 treatment, SMA expression is much higher in the NCSCs on stiff substrate than those on soft substrate, however, other SMC makers, including CNN1, SM22, and MHC, were weakly expressed in the NCSCs on both groups, suggesting that stiffness can promote SMC differentiation to some extent. With TGF-β1 treatment, the expression of all the SMC markers examined in this study was significantly promoted in the stiff group. However, the effect was modest in the soft group. These results suggest that both TGF-β1 and stiffness are required to obtain maximal differentiation of NCSCs.

Effects of collagen gel and TGF-β1 on NCSC differentiation into SMC lineage. NCSCs were cultured on collagen-coated culture dishes (Stiff, Stiff+TGF) or collagen gels (Gel, Gel+TGF) in the absence (Stiff, Gel) or presence (Stiff+TGF, Gel+TGF) of TGF-β1 (10 ng/mL) for 2 weeks. Cells were immunostained for smooth muscle markers CNN1
In general, these striking differential effects suggested that substrate stiffness played an important role in the divergence of lineage specification into SMC lineage, and TGF-β1 further enhanced the difference on the way of specific differentiation.
Characterization of vascular grafts before and after in vivo implantation
Electrospun nanofibrous vascular scaffold (Fig. 3a) with an inner diameter of 1 mm and the thickness of 100 μm (Fig. 3b) comprised a luminal region of longitudinally aligned nanofibrous layer (Fig. 3c) and an outer region of circumferentially oriented layer (Fig. 3d). Generally, the aligned lumen surface facilitated the migration and proliferation of endothelial cells, and the outer layer stabilized the three-dimensional structure and enhanced extracellular matrix (ECM) remodeling and mechanical property of the scaffolds as described previously [9].

