
Abstract
The objective of this study was to determine whether corneal stromal cells (CSCs) from the limbal and central corneal stroma in dogs have multipotent mesenchymal stem/stromal cell (MSC) properties, and whether this cell population can be differentiated into keratocyte-like cells (KDCs). Normal, donated, mesocephalic dog corneas were used to isolate CSC in vitro. Immunohistochemistry demonstrated a distinct population of CD90 expressing cells in the anterior stroma throughout the limbal and central cornea. CSC could be cultured from both the limbal and central cornea and the culture kinetics showed a progenitor cell profile. The CSC expressed stem cell markers CD90, CD73, CD105, N-cadherin, and Pax6, while CD34 was negative. Limbal and central CSC differentiated into osteoblasts, chondrocytes, and adipocytes confirming their multipotency. Coculturing allogeneic peripheral blood mononuclear cells (PBMCs) with limbal CSCs did not affect baseline PBMC proliferation indicating a degree of innate immune privilege. Limbal CSC could be differentiated into KDCs that expressed Keratocan, Lumican, and ALDH1A3 and downregulated Pax6 and N-cadherin. In conclusion, canine CSCs have multipotent MSC properties similarly described in humans and could serve as a source of cells for cell therapy and studying corneal diseases.
Introduction
The cornea is an optically clear tissue permitting light transmission into the eye and is essential for the biodefense and refractive system of the eye [1]. The basic anatomical structure of the cornea between mammals is similar. It is composed of an anterior epithelium and its basement membrane, the stroma, the Descemet's membrane, which is the basement membrane formed by the single-layered posterior endothelium. However, a distinct layer comparable to the anterior limiting lamina (Bowman's layer) that is present in the human cornea does not exist in dogs [2,3].
Dogs can suffer from inherited diseases affecting the cornea, such as corneal crystalline dystrophy and corneal fibrosis, which remain the leading causes of blindness in animals and people worldwide [4,5]. Blindness has severe consequences for working dogs and also negatively affects the quality of life in pet dogs. In contrast to human ophthalmology, corneal transplantation is rarely performed in dogs. This is due to a shortage of donors, expensive storage costs, and the lack of a canine corneal tissue bank [6].
Funderburgh et al. described a population of Pax6 expressing cells (<4%) in the bovine corneal stroma and identified them as progenitor cells. Stromal cells expressing mesenchymal stem/stromal cell (MSC) markers in the anterior stroma close to the limbus and central cornea were also identified in humans [7]. A recent publication has also described a stromal stem cell population that originates from the human central corneal stroma [8]. Corneal stromal cells (CSCs) of the limbus in humans were found to fulfill the minimal criteria of The International Society for Cellular Therapy (ISCT) for multipotent MSCs [9 –11].
In vitro, human CSCs that are also referred to as corneal stromal stem cells (CSSCs) were differentiated into keratocytes that expressed Lumican, Keratocan, and ALDH1A1 and lost expression of Pax6 and ABCG2 [12 –14]. A stable keratocyte phenotype has been described in relationship to the differentiation process [15].
Similar to many other multipotent MSCs, CSSC are described to have immunomodulatory properties, which has relevance for preventing graft rejection for the purpose of tissue engineering. An in vivo study in mice has shown lack of tissue rejection after injecting CSSC in a corneal fibrosis model [16]. A study using a mixed lymphocyte reaction with limbal CSSC showed an inhibition of the proliferation of activated peripheral blood mononuclear cells (PBMCs) [8].
To date, CSSCs are thought to primarily have a support function for limbal epithelial stem cells (LESCs). This is reinforced by the fact that basal epithelial cells in the limbal niche are in direct contact with stromal cells, both cells produce N-cadherin, a cell–cell junction protein and LESC cocultured with CSSC have higher expansion and clonogenicity rates [17,18]. However, they may also have a role in corneal tissue homeostasis as an ex vivo model culturing sheep corneas with seeded CSSC after a LASIK (laser in situ keratomileusis) procedure showed significantly increased adherence of LASIK-like flaps while maintaining corneal transparency [8,19].
The current literature is based mainly on human, murine, rabbit, and porcine corneal cells [20,21], but studies in canine corneas are not available This led to the aim of the study to locate and characterize CSCs in dogs and investigate whether they have multipotent MSC properties. This study will provide a baseline for researchers working with canine corneal disease models and for potential cell-based therapy in veterinary ophthalmology.
Materials and Methods
Donor material, selection, and exclusion criteria
Corneas were isolated from 14 healthy donor corneas of dogs euthanized for reason unrelated to this project and with the consent of the Animal Health Trust Ethical Review Committee (AHT_31_2013). Before excision of the cornealscleral buttons, the eyes were examined using a handheld slit lamp (KOWA SL14; Kowa Company Ltd., Nagoya, Japan) by a board-certified veterinary ophthalmologist (C.K.).
The mean age (±standard deviation, range) of corneal donors was 4.61 years (±3.97; median: 2; range: 0.2–12 years). There were eight different mesocephalic breeds that were included and brachycephalic breeds were excluded. The gender distribution was composed of intact males (n = 6), intact female (n = 1), spayed females (n = 4), and neutered males (n = 3). Details about the donor tissues are listed in the Supplementary Table S1.
Cultured CSCs of each donor were expanded, cryopreserved, and then randomly selected in the following experiments. For each experiment, ≥3 independent biological replicates were used (ie, cells derived from three or more different donor dogs).
Cell isolation and cell culture
Before excision, the eyes were examined using a handheld slit lamp (KOWA SL14; Kowa Company Ltd.) by a board-certified veterinary ophthalmologist (C.K.).
Within an average time of 8.28 ± 3.38 h after confirmed death, the cornealscleral buttons were excised. Left and right eyes were placed in separate containers containing Eagle's Minimum Essential Medium (Sigma-Aldrich, Gillingham, Dorset, UK), 5% dextran (Sigma-Aldrich), 2% fetal bovine serum (Gibco, Life Technologies, Paisley, UK), and 1% Antibiotic-Antimycotic mix (Sigma-Aldrich) at room temperature (RT) for an average time of 13.5 ± 6.38 (range: 6–21) days with a weekly change of the media.
The CSCs were isolated using a method similar to that described previously [12]. In brief, with magnification of 5.5 × surgical loupes (Keeler, Winsdor, UK), the superficial corneal limbal region was excised using a 300 μm set depth knife (BD, Beaver Visitec International, Bidford-Upon-Avon, Warwickshire, UK). The limbal anterior stroma, including the epithelium, was removed using corneal scissors and dissected into small fragments (3 × 3 mm). The central cornea was trephined using a 7.5 mm corneal handheld trephine (Altomed, Bolden, UK) and dissected into strips using a 300 μm set depth knife.
The limbal and central tissue fragments of each donor were processed separately and digested in 50% Dulbecco's modified Eagle's medium (DMEM) high glucose (Gibco), 50% DMEM/F12 (Gibco), supplemented with 50 μg/mL gentamicin (Gibco), 1% penicillin–streptomycin solution (Gibco), containing type L collagenase from Clostridium histolyticum (0.5 mg/mL; Sigma-Aldrich) and incubated at 37°C, 5% CO2 for 12–14 h. Selective trypsinization was used (TripLE express; Gibco) to separate the small oval shaped CSC from epithelial cells, keratocytes, and melanocytes. CSCs were expanded to a maximum confluency of 60%–70%. The medium was changed every 48–72 h.
CSC medium was composed of DMEM low glucose (Gibco) and MCDB-201 (Sigma-Aldrich) medium, 10 ng/mL epidermal growth factor (EGF; Sigma-Aldrich), 10 ng/mL platelet-derived growth factor (PDGF-BB; R&D Systems, Abingdon, Oxford, UK), Insulin-Transferrin-Selenium (Gibco), 0.1 mM ascorbic acid-2-phosphate (Sigma-Aldrich), 10−8 M dexamethasone (Sigma-Aldrich), 1% penicillin–streptomycin (Gibco), 50 μg/mL gentamicin (Gibco), 100 ng/mL cholera toxin (Sigma-Aldrich), supplemented with 2% fetal bovine serum (FBS; Invitrogen, Thermo Fisher Scientific, Paisley, UK) [15].
Population doubling time and population doublings
The mean population doubling time (PDT) was calculated from three biological independent limbal- and central-derived CSC lines [8-year-old Springer Spaniel (No. 10), 12-year-old Lurcher (No. 11), 12-year-old Golden Retriever (No. 12)] (Supplementary Table S1).The doubling time and number of cell doublings (NCD) were calculated at each passage until the stationary phase was reached (P1–P6; Roth V. 2006 Doubling Time Computing, available from:
The population doubling of cells was calculated as:
where y is the final density and x is the initial seeding density of the cells [10].
