
Abstract
In mammals, spermatogenesis is closely related to intercellular interactions of germ cells and surrounding Sertoli cells, that is, blood–testis barrier and gap junction, which are subjected to hormone signals in testicular seminiferous tubules. Androgen signal plays pivotal role in regulating spermatogenesis, but the underlying mechanism is largely unknown. Our recent study demonstrated a novel regulatory pattern of androgen in regulation of spermatogonia differentiation, in which androgen indirectly regulates the expression of ITGB1 on Sertoli Cells through intermediate molecule Wilms tumor-1 (Wt-1) during spermatogenesis. In this study, we identified that Connexin 43 (Cx43), the key component for gap junction distributed between spermatogonia and Sertoli cells, was also regulated by androgen signal. Chromatin immunoprecipitation quantitative polymerase chain reaction (ChIP-qPCR) demonstrated that WT1 occupied Cx43 promoter in Sertoli cells, suggesting WT1 as an intermediate molecule in regulation of Cx43. Finally, we revealed a regulatory pattern of Cx43 by androgen in Sertoli cells, and the Sertoli cells in germ cell depleted microenvironment were sensitive to androgen signal, which enhances the understanding of the mechanism of spermatogenesis.
Introduction
Spermatogonial stem cells (SSCs) determine the fertility of male animals, and the capacities of self-renewal and differentiation of SSCs are regulated by many factors. The microenvironment of SSCs (also nominated as niche) is a special structure formed by somatic cells, including Sertoli cells, and is able to regulate SSCs' fates through intercellular interactions between SSCs and Sertoli cells. In spermatogenesis, SSCs form different subpopulations (As, Apr, Aal, A1…A4, AIn, B, etc) through differentiation. These subpopulations, including As, Apr, and Aal spermatogonia, are usually referred to as undifferentiated population and are nominated as spermatogonial progenitor cells (SPCs) [1]. The size of SPCs (usually represented by PLZF+ population) determines the capacity of spermatogenesis [2], and our recent study noticed that this population is stable during testis development [3]. Moreover, we observed that Sertoli cells are essential to regulate SPC differentiation stimulated by androgen [3], indicating that androgen indirectly regulates neighboring SPCs of Sertoli cells through intercellular interaction. Among many types of intercellular interactions, gap junctions between two Sertoli cells are identified as an important component in blood–testis barrier (BTB), which separates lumen and basal space and efficiently blocks molecule diffusion between those two spaces to provide undifferentiated spermatogonia with an independent microenvironment [4]. In the niche, the functional mechanism of gap junction is interesting but not fully understood yet.
Connexin is a type of transmembrane protein functional as a major component of gap junctions in vertebrates. In rodent, Connexin 26, 30.2, 31, 31.1, 32, 33, 36, 37, 40, 43, 45, 46, 50, and 57 have been identified, and in gap junction of mouse Sertoli cells the mainly subtype is connexin 43 [5]. Cx43 is involved in the development of spermatogonia and spermatogenesis [6] and is closely related to male fertility [5]. Recent studies reported that the toxic effect of chemotherapeutic drugs is able to spread to neighbor cells through gap junction [7,8]. For male cancer patients subjected to chemotherapeutic treatment, impairment of fertility probably results from the transfer of toxic chemicals from Sertoli cells to germ cells through gap junction. These evidences suggest that Cx43 is a key component in the testicular microenvironment; thus, the underlying regulatory mechanism on Cx43 is valuable to dig in.
Our recent study screened the potential downstream targets of androgen receptor (AR) and WT1 in mouse Sertoli cells and investigated the role of androgen signal in spermatogenesis. Besides the known role as a tumor suppressor, Wilms tumor-1 (Wt-1) gene is an essential gene for embryo development, and loss of function assay revealed that mutation of Wt-1 in germinal ridge causes developmental obstacle of germ line [9]. Moreover, Wt-1 functions as an activator or a suppressor in gene transcription regulation [10]. Data from in vitro experiments reveal that WT1 coactivates the transcription of Sry with GATA4 [11]. As the key gene of testis development, WT1 has also been identified as a repressor of AR [12]. However, we demonstrated that AR could regulate Wt-1 expression through Gata2 [3], indicating a complicated regulatory pattern of androgen signal in testis. These observations suggest that WT1 involves in multiple signal pathways correlated to the functions of Sertoli cells. A study reported that androgen regulated permeability of the BTB by regulating expression of Claudin-3 in Sertoli cells, indicating the important role of androgen in controlling the process of spermatogenesis [13]. Consistently, our study demonstrated that AR regulated WT1 through Gata2 in Sertoli cells, and WT1 was able to bind to many target genes related to cellular interaction, that is, β1-integrin [3]. Therefore, we reanalyzed the raw data from two ChIP-seq assays (
In this study we investigated the expression of Cx43 in mouse testis, primary SPCs, and long-term cultured SPCs and detected the Cx43 signal in the boundary of spermatogonia and Sertoli cells in the seminiferous tubules and in primary SPCs, but not in the long-term cultured SPCs. Subsequently, the link of AR and Cx43 was investigated in Sertoli cells, and a potential regulatory pattern of gap junction by AR signal was proposed.