Characterization of vascular scaffolds before and after implantation.
Two experimental groups of vascular grafts (Cell and Cell+TGF-β1) were implanted into rat CCA by anastomosis (Fig. 3e). After 3 months, the grafts were explanted and cryosectioned for immunostaining. As shown in Fig. 3f, a tight outer layer of the graft was surrounded by NCSC-containing tissue via H&E staining, and the luminal surface was covered by endothelial cell layers via CD31 staining, moreover, positive CD31 was also present in the surrounding tissue outside the graft. These findings indicated that some microvessels have developed in the NCSC-wrapped layer and provided nutrition and oxygen for further ECM remodeling.
Differentiation of NCSCs into SMCs in the vascular grafts in vivo
To confirm SMC lineage differentiation, we demonstrated the expression of mature SMC marker MHC at different time points (2 weeks, 1 month, 3 months) by immunohistochemistry. As shown in Fig. 4, there was weak MHC expression in both Cellular and Cellular+TGF grafts at 2-week time point (Fig. 4a, b). At 1 month (Fig. 4c, d) and 3 months (Fig. 4e, f), it was obvious that Cellular+TGF group expressed more MHC than Cellular group in the fibrous capsule layer, which was consistent with previous data that the implanted NCSCs differentiated into SMC in the stiff layer [9] and TGF-β1 further promoted this process.
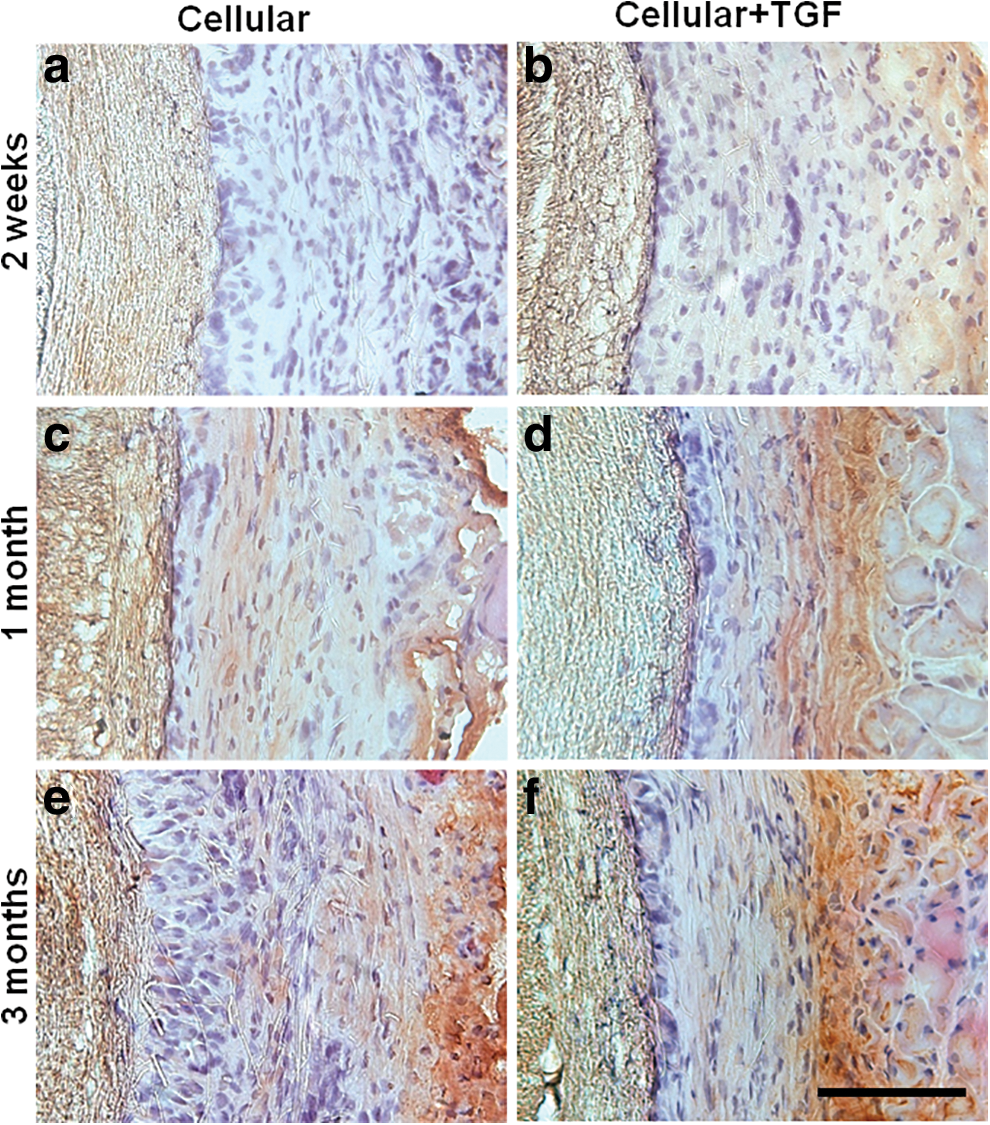
H&E and MHC staining of vascular grafts with NCSCs in the absence (Cellular) or presence (Cellular+TGF) of TGF-β1 after 2-week