The cell number of P0–P1 was not included as CSC derived from cultured limbal and central cells had epithelial cell contamination.
Keratocyte differentiation of CSCs
Limbal- and central-derived CSCs (passage 2–5) of a 12-year-old Lurcher, (No. 11), 12-year-old Golden Retriever (No. 12), and a 2-year-old Border Collie (13) (Supplementary Table S1) were cultured in keratocyte differentiation medium (KDM) consisting of advanced DMEM (Sigma-Aldrich), 10 ng/mL human basic fibroblast growth factor (bFGF; Sigma-Aldrich), 0.1 mM
A seeding density of 1 × 103 cells/cm2 and one passage between 10 and 14 days were necessary to maintain stellate cell morphology, avoid cell confluency, and cell-sheet formation. A second culture experiment was conducted with the same media but lacking human bFGF (Sigma-Aldrich).
Cell culture of adipose-derived MSCs
Three lines of canine adipose-derived MSCs (adMSC) that had been derived previously were used as positive controls [11-year-old Mix breed (No. 15), 8-year-old Irish setter (No. 16), and 6 weeks old Dalmatian (No. 17)] (Supplementary Table S1) [22]. The cryopreserved MSCs were thawed into DMEM (Gibco) with 10% FBS (Gibco), 1% penicillin–streptomycin (Gibco) and 2 mM
Histology, immunohistochemistry, and immunofluorescence staining
Three corneas from biological independent donors [1.2-year-old (No. 1), 1.3-year-old (No. 2), and 1.5-year-old (No. 3) Staffordshire bull terrier ](Supplementary Table S1) were embedded in O.C.T. tissue compound (Tissue Tek®; VWR, Lutterworth, Leicestershire, UK). The tissue was separated in the dorsal, temporal, ventral, and nasal regions, snap frozen in liquid nitrogen, and cut in 7 μm thick frozen sections for routine automated hematoxylin-eosin and periodic acid-Schiff (PAS) staining. Immunohistological investigations were performed on each of the four indicated regions.
The sections were fixed in 3% paraformaldehyde for 20 min, washed in phosphate-buffered saline (PBS) and permeabilized in 0.1% Triton X-100 (Sigma-Aldrich) in PBS for 20 min.
Limbal and central CSCs, their differentiated keratocyte-like cells (KDCs) from three different donors [12-year-old Lurcher (No. 11), 12-year-old Golden retriever (No. 12), 2-year-old Border collie (No. 13)] (Supplementary Table S1), and adMSCs were plated onto Fibronectin (FNC coating mix; Athena Enzyme Systems,, Baltimore, MD)-coated permanox slides at a density of 4 × 104 cells/slide, cultured for a 2–3 days in CSC, MSC (at passage 2–5) or KDM, fixed with 3% paraformaldehyde for 20 min. For sections and fixed cells prepared for identification of keratocyte markers, 0.1% Tween 20 (Sigma-Aldrich) in PBS was used for washing procedures. Cells/sections were blocked with 10% goat/donkey serum (Sigma-Aldrich) in PBS for 60 min at RT. Details of primary and secondary antibodies are summarized in Supplementary Table S2.
In brief, the cells/sections were incubated with the primary antibody in 5% blocking serum [goat or donkey serum (both Sigma-Aldrich)] in PBS at 4°C overnight. Slides were washed in PBS and incubated with corresponding secondary antibody diluted in 5% blocking serum in PBS for 1 h at RT and fluorescein isothiocyanate (FITC)-labeled phalloidin (1:1,000 concentration; Sigma-Aldrich), which binds to the actin cytoskeleton, mounted in VECTASHIELD with the nuclear stain DAPI (Vector Laboratories, Inc., Peterborough, UK) and evaluated with a confocal microscope (Zeiss LSM 700, Cambridge, UK). Rabbit and Mouse IgG isotype controls were performed.
Flow cytometry
Flow cytometry was performed on three independent biological replicates of limbal CSCs [12-year-old Lurcher (No. 11), 12-year-old Golden retriever (No. 12), and 2-year-old Border collie (No. 13)] (Supplementary Table S1). Cells were fixed in 3% paraformaldehyde (Sigma-Aldrich) for 20 min before washing, permeabilized in 0.1% Triton X-100 for 1 h (except for CD90), and blocked in 10% goat or donkey serum (Sigma-Aldrich) for 30 min. Cells (1 × 106) were incubated for 45 min at 4°C with primary antibody, rabbit, or sheep IgG, followed by incubation with the appropriate FITC labeled secondary antibody for 45 min 4°C. Details of all primary and secondary antibodies used for immunostaining are listed in Supplementary Table S2.
A FACS Calibur flow cytometer (BD Biosciences Immunocytometry Systems, Franklin Lakes, NJ) was used and the data were analyzed using NovoExpress® Software (ACEA Biosciences, San Diego, CA). Events were gated to exclude dead cells and cell debris by forward versus side scatter height (R1). R1 events were gated to remove doublets (R2) by forward scatter height versus forward scatter area. Two percent of events in the R2 isotype control and everything to the right were included in a new gate (R3) by FITC height versus forward scatter height [23]. The overlaid events of each primary antibody gated within R3 were considered positive. The events were expressed as means of positive cells (%).
RNA extraction and quantitative reverse transcriptase polymerase chain reaction
Total RNA was extracted from corneal tissue, 1 × 106 limbal CSC (passage 4–5), and 1 × 106 of their KDC of three different donors [12-year-old Lurcher (No. 11), 12-year-old Golden retriever (No. 12), and 2-year-old Border collie (No. 13)] (Supplementary Table S1) using TRI-Reagent (Sigma-Aldrich), purified using the RNeasy Mini Kit (Qiagen, Germantown, MD) and treated with Ambion DNA-free (Life Technologies) to remove genomic DNA. Complementary DNA (cDNA) was made from 1 μg of RNA using the sensiFAST cDNA synthesis kit (Bioline, London, UK).
Aliquots of 2 μL cDNA were used in quantitative polymerase chain reaction (qPCR), which was carried out using SYBR Green containing supermix (Bioline) on the Bio-Rad C1000 Touch Thermal Cycler (Bio-Rad, Watford, UK). qPCR reactions were performed in duplicate. The cycle parameters were 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 15 s, and 72°C for 15 s. Finally, a melt curve was produced by taking readings every 1°C from 65°C to 95°C. The levels of canine 18S ribosomal RNA (rRNA) did not change between treatments (data not shown) and it was used to normalize gene expression using the 2−ΔΔCt method [24].
Primers were designed from annotated canine exon sequence for the genes of interest using primer3 (
Assays for adipogenesis, osteogenesis, and chondrogenesis
Central [donor: 6 weeks old Dalmatian (No. 8), 8 weeks old Old English Sheepdog (No. 9), 12-year-old Golden Retriever (No. 12), and 2-year-old Border Collie (No. 13)]-derived and limbal [(donor: 1-year-old Staffordshire crossbreed (No. 6), 8-week-old Old English Sheepdog (No. 9), 12-year-old Lurcher (No. 11), and 2-year-old Border Collie (No. 13)]-derived CSC and adMSC from three different donors of passage 2–5 (Supplementary Table S1) were differentiated into adipocytes, chondrocytes, and osteoblasts. The following modified protocols by Guest et al. were used [25].
For adipogenic differentiation CSC and adMSC were treated with antibiotic-free fat induction media [DMEM supplemented with 10% fetal calf serum, 15% rabbit serum (Invitrogen), 10 μg/mL insulin, 1 μM dexamethasone, 0.02 mM indomethacin, and 0.5 mM 3-isobutyl-1-methylxanthine (Sigma-Aldrich)] for 72 h and followed by antibiotic-free fat maintenance media (DMEM supplemented with 10% fetal calf serum and 10 μg/mL insulin) for 72 h for three alternating cycles. Oil red O staining for lipid droplets was then carried out.
For chondrogenic differentiation cells were treated with cartilage induction media [DMEM supplemented with 10% fetal calf serum, 2 mM
For osteogenic differentiation, cells were cultured for 21 days in osteogenic induction media [DMEM supplemented with 10% fetal calf serum, 2 mM
Western blot analysis
Antibodies without canine-specific references were tested for cross-reactivity and specificity to dog proteins using western blot. Details are in Supplementary Table S2.