Materials and Methods
Animals
The CD-1 mice were supplied by Comparative Medicine Centre of Yangzhou University for experiments in this study, and all of the procedures for animal experiments were approved by the Ethics Committee at Nanjing Agricultural University.
Sorting and culture of mouse SPCs and Sertoli cells
SPC sorting was preformed according to previous protocol [14]. Briefly, testes harvested from 5-day male mice were cut into small particles and then were digested with collagenase IV and trypsin to obtain single cell suspension. After centrifugation, cell pellet was resuspended with culture medium and filtered with 70-μm cell filter and, subsequently, sorted using mouse Thy-1.2 antibody coated magnetic beads (Cat. no. 551518; BD). The Thy1.2+ fraction was cultured on mitotically inactivated mouse embryonic fibroblast (MEF) feeder layer at 37°C with 5% CO2. Methods for making culture medium for SPCs and preparation of MEF feeder cells were also followed with previous protocols [14].
The primer sequences for RT-PCR were listed below:
Gfra1 F: ACTCCTGGATTTGCTGATGTCGG, Gfra1 R: CGCTGCGGCACTCATCCTT;
Plzf F: CACACTCAAGAGCCACAAGC, Plzf R: ATCATGGCCGAGTAGTCTCG;
Ar F: GGACCATGTTTTACCCATCG, Ar R: TCGTTTCTGCTGGCACATAG;
Cx43 F: TGAAAGAGAGGTGCCCAGACA, Cx43 R: CGTGAGAGATGGGGAAGGACT;
Wt-1 F: GTGAAACCATTCCAGTGTAAAAC, Wt-1 R: GCCACCGACAGCTGAAGGGC;
Gapdh F: CCTGGAGAAACCTGCCAAGTATG, Gapdh R: GAGTGGGAGTTGCTGTTGAAGTC
The sorting procedure of Sertoli cells was identical to previous study [3].
Chromatin immunoprecipitation quantitative polymerase chain reaction
The protocol for chromatin immunoprecipitation quantitative polymerase chain reaction (ChIP-qPCR) was identical to previous study [3]. Briefly, Sertoli cell samples were cross-linked with formaldehyde and then were inactivated by glycine. Subsequently, the sonicated DNA fragments of 100–300 bp were precleared and then immunoprecipitated with Protein A + G Magnetic beads coupled with Anti-WT1 antibody (ab89901; Abcam). After reverse cross-linking, immunoprecipitated DNAs and input DNAs were end-repaired and ligated to adapters using NEBNext Ultra End-Repair/dA-Tailing Module (E7442, NEB) and NEBNext Ultra Ligation Module (E7445, NEB). Finally, the DNA samples were used for real-time PCR.
Sequences of primers for ChIP-qPCR were listed below:
Cx43 F: GCTAGAGAGTGCATTGGGGA
Cx43 R: AGTTCCTGGGGCTGCGAG
ITGB1 F: AGCTACATTTCTGAGCAATTATGGA
ITGB1 R: CTGCTCTCCCAGGACTCAAC
Control F: TTAGGTGGCCTCAGATCCTC
Control R: CCTGCCTCTCTTTTGGACAG
Immunohistochemistry and immunofluorescence
The protocols for immunohistochemistry (IHC) and immunofluorescence (IF) staining were identical to previous study [14]. The information for antibodies used in this study is as follows: Antibody: E-cadherin (ab76055; Abcam), promyelocytic leukemia zinc-finger (PLZF) (sc-2831 and sc-22839; Santa Cruz), Cx43 (A0239; ABclonal), WT1 (ab89901; Abcam), AR (ab133273; Abcam), β-tubulin (abs830032; absin), and biotin labeled goat anti-mouse immunoglobulin G (IgG) (BA-9200; Vector) or goat anti-rabbit IgG (BA-1000; Vector).
siRNA and plasmid transfection
The siRNA oligos were synthesized by Shanghai GenePharma Company. AR expression plasmid was a gift from Prof. Zijie Sun (School of Medicine, Stanford University). Wt1 cDNA expression plasmid was purchased from GeneCopoeia (EX-Mm24822-02). Plasmids or validated siRNA oligos were transfected using Lipofectamine 3000 (Life Technology).