To assess NCSC differentiation into SMCs in vascular grafts, the cross sections of the explants at different time points were stained for human antigen NuMA and SMC marker MHC (Fig. 5). At 3 months, more NuMA+/MHC+ cells were found in the outer layer of Cellular+TGF grafts than those of Cellular grafts (Fig. 5g, h), suggesting TGF-β1 enhanced the specific differentiation of NCSCs into SMC lineage.
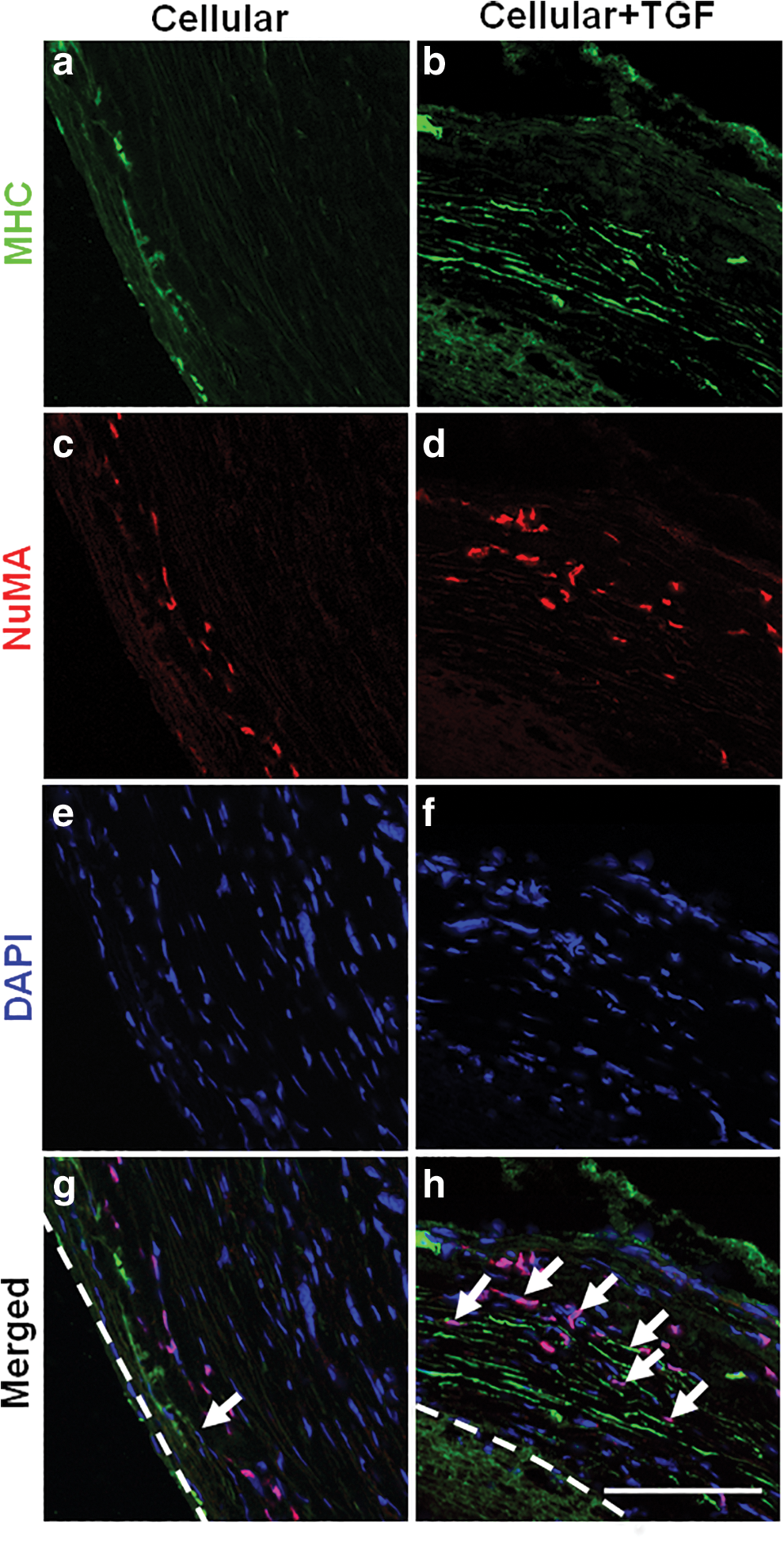
In vivo differentiation of NCSCs into SMC lineage. After 3 months of implantation, immunofluorescence staining of SMC marker MHC
ECM remodeling and mechanical properties of vascular grafts in vivo
Since ECM remodeling is a critical factor for the long-term stability of vascular grafts, we further investigated the synthesis and remodeling of ECM in the wall of vascular grafts. The cross sections of explanted grafts were stained for elastin, a main structural ECM component in the native artery. By Verhoeff's staining, we found that Cellular+TGF group had more elastin in the capsule layer close to the graft than Cellular group (Fig. 6a, b).
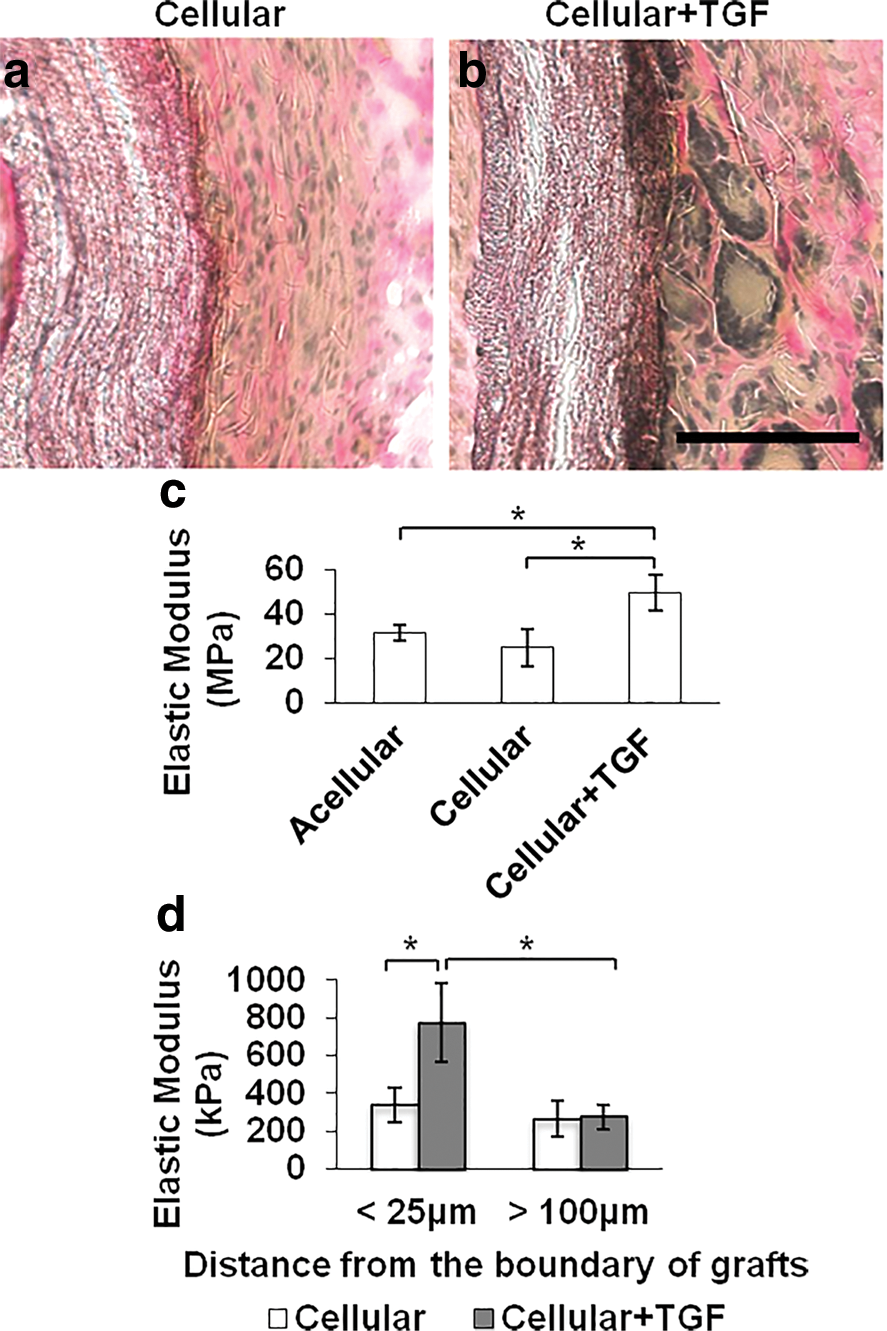
Extracellular matrix remodeling, elastic modulus of vascular grafts, and matrix stiffness around the grafts in vivo.
Instron mechanical test showed that Cell+TGF grafts had similar elastic modulus to native artery, which was much higher than those of acellular and cellular grafts (Fig. 6c). This result suggested that TGF-β1 significantly promoted MHC and elastin expression in the capsule layer and strengthened the mechanical property of vascular graft in Cellular-TGF group.
AFM measurement of matrix stiffness around the polymer grafts