Protein was extracted from whole-cell extracts (WCEs) from adMSC (P4), CSC (P4), and corneal tissues using 500 μL WCE buffer (5 mM EDTA, 5 M NaCl, 0.5 M Hepes pH7.9, 100 mM PMSF), followed by three freeze and thaw cycles. Supernatants were collected by centrifugation at 4°C and stored at −20°C.
To test the keratocyte markers Lumican and Keratocan, a deglycosylation step was performed using a combination of endo-β Galactosidase (Sigma-Aldrich) (0.57 U/mg protein for 4 h at 37°C) and Chondroitinase ABC (Sigma-Aldrich) (0.6 U/mg protein) for 4 h at 37°C) on 100 μg of corneal protein. Twenty micrograms of denatured protein was run either on a 10% or 5% sodium dodecyl sulfate (SDS)-polyacrylamide gel and transferred to a PDVF membrane. Immunoreactivity was detected using the ECL plus detection system (Amersham, Buckinghamshire, UK). Mouse anti-β actin antibody (Abcam) was used as a positive control.
PBMC media and isolation from lymph nodes
The rapid destruction of peripheral blood PBMCs through the use of commercial euthanasia agents prevented the isolation of PBMCs from the blood of the euthanized patients from the corneal donor study population.
Therefore, the popliteii lymph nodes of two donor dogs were harvested 2 h after euthanasia and transported on ice in PBMC media as described by Dutton et al. [26]. PBMC media was composed of RPMI 1630, 10% heat inactivated FBS, 1% penicillin–streptomycin (Gibco), 2 mM
In brief, lymph nodes were excised, cut into small pieces, and passed through a 70 μm sterile cell strainer (Sigma-Aldrich) followed by several washes. The cell suspension was centrifuged at 300 g for 15 min, and cells were frozen in liquid nitrogen a concentration of 4 × 106/mL in 90% PBMC media and 10% dimethyl sulfoxide (DMSO). After thawing, 77%–95% of PBMCs were viable.
Canine PBMCs of two dogs were commercially purchased (3H Biomedical, Uppsala, Sweden).
PBMC cocultures
CSCs were growth arrested with 15 μg/mL (2-h incubation) Mitomycin C (MMC; Sigma-Aldrich). PBMC cocultures were performed by incubating growth arrested, allogeneic CSC on a 96-well plate with effector PBMCs of 2 different donors in a ratio of 1:5 (CSC:PBMC). The experiment was performed on three biologically independent limbal CSCs from three different donors [donor: 1.3-year-old Staffordshire bull terrier (No. 2), 1.5-year-old Staffordshire bull terrier (No. 3), and 12-year-old Golden retriever (No. 12) (Supplementary Table S1), (ie, CSCs of three different donors were used with two different PBMC donors)]. For the negative controls, PBMCs were MMC treated for 30 min at a concentration of 50 μg/mL before being washed and cultured with autologous effector PBMCs.
For positive controls, growth arrested PBMCs were cultured with allogeneic effector PBMCs. In addition, PBMCs were activated with 10 μg/mL phytohemagglutinin (PHA; Sigma-Aldrich).
After 4 days, the PBMCs were incubated with radioactive thymidine ([3H] thymidine; GE Healthcare Bio-Sciences) at a final concentration of 0.5 μCi per well, at 37°C, 5% CO2 for 16–18 h before being frozen at −20°C. Following thawing, cells were harvested using a Filtermate and counted using TopCount NXT equipment.
Statistical analyses
Statistical analyses of the cell culture kinetics and gene expression data followed a normal distribution calculated by Shapiro-Wilk normality test (W) and P values >0.05 (XLSTAT-base, Witzenhausen, Germany). Group differences were calculated using an unpaired Student's t-test and P < 0.05 was considered to be statistically significant.
Results
The canine limbus has epithelial invagination and an undulating basement membrane. A distinct population of CSCs in the limbus and central stroma express the MSC marker CD90.
By histology, limbal crypts and palisades of Vogt (as in humans) were absent in canines (Fig. 1B), however, a slight invagination of the epithelium in the stroma was noted. In the canine corneal limbus, the basement membrane was undulated and irregularly formed. There was no evidence of regional differences (dorsal, ventral, nasal, temporal). In the limbus, the basal cell layer of the epithelium was cell rich with more elongated basal cell nuclei similar to LESCs. The CSCs were in direct contact or in close proximity to the irregularly formed basement membrane (Fig. 1B). A distinct, small population of these stromal cells expressed CD90 and extended into the mid-stroma of the limbal region in all four quadrants (Fig. 1B).
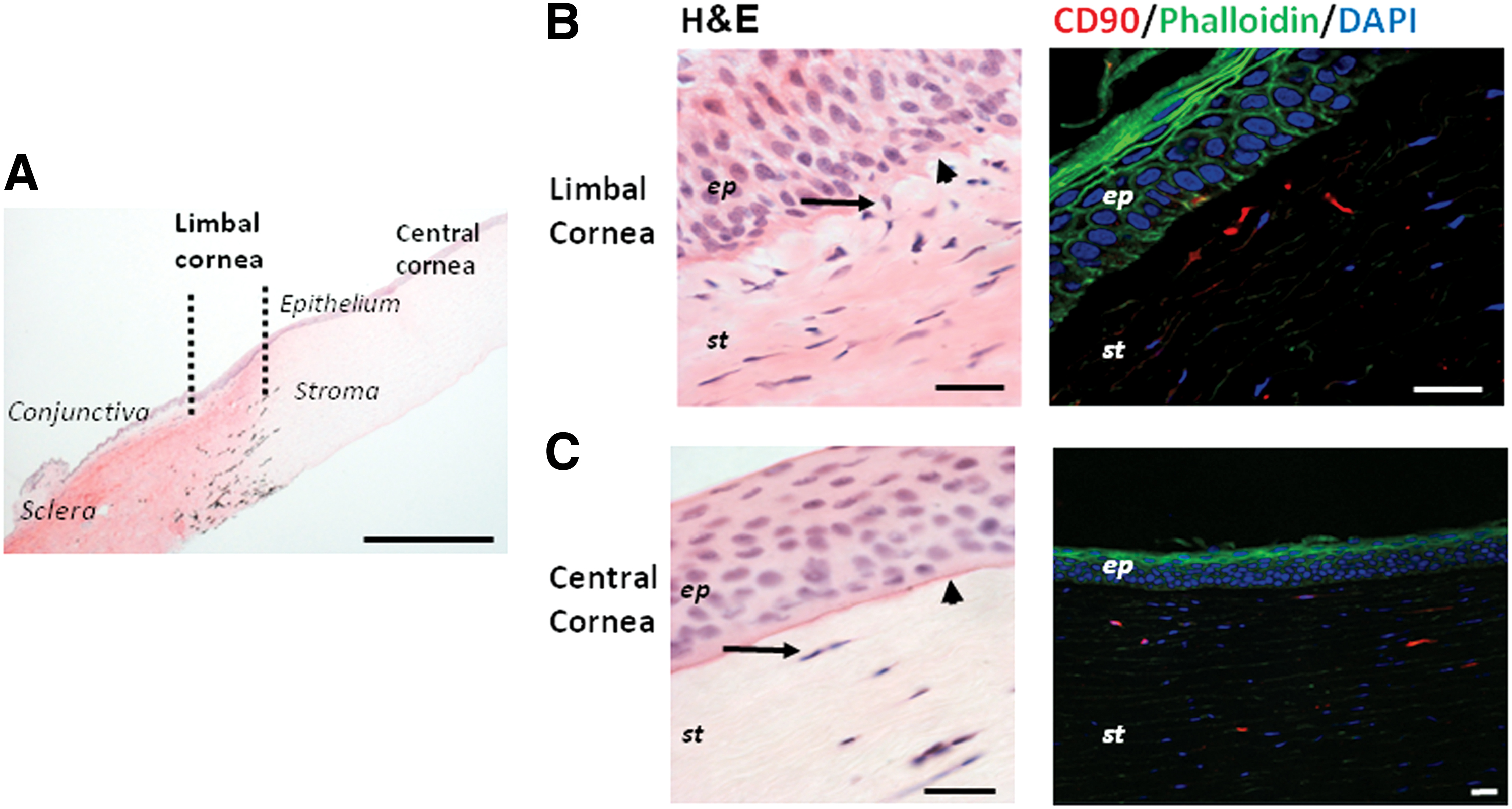
Histological and immunohistochemical staining of the canine limbal and central cornea with H&E and CD90.