Cx43 siRNA sequence: CAAUUCCUCCUGCCGCAAUTT [15],
Wt1 siRNA sequence: GGCGAUUGUUAAAGCUCAUUU [16],
control siRNA sequence: UUCUCCGAACGUGUCACGUTT [3].
Western blot
For western blotting, tissue and cell lysates were separated in sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel and transferred to nitrocellulose membrane, which was subsequently blocked with 5% skim milk at 4°C overnight and then incubated with primary antibodies at room temperature for 1 h. Then the membranes were incubated with horseradish peroxidase-conjugated goat anti rabbit or mouse IgG (sc-2004, sc-2005, 1:3,000; Santa Cruz) and enhanced chemiluminescence and, finally, exposed the film (see antibody information in Supplementary Table S1 through this
Busulfan/bicalutamide intraperitoneal injection
The protocol for busulfan/bicalutamide intraperitoneal injection was identical to previous study [3]. Briefly, 24 six-week-old male CD1 mice were intraperitoneally injected with busulfan at dose of 40 mg/kg and were injected with 40 mg/kg busulfan again after 10 days. Twenty days post busulfan injection, these mice were randomly divided into two groups and were intraperitoneally injected with 20 mg/kg bicalutamide or dimethyl sulfoxide (DMSO) once every other day for four times, respectively.
Statistical analysis
All of the sections or immunofluorescent visual fields were selected randomly for cell counting, and data were subsequently analyzed by Excel and were presented as mean ± standard deviation. Statistical significance was determined by t-test.
Results
Expression profile of Cx43 at different development stages of mouse testis
To reveal the expression profile of Cx43 in mouse testis of each developmental stage, the expression of Cx43 in testes from 5-day (neonate), 10-day (puberty), 20-day (juvenile), and 42-day (adult) mice was detected using IHC. Cx43 was detected at the boundaries of Sertoli cells and spermatogonia in 5-day testes (Fig. 1A). In 10-day (Fig. 1B), 20-day (Fig. 1C), and 42-day testes (Fig. 1D), Cx43 signals were detected not only on boundaries of Sertoli cells and spermatogonia but also on the boundaries of Sertoli cells and spermatocytes. Notably, Cx43 signals were detected on the surface of SSCs in 42-day testis (Fig. 1D, inset). To further confirm the expression of Cx43, PLZF/Cx43 dual IF assay was adopted to detect the expression of PLZF and Cx43 on SPCs. Results displayed that some of PLZF+ spermatogonia localized on the base membranes were also Cx43 positive (Fig. 1E–H), indicating that the undifferentiated spermatogonia in the basal membrane express Cx43 and may form gap junctions with neighboring cells.