Our previous AFM measurement revealed that elastic modulus of both polymer scaffold and the native carotid artery had similar stiffness (∼6,000 kPa) [9]. In this study, AFM measurement of the outer layer of vascular grafts presented that Cellular group had similar stiffness across the whole layer (Fig. 6d), however, Cellular+TGF group had much higher elastic modulus in the region close to graft boundary (<25 μm), but much lower in the region far away (>100 μm). This is consistent with the results in vitro that the combination of stiffness and TGF-β1 contributed to the maximal SMC differentiation (Fig. 2).
Discussion
The difficulty in vascular replacement can be attributed to multiple factors, including the limited availability of venous and arterial grafts and the lack of appropriate mechanical properties, chemical stimuli, and cell recruitment in synthetic vascular grafts. Most efforts to improve the strength and remodeling of synthetic vascular scaffolds have been directed toward fabricating the ideal biocompatible and biodegradable materials and providing appropriate biochemical environment and cell sources to enhance cellular organization [16 –18]. Our in vitro studies using collagen gel showed that NCSC differentiation was regulated by substrate stiffness, and presented that matrix stiffness alone can promote a particular lineage over another, that is, SMCs on stiff substrates, and neural cells on soft substrates, which was consistent with in vivo differentiation of NCSCs [9]. In the vascular wall where biochemical and hemodynamic microenvironmental factors are similar in the neighborhood of ∼100 μm, the differentiation of NCSCs into SMCs happened and was well distributed in the surrounding tissue of the polymer scaffold, where stiffness is gradient from inner area (∼340 kPa) to outer (∼260 kPa) in 100 μm.