In the central cornea, the basal cells of the epithelium were cuboidal in shape. The fine basement membrane was regularly formed and the stromal cells were separated from the basement membrane by a layer of corneal stroma (Fig. 1C). A distinct population of stromal cells expressing CD90 extended into the mid-stroma (Fig. 1C). Pax6 expressing stromal cells were not evident in the limbal or the central cornea.
CSCs can be cultured in vitro from both the limbal and central cornea but have a limited self-renewal
Adherent cells were visualized ∼6 h postseeding. The cells adopted an ovoid morphology after 24 h, but after the first passage, they showed a more spindle cell morphology and formed a syncytium of spindle cells with increasing confluency (Fig. 2A.i–iii).
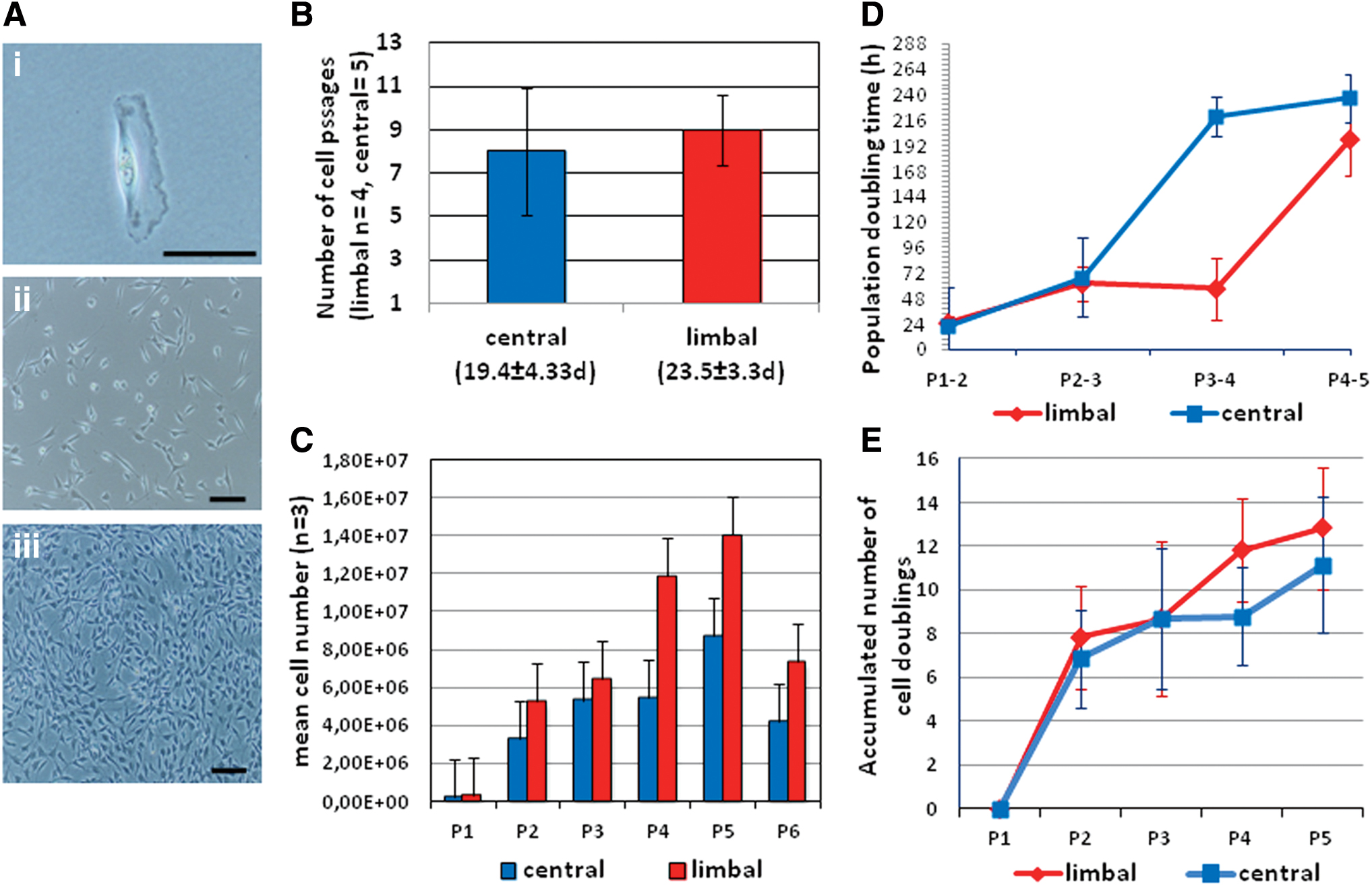
Cell morphology and culture kinetics of limbal versus central CSC.
The average maximum passage was 9 (±2.94) and 8 (±1.64) (P = 0.55) passages over a culture time of 23.5 (±3.31) and 19.6 (±4.33) days (P = 0.18) in limbal and central-derived CSCs, respectively (Fig. 2B). There were no statistically significant differences in cell numbers of the limbal and central CSCs over six passages [P = 0.54 (P1), P = 0.67 (P2), P = 0.83 (P3), P = 0.12 (P4), P = 0.49 (P5), P = 0.44 (P6)] (Fig. 2C).
The mean PDT between limbal- and central-derived CSC showed a trend that central CSC reached the stationary phase at an earlier passage (P3–4) than the limbal CSC, but the difference did not reach statistical significance (P = 0.068) (Fig. 2D). In the early log-phase, between P1 and P2, a PDT of 25.02 and 21.67 h of limbal- and central-derived CSCs, respectively, were noted, which increased exponentially and reached the stationary phase after five passages for both central and limbal CSCs (Fig. 2D). The accumulative NCD between P1 and P5 was 12.84 and 11.15 in limbal and central CSCs, respectively (Fig. 2E).
Canine CSCs have MSC characteristics
Cultured limbal- and central-derived CSCs expressed MSC markers. CSC showed strong expression of CD90 and CD73, whereas CD105 was less intensely expressed (Fig. 3). The nuclear paired box protein Pax6 (oculorhombin) and the intercellular membrane marker N-cadherin were expressed (Fig. 3). CD34, a hematopoietic stem cell marker and alpha smooth muscle actin (α-SMA), a myofibroblast (MF) marker were absent (Fig. 3).
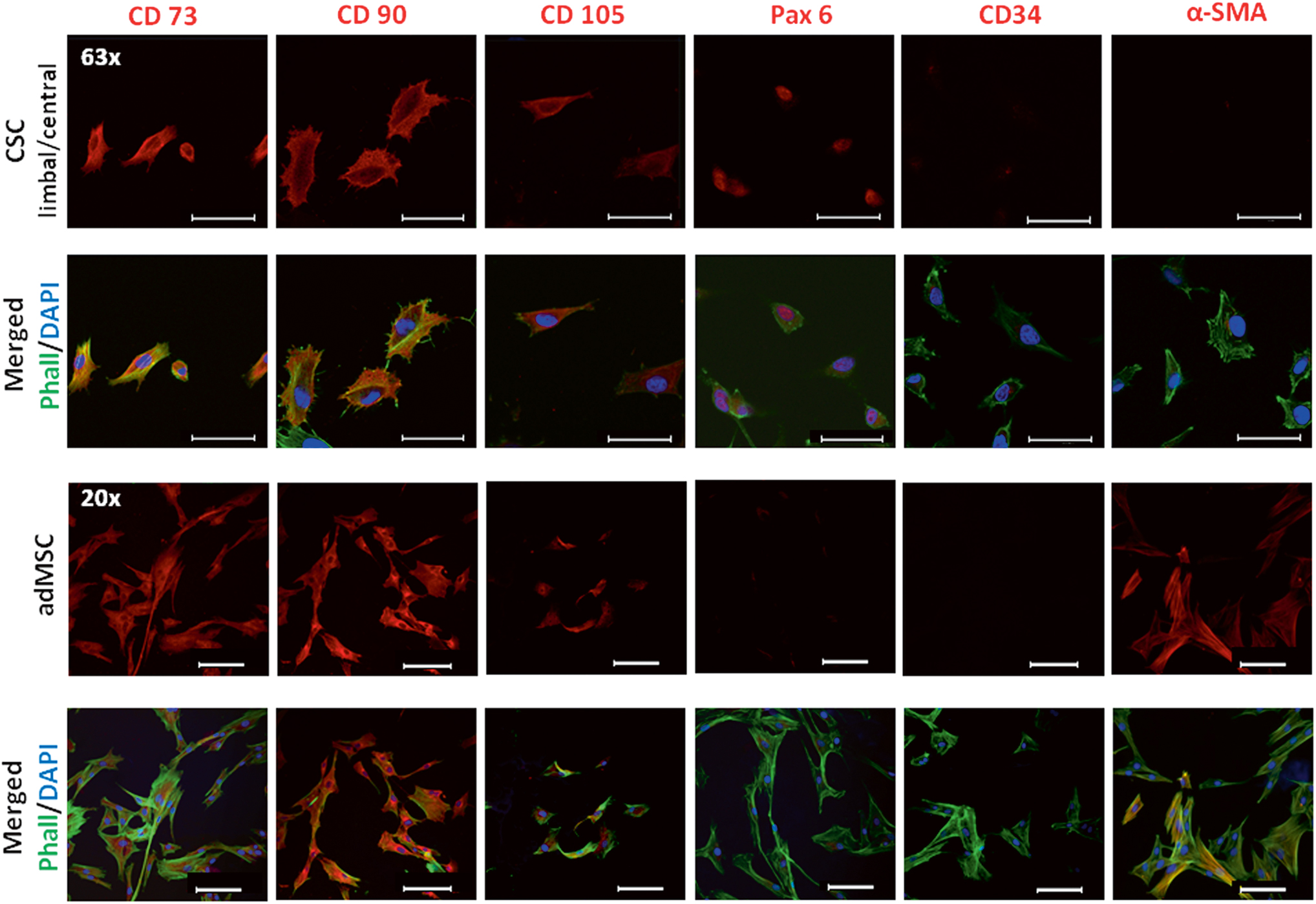
Immunocytochemical profile of CSC in comparison to canine adMSC: CSC and adMSC expressed CD73, CD90, and CD105, but not CD34; only CSC expressed Pax6 and only adMSC expressed α-SMA. Limbal and central CSCs showed the same staining profile. Nuclei are shown by DAPI counter staining and the cytoskeleton by Phalloidin (Phall). Scale bar: 20 μm; magnification: CSC (63 × ), adMSC (20 × ). α-SMA, alpha smooth muscle actin; adMSC, adipose-derived mesenchymal stem/stromal cell. Color images are available online.