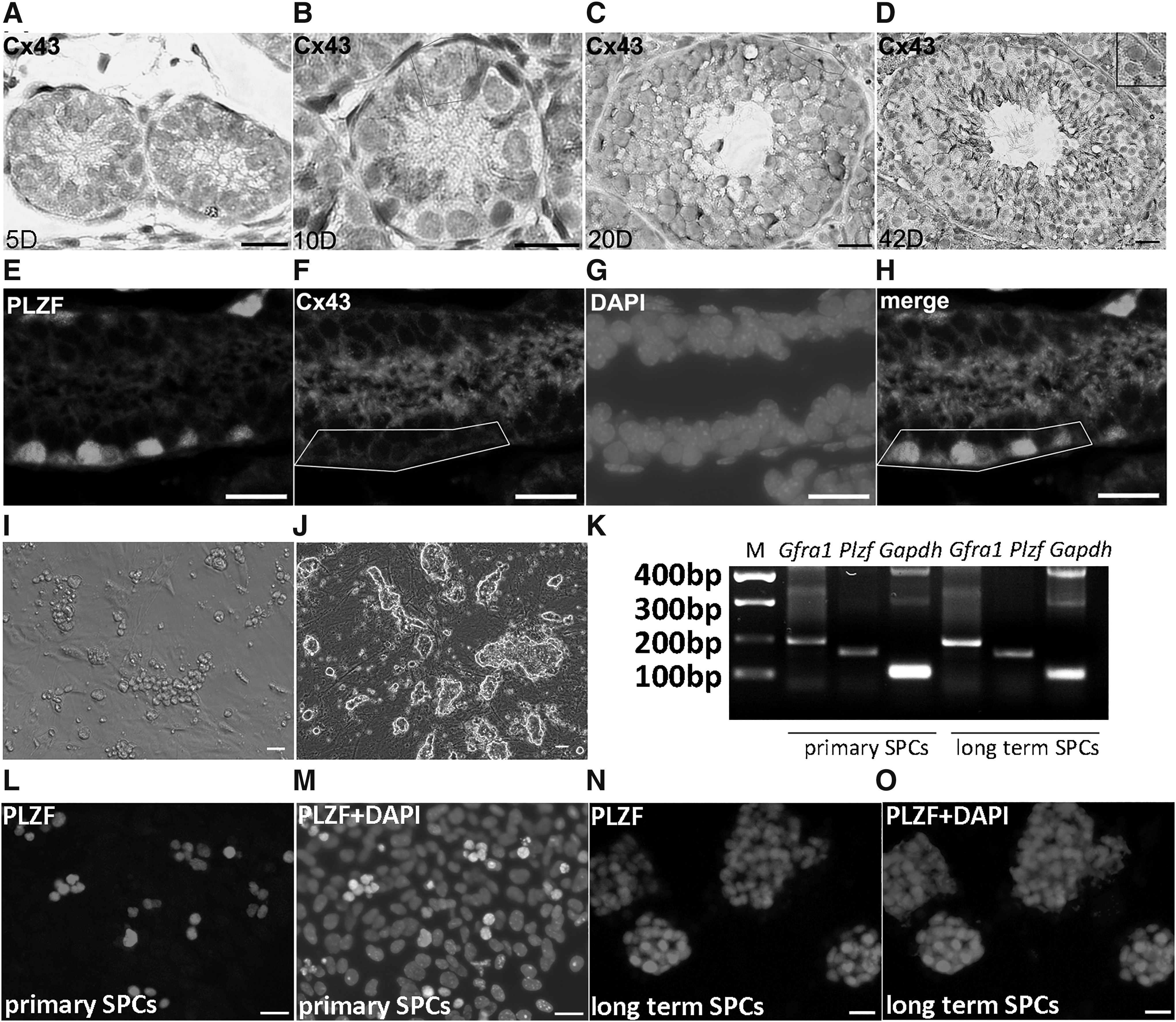
Expression profile of Cx43 in mouse testis. Expression of Cx43 was determined in testes of 5 days
Primary SPCs and Sertoli cells express Cx43
Aforementioned IHC assays detected Cx43 signal between SPCs and Sertoli cells in the niche and then we ask whether both of these two types of cells independently express Cx43. To answer this question, primary SPCs (Fig. 1I) and long-term cultured SPCs (more than 20 passages) (Fig. 1J) were used for analysis. Both of them expressed SPC marker Gfra1 and Plzf (Fig. 1K–O). Subsequently, E-cadherin/Cx43 dual IF assay was used to verify whether SPCs express Cx43 independently. Cx43 signal was detected on E-cadherin+ spermatogonia (Fig. 2A). Interestingly, long-term cultured SPCs were E-cadherin positive but Cx43 negative (Fig. 2B), implying that the expression profile of SPCs was changed during long term culture, and these cells became dispensable for gap junction with neighboring cells. Moreover, purified Sertoli cells (WT1+ ratio was 91.3 ±2.12%) (Fig. 2C, D) were used for Cx43 IF staining, and Cx43 signal was detected on Sertoli cells as well (Fig. 2E), indicating that probably gap junction is a potential interaction pattern for SPC-Sertoli cell communication.