Recent reports suggested that matrix rigidity and chemical stimuli might work synergistically to direct specialized cellular differentiation and fate determination [8,19,20]. To further investigate the integrative effect on NCSCs differentiation, our study wrapped vascular graft in cellular collagen gel with TGF-β1 administration. Interestingly, our results demonstrated significant expression of SMC markers and elastin deposition in the fibrous layer with TGF-β1 treatment compared to that of no TGF-β1 treatment group. The TGF-β family regulates a diversity of cellular functions during development and homeostasis through their effects on cell migration, proliferation, and differentiation [21]. Recent findings have reported that TGF-β1 signaling regulated mechanotransduction to promote MSC differentiation into SMC lineage on stiff substrate [8]. Those are also in line with our findings that TGF-β1 facilitated the organization of SMCs and ECM remodeling, as well as results showed that the stiffness close to the polymer graft (<25 μm) was increased two times after TGF-β1 treatment, suggesting TGF-β1 works together with matrix stiffness to further stimulate the differentiation of NCSCs into SMCs. Meanwhile, our in vitro study using collagen gel confirmed that NCSC differentiation was also regulated by the substrate stiffness. It was noteworthy that soft surface induced NCSC differentiation into glial genes, whereas MSCs were committed to the chondrogenic lineage [8,9]. Moreover, the combination of TGF-β1 and stiffness strengthened this NCSC differentiation process, which provided a mechanistic and biomimetic explanation of the findings in vivo.
Our study presented a one-step electrospinning process to fabricate a novel, seamless, and tubular nanofibrous vascular scaffold [9,15,22], which was composed of two fully integrated layers: a luminal layer with longitudinally aligned nanofibers and an outer layer with circumferentially organized nanofibers. The biomimetic micro-/nanostructure of the scaffold, although it is not the focus of this study, may benefit for vascular graft remodeling. Longitudinally aligned fibers in the lumen mimicked the structure of native artery and may promote endothelial cell attachment and migration to maintain patency of vascular grafts. The circumferential nanofibrous layer may facilitate the organization of SMCs and ECM remodeling to improve the mechanical properties. According to previous investigation, the designed polymer scaffold had a similar stiffness to native artery and this mechanical compliance matching was appropriate for vascular graft fabrication. In addition, natural polymers such as collagen and elastin were applied for vascular grafts construction with favorable mechanical properties and cellular integration [23]. Thus, the combination of synthetic polymer PLLA and collagen gel with stem cells may provide a synergistic effect on biomechanical strength and structural remodeling.
Phalloidin staining of actin cytoskeleton demonstrated that NCSCs on stiff substrates exhibited extensive stress fibers. TGF-β1 induced thicker stress fibers in NCSCs on stiff substrates than soft ones, suggesting that TGF-β1 accelerated cell adhesion and spreading. Accordingly, in the presence of TGF-β1, the differentiated SMCs in vivo showed both stretched cytosol and nuclear morphology and expressed significant MHC marker. Interestingly, those implanted cells might incorporate host cells to develop the smooth muscle layer to enhance mechanical strength. Moreover, the wrapped collagen gel could be conductive to endothelial cell attachment and migration, and increased microvessel formation may encourage the local circulation to provide abundant nutrition and oxygen for cell infiltration and organization. Altogether, these results provide compressive perception of the interconnections between biochemical and biophysical factors in directing stem cell differentiation in vivo, which will significantly promote the development of stem cell transplantation in vascular graft engineering and regenerative medicine application.
Conclusion
Overall, this study demonstrated that nanofibrous scaffolds offered a valuable platform of cellular therapies for vascular graft replacement in addition to the guidance of SMC organization. Scaffold/gel-mediated NCSC implantation with TGF-β1 treatment not only induced in vivo differentiation to smooth muscle lineage but also advanced the mechanical properties. The results have imperative implications for the potential of iPSC-derived NCSCs in autologous regenerative therapies and also for understanding the physical effects of the in vivo vascular microenvironment.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financially supported in part by Chongqing Medical University (NSFYY201822, X.L.), National Natural Science Foundation of China (U1601220, X.T.), Science and Technology Commission of Shanghai Municipality (13PJ1401300, Y.Z.), and Foundation of Shanghai Municipal Commission of Health and Family Planning (201640306, Y.Z.).