Canine adMSCs were used as a positive control and showed a similar marker expression to CSCs, but lacked expression of Pax6 and CD34 and had strong expression of α-SMA (Fig. 3).
Flow cytometry revealed that 83% of CSCs expressed Pax6, 96% expressed CD90 and CD73, whereas 1% expressed CD34 (Fig. 4).
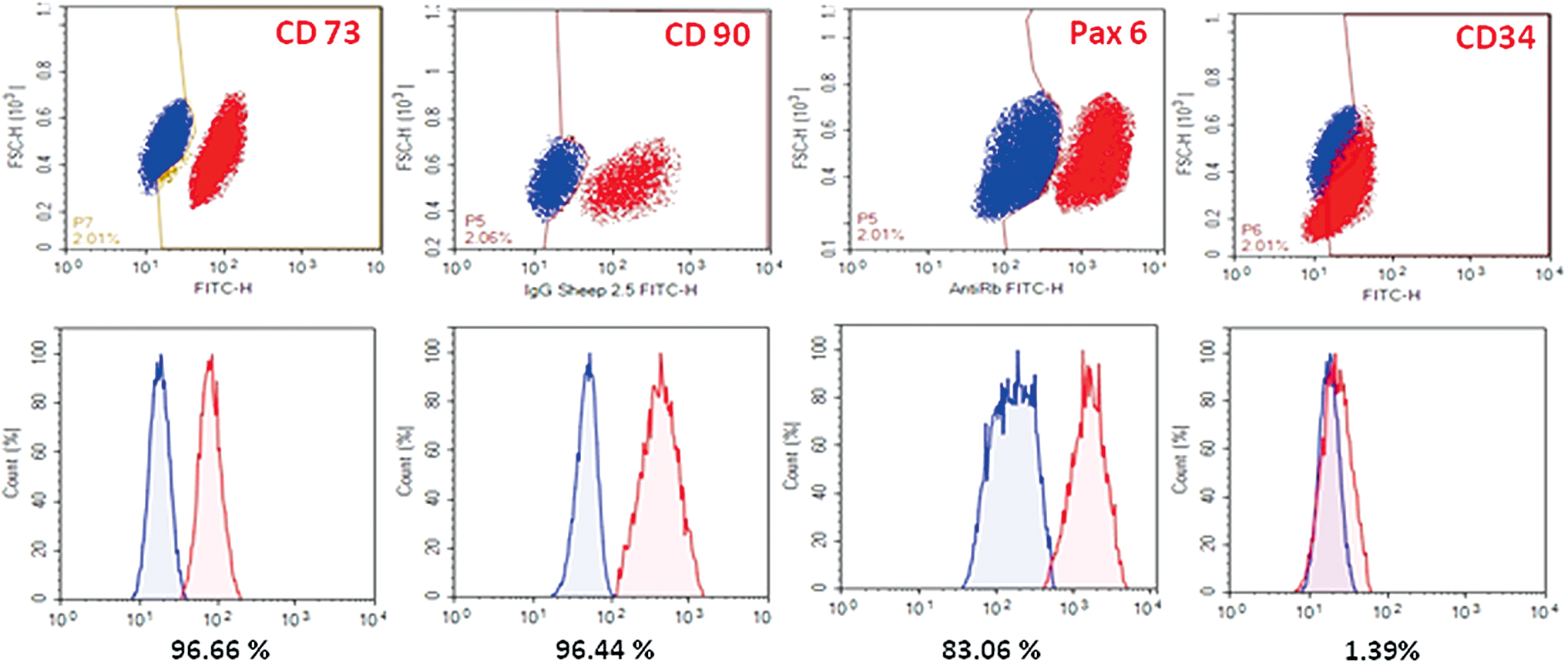
Quantification of MSC markers and Pax6 in limbal CSCs by flow cytometry. Cell quantification demonstrated in dot plots (FITC-H/FSC-H) and histograms of normalized cell counts (%) of positive gated limbal CSC (red) with an overlap allowance of 2% to the IgG isotype (blue). 96.66% of limbal CSC were CD73+, 96.44% CD90+, 83.06% Pax+ and 1.39% CD34+ (% in mean, n = 3). FITC-H, fluorescein isothiocyanate height; FSC-H, forward scatter height; MSC, mesenchymal stem/stromal cell. Color images are available online.
Western blot analysis confirmed the specificity and cross reactivity of the antibodies to the dog proteins for Pax6, N-cadherin, CD90, and CD73 (Supplementary Fig. S1).
Furthermore, CSCs could differentiate into bone, fat, and cartilage producing cells demonstrating their multipotency (Fig. 5). While there were no apparent differences in the high efficiency of CSC and MSC differentiation into adipocytes (Fig. 5A.vii, viii), the bone differentiation showed a moderate osteogenic response in both CSCs and adMSCs (Fig. 5A.i–vi) (compare to Supplementary Fig. S2). CSC differentiation into cartilage 3D (Fig. 5B.i–iv) and 2D (Fig. 5B.v, vi) required a higher concentration of TGF-β1 than that used for MSC differentiation (15 vs. 10 ng/mL).
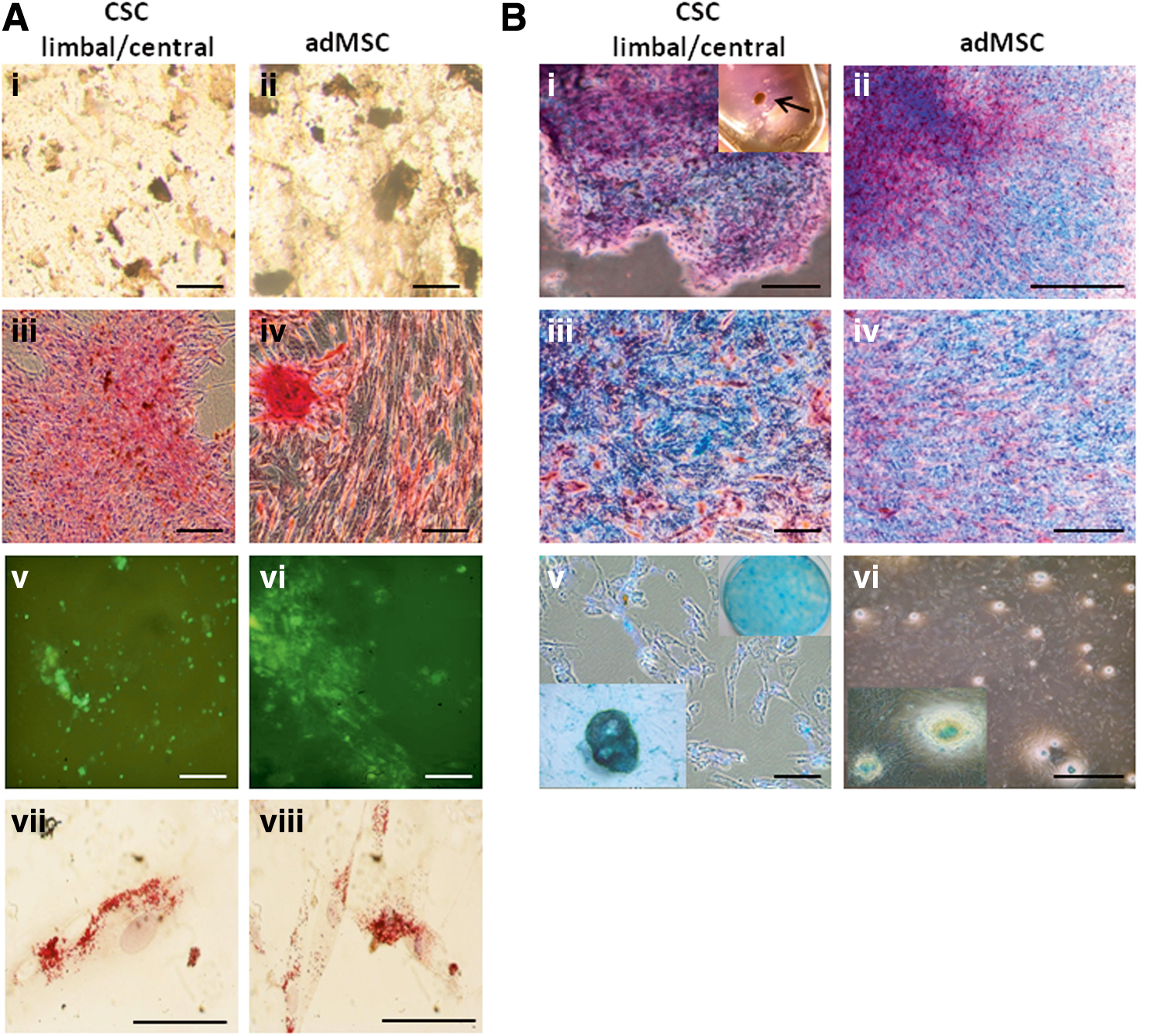
Trilineage differentiation of limbal/central CSC, adMSC
CSCs are immune privileged in vitro
Allogeneic limbal CSC cocultured with effector PBMCs did not modify the baseline level PBMC proliferation. In comparison, cocultures of effector PBMCs with allogeneic PBMCs showed a significant (P < 0.001) increase in proliferation compared to nonactivated PBMC (Fig. 6).
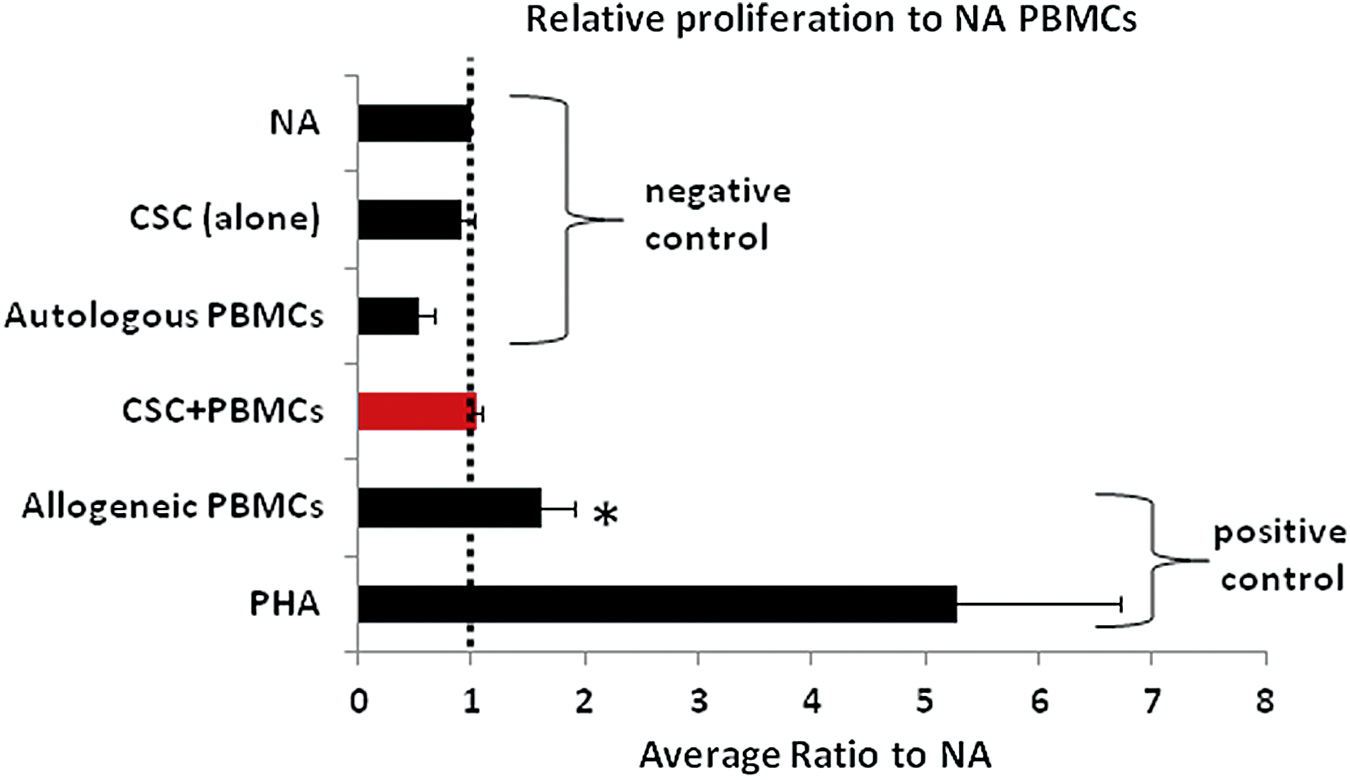
The PBMC coculture assay showed that allogeneic limbal CSC cocultured with PBMCs (red bar) did not modify the baseline level of NA PBMC proliferation. In comparison, cocultures of effector PBMCs with allogenic PBMCs showed a significant increase in proliferation compared to NA PBMC (P < 0.01). The values represent the mean average proliferation ratio (±SE) to NA PBMC highlighted with a dashed line; n = 3, *P < 0.05. NA, nonactivated; PBMCs, peripheral blood mononuclear cells; PHA, phytohemagglutinin; SE, standard error. Color images are available online.
CSC differentiate into KDCs and bFGF plays an essential role in the downregulation of α-SMA
Limbal- and central-derived CSCs were cultured under low glucose, serum-free conditions with substituted ascorbic acid to induce their differentiation into keratocytes (KDM). The small, polygonal morphology characteristic of CSCs changed to a stellate morphology typical of keratocytes within 7–10 days (Fig. 7A).
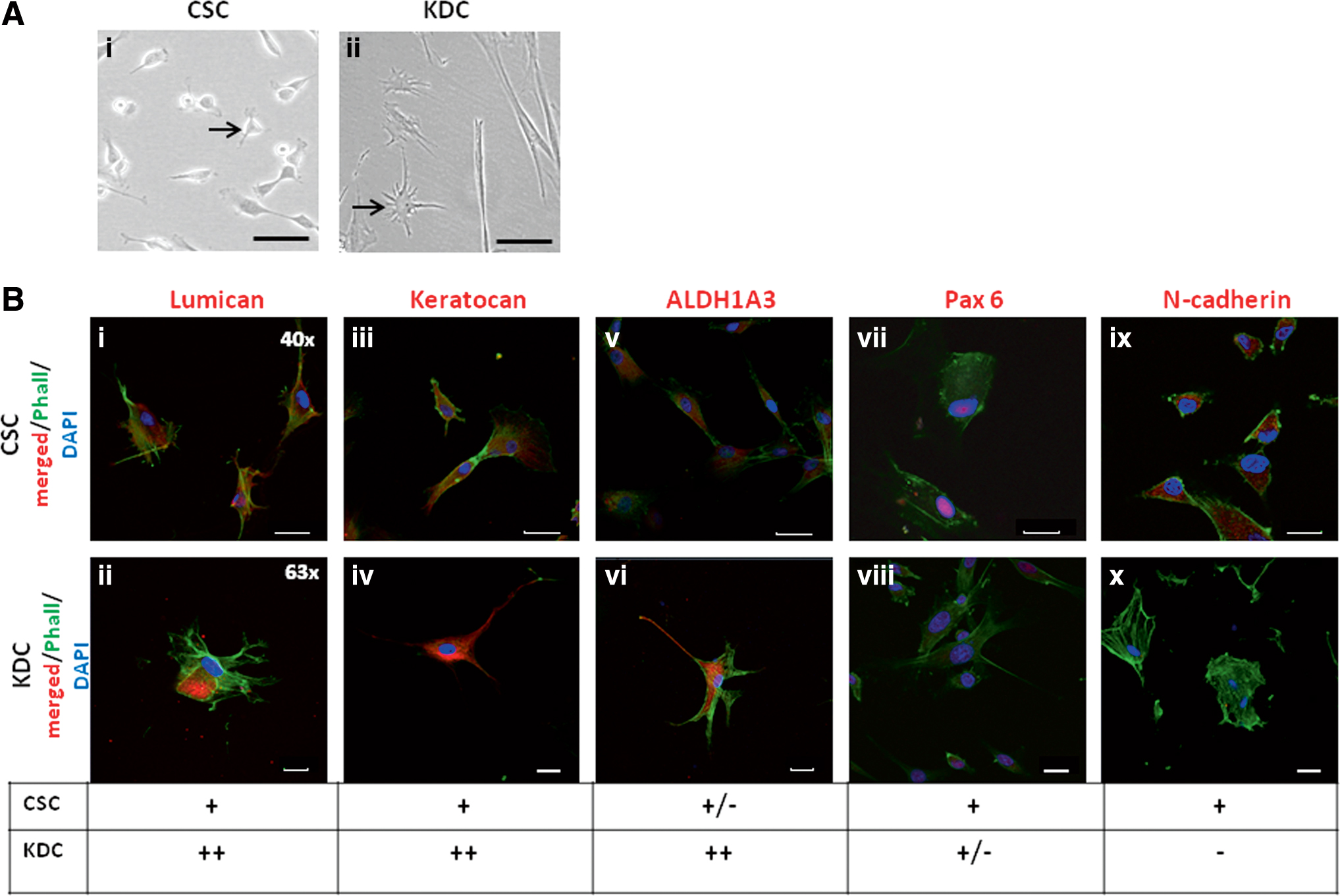
Limbal CSCs differentiation into KDCs in vitro.
A seeding number of 1 × 103 cells/cm2 differentiated in KDM containing human bFGF with an additional passage step maintained keratocyte marker expression without the expression of α-SMA over 21 days.
KDCs showed increased protein expression of Lumican, Keratocan, and ALDH1A3, whereas Pax6 expression decreased and N-cadherin expression became undetectable following differentiation (Fig. 7B). This was reflected in the fold change in gene expression of KDCs to undifferentiated CSCs where Keratocan was significantly upregulated (P < 0.05). Lumican (P = 0.27) and ALDH1A3 (P = 0.21) were also upregulated following differentiation, but did not reach significance. In contrast, the stem cell-associated gene Pax6 (P = 0.077) was downregulated and gene N-cadherin (P < 0.0001) was significantly downregulated (Fig. 8A).

Limbal CSCs differentiation into KDCs in vitro.
α-SMA is a myofibroblastic marker, which is minimally expressed at the gene level and not detected at the protein level in undifferentiated CSCs (Fig. 8B.i, C) or following keratocyte differentiation in the presence of bFGF (Fig. 8B.ii, C). However, in the absence of bFGF, α-SMA gene is significantly upregulated (P < 0.001) and protein expression is detected in the differentiated cells (Fig. 8B.iii, C).
Western blot analysis confirmed the specificity and cross reactivity of the antibodies to the dog proteins for Lumican, Keratocan, and ALDH1A3 (Supplementary Fig. S1).
Discussion
The corneal limbus in humans is located in the junction between the cornea and the sclera. In humans, the limbus is composed of limbal crypts and Palisades of Vogt, which serves as a stem cell niche, for LESCs, which are responsible for self-renewing and repair of damaged corneal epithelium [27]. The loss of LESCs results in Limbal Stem Cell Deficiency and leads to inflammation, scarring, neovascularization, and possible blindness [28]. The fate of LESCs within their niche is influenced by the stromal microenvironment. In humans, limbal stromal cells have MSC characteristics and are described as CSSCs [29,30] or limbal MSCs (L-MSCs) [9,10]. In humans, these stromal cells are hypothesized to assist in the maintenance of the LESC niche [31].
This is the first description of limbal- and central-derived CSCs with multipotent MSC characteristics in dogs, similar to those described in humans [7,8,12,20,32]. We also provide essential baseline data of the limbal anatomy in dogs.
A major anatomical difference to humans was the absence of limbal crypts and palisades of Vogt [33]. We can also confirm a slight invagination of the epithelium in the stroma as described by Patruno et al. [33]. Well-defined palisades of Vogt are present in the pig eye, but reportedly absent in rabbits and rodents [34,35].
Immunohistologically, a small population of CD90 expressing stromal cells in the limbal and central anterior corneal stroma was present. CD90 is a well-accepted MSC marker, however, it can also be expressed by fibroblasts [11]. To characterize the CD90+ cell population in more detail, double or even triple immunohistochemistry using CD90, α-SMA, and Vimentin should be included in future studies.
Canine MSCs are characterized in the veterinary field, including adipose-derived, bone marrow (BM)-derived MSC's, and other tissue sources as amniotic-derived, synovial-derived, periosteum [36 –42]. ISCT has defined a set of criteria that must be established to be characterized as a multipotent MSC [11]. Canine MSC studies fulfilled these criteria, however, positive and negative MSC marker profiles differ in various studies, some studies also demonstrated that cells were positive for the pluripotency-associated genes NANOG, OCT4, and SOX2 [41,43].
Canine MSCs are not a homogenous cell population. Hence, proliferation capacity has been reported to be lower in BM-MSCs in contrast to adMSCs and synovial MSCs [37,41]. Proliferation is also lower in MSCs isolated from older donors, “stemness” can decrease with passage frequency and the harvest site can have significant impact [43,44]. Breed-related differences have also been described. For example BM-MSCS of Howavart's had significantly less colony-forming units (CFU) than German Shepherd, Flat-coated Retriever, or Golden Retriever [45]. Therefore human MSC culture protocols were adjusted for canine MSCs [42,46,47].
This is the first report of canine MS-like cells of corneal origin and therefore the results are primarily compared to human corneal stromal MSC studies. The CSCs are isolated and cultured using different culture conditions to MSCs and therefore more detailed comparisons to canine MSCs in terms of culture conditions, cell proliferation, marker expression, and differentiation potential may be beneficial in future work.
Funderburgh et al. described a small population of limbal stromal cells expressing the transcription factor Pax6 (oculorhombin) in bovine corneas [7]. This could not be confirmed histologically in canine corneas [8].
Stromal cells from the canine limbal and central cornea were successfully isolated and cultured. Differential trypsinization of small polygonal cells, which reached a maximal confluency of 60%–70%, allowed the selective culture of CSC. A stem cell-like appearance could largely be maintained throughout several passages [15]. CFU-f assay of limbal and central CSCs as shown for human limbal CSSC was not performed, but should be included in future studies [9,10,48].
In contrast to Du et al., selective cell sorting to expand a CSSC “side population” was not performed [12]. This approach in human cells, which used the same low serum CSSC culture media as in the present study, resulted in a population with a replicative lifespan typical of stem cells (18 passages and a cumulative population doubling of 80 until senescence was reached) [12]. Human CSC populations cultured in 10% containing FBS without growth factor (EGF, PDGF), isolated without cell sorting produced more limited replication with 22.9 cell doublings between passages 2 and 6 (higher than in dogs). However, a PDT of 29.1 and 34.1 h similar to human MSC was observed only at early passages, similar to the present study [10].
Canine CSC underwent 12 and 11 population doublings for the limbal- and central-derived CSC, respectively, over 5 passages until senescence was reached. A PDT of ∼24–30 h as described in canine MSCs was only reached in the early log-phase (P1–P2) [36]. Guercio et al. also reports a limited life-span and senescence after passage 6 in canine adMSC (cultured in DMEM low glucose, 20% FBS) and revealed similar accumulated PD (11–12) in a similar time (27–28 days) to the present study of CSC. Bearden et al. showed that adipose and synovium canine MSCs proliferated first more rapidly with a rapid decline after passage 3, than marrow cMSCs, which we could very similarly observe in CSC. However, we hypothesis that the limited self-renewal of CSC population in dogs may suggest that they are proliferating progenitor cells rather than stem cells or might resemble a mixed population of cells [49]. Hence, the term canine CSC was used instead of CSSC as in humans.
The corneal stem cell media could also influence the degree of self renewal and might require optimization to the canine species in future studies. CSSC media contains only low serum levels of 2% FBS and not 10% or higher as in most canine MSC culture conditions. This was performed as higher serum concentrations have been shown to induce a myofibroblastic cell fate in rabbit keratocytes [50]. Serum supplementation (10% v/v) led to human keratocyte differentiation into fibroblasts with loss of keratocan expression in human CSSC [12 –14]. Whether this holds true for canine keratocytes and canine CSC is unknown.
The CSSC media also contains the growth factors EGF and PDGF. It is described that factors such as FGF, TGF- β, PDGF, insulin-like growth factor 1 (IGF-1), and EGF regulate keratocyte differentiation, migration, and expression and are known to be essential growth factors in corneal in vitro studies [51,52]. A canine study of BM-MSCs reported that cells expanded best in a-MEM supplemented with bFGF [45] and future studies to optimize canine CSC media to maximize self-renewal and differentiation potential are required.
No significant differences in culture kinetics were observed between limbal- and central-derived CSCs, although the central CSC showed a trend toward lower frequency of passage, lower cell numbers and earlier senescence. There was a high degree of heterogeneity in cell expansion, which might be explained by the wide age range of donor tissue (0.2–12 years) that was used in this study, comparative studies of the influence of age on human CSSCs, which are isolated and cultured similar to canine CSC in the present study, are lacking. However, donor age can negatively impact human and canine MSC cell expansion and differentiation [44,53].
Effects of age in skeletally immature (mean age: 4.9 ± 1.9 months) and mature (mean age: 89.5 ± 20.9 months) dogs on canine BM-MSCs were studied by Volk et al.. BM cells were isolated from long bones (humeri, femurs, and tibias) and cultured in high-glucose DMEM substituted with 10% FBS, showed a significant negative effect of age on both MSC frequency and the ability to differentiate along the osteogenic lineage [44].
Guercio et al. compared the age (young dogs: 1–4 years; adult dogs: 8–14 years) and the harvest site (subcutaneous vs. visceral) of adMSCs cultured in low-glucose DMEM and 20% FBS. Population doubling values at passages >2 were significantly higher for MSCs derived from subcutaneous fat and in younger dogs. CFU-f assay in low glucose DMEM and 5% FBS did not differ according to age and harvest location site, but declined after passage >2.
The influence of donor age on cell expansion and differentiation of canine CSCs warrants further investigation in future studies.
We were able to differentiate both limbal- and central-derived CSC and adMSC into chondrogenic, adipogenic, and osteogenic cells in vitro. However, it should be noted that canine chondrogenesis in MSC has not been robustly demonstrated in the literature and difficulties in establishing a protocol are described [37,42]. To successfully drive chondrogenic differentiation of the CSCs, we had to use an increased concentration of TGF-β1 compared to the MSCs. However, further optimization of the differentiation protocol could be performed [41] or a standardized commercial available differentiation media could be tested in future studies [43].
The induction of the osteoblastic lineage was demonstrated using a combination of van Kossa (mineralization–phosphate) and Alizarin S red (calcium) and hydroxyapatite staining but differentiation was not as robust when compared to SAOS-2 cells (Supplementary Fig. S2). Passage number, pluripotency gene expression [43], and donor age have all been shown to affect the differentiation capacity of canine MSCs and were not tightly controlled in the present study (cells were at passage 2–5 and from donors aged 0.2–12 years). Future work could also optimize the osteogenic differentiation protocol, for example, through the addition of BMP-2 or IGF-1 [41,47,54].
The immunophenotyping of limbal- and central-derived CSC showed remarkable similarity with the surface antigen profile of adMSC for CD90, CD73, CD105, N-cadherin, and CD34. In contrast to most human studies characterizing limbal CSCs, the present study used a limited number of protein markers; however, this reflects the nature of establishing protocols for the use of markers on canine tissue and cells. Most of the markers are not established in this species, and therefore, western blot analysis had to confirm the specific binding.
The expression of CD34 in human CSSCs and keratocytes is controversial in human corneal research. CD34 is a hematopoietic stem cell surface marker, which is defined to be absent (<2%) in MSCs [11]. This was also confirmed for human limbal and central CSSCs [8,10].
CD34 has been described as a characteristic marker of quiescent keratocytes in the human cornea [55]. Another study characterizing L-MSCs showed a drop of CD34 marker expression with increasing cell proliferation and a shift to more progenitor cell type. This seems to be influenced significantly by the composition of culture media [9,56,57]. In our study, we found CD34 protein expression in <2% of canine CSCs. Canine adMSC in the present study also did not express CD34, which is similar to other reports for canine MSCs [36 –38].
In contrast to CSCs, adMSC did not express Pax6, which was not expected given their origin. However, adMSC did express α-SMA, which was not detected at the protein level in undifferentiated CSCs. α-SMA expression has been reported in human, rat, and murine MSCs previously [58 –60].
α-SMA, the earliest known protein expressed in differentiation of the smooth muscle cells during development and is also transiently expressed by a variety of mesodermal cells during development, tissue repair, and neoplastic growth [60]. Tissue repair and tumor microenvironment can convert MSCs into contractile MFs that form α-SMA-containing stress fibers [61,62]. MF activation is part of the wound healing response, but persistent MFs contribute to fibrosis by excessively producing and contracting collagenous extracellular matrix into stiff scar tissue [62].
Human CSCs have been shown to not only fail to induce the proliferation of allogeneic PBMCs, but actually suppress the proliferation of activated PBMCs in vitro [8]. We have demonstrated that canine CSCs also appear to be immune privileged in vitro as they fail to induce the proliferation of PBMCs isolated from different donors. We carried out these assays on three different lines of CSCs each using PBMCs from two different donors (with PBMCs isolated from both lymph nodes and commercial suppliers) and all replicates produced very consistent results. However, the cells were not typed for dog leukocyte antigens (DLA) and, therefore, we cannot rule out the possibility that the cells were not true-mismatches. Further work is also required to determine if the CSCs are immune suppressive.
Canine CSCs (limbal and central) were successfully differentiated into cells with stellate keratocyte typical morphology that expressed keratocyte markers Keratocan, Lumican, and ALDH1A3 [15,63,64]. ALDH1A3 is expressed in the murine cornea and is also required for corneal maintenance [65,66]. Similar to human studies, we could demonstrate that the gene expression levels of stem cell markers were downregulated and keratocyte markers were upregulated after 21 days of differentiation [12,67].
Canine CSCs continued to proliferate during the keratocyte differentiation process, which might be driven by bFGF [68]. Therefore, KDM without bFGF was tested, which led to stable cell numbers during differentiation. Although the absence of bFGF during the differentiation had no influence on keratocyte marker expression, it did lead to a significant increase in α-SMA gene expression and the induction of detectable levels of protein. bFGF has been shown to downregulate α-SMA expression in adMSCs previously [69]. In the injured cornea, keratocytes differentiate into MFs in response to TGF-β1 [70]. To date, the reverse process turning MFs into keratocytes has not been described [71]. From a translational medicine perspective, using α-SMA expressing cells in vivo should be further investigated carefully.
In conclusion, the limbus of dogs lacks limbal crypts as in humans but has an undulating basement membrane with CD90+ stromal cells in close proximity to the limbal epithelium. For the first time, we characterize a cell population in the canine corneal stroma (limbal and central) that contains MSC-like cells similar to CSSCs in humans with keratocyte differentiation potential and possible immune privilege properties. This novel finding of CSCs with multipotent MSC properties will provide a baseline for researchers working with canine corneal disease models and for cell-based therapy in veterinary ophthalmology.
Footnotes
Acknowledgment
The authors thank Dr Jan van Dijk (Animal Health Trust) for support with the statistical analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank Petplan Charitable Trust (S16-204-397), Bertie's Mission, and European College of Veterinary Ophthalmologists (research grant 2017) for funding the work.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.