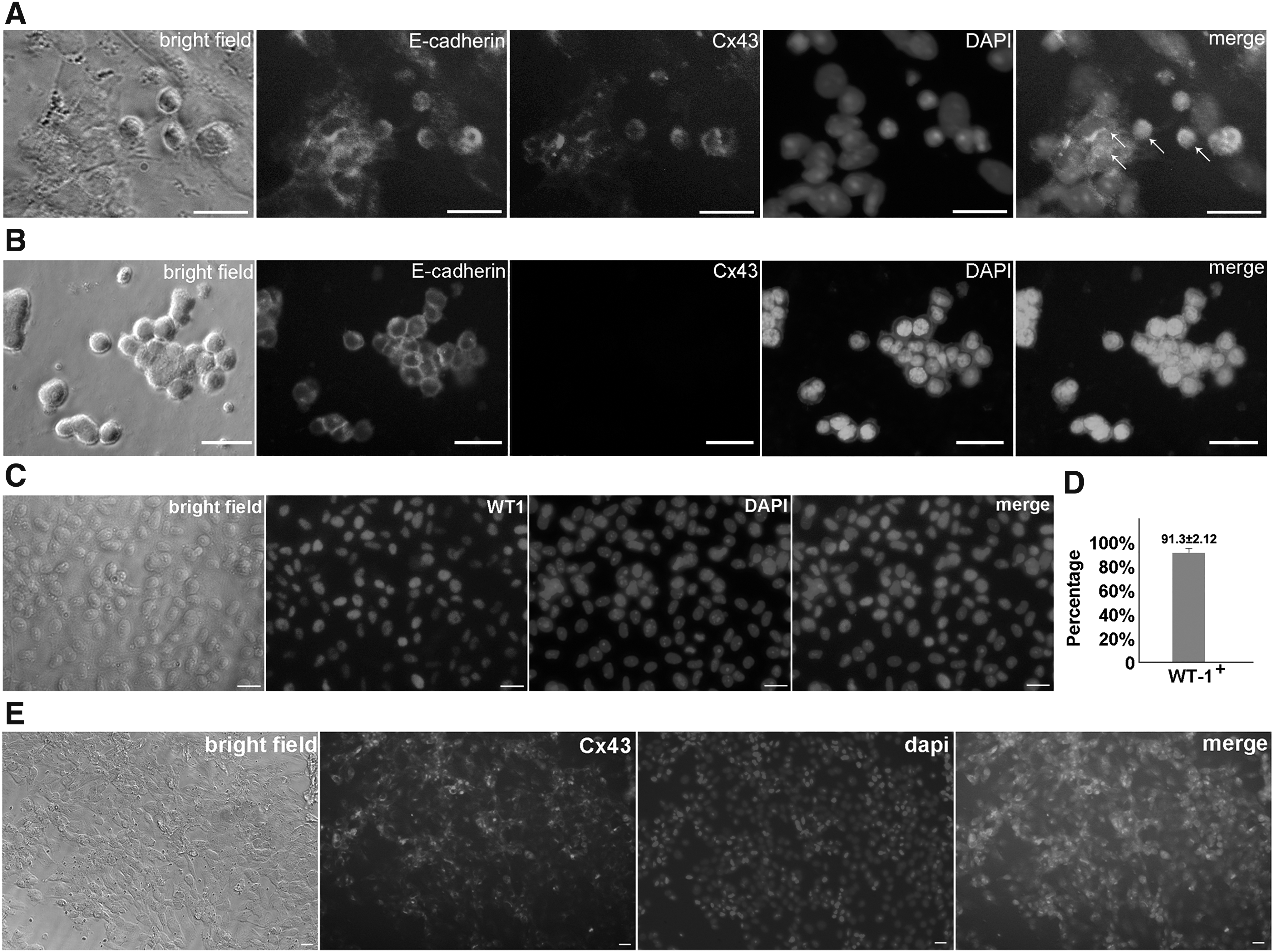
Detection of the expression of Cx43 in SSCs and Sertoli cells. Dual immunofluorescent staining of E-cadherin and Cx43 in SPCs derived from 5-day testes
Link of androgen and Cx43 in Sertoli cells
Our previous study revealed that androgen regulates many potential target genes, including Wt-1, which is closely related to spermatogenesis [3]. In this study, we ask whether androgen regulates expression of Cx43 in Sertoli cells. Purified Sertoli cells were treated with dihydrotestosterone (DHT) and/or bicalutamide (an efficient AR antagonist), respectively. DHT stimulated the expression of Cx43, whereas bicalutamide caused an opposite effect (Fig. 3A). To further confirm the regulatory effect of androgen, expression of AR was disturbed or enhanced with transfected siRNA or AR overexpression plasmid in Sertoli cells, respectively. Consistent results were observed. WT1 was negatively regulated by AR, whereas Cx43 exhibited a positive correlation to AR signal (Fig. 3B, C). These observations primarily confirmed our proposal that androgen signal regulated the expression of gap junction pivotal component Cx43 in Sertoli cells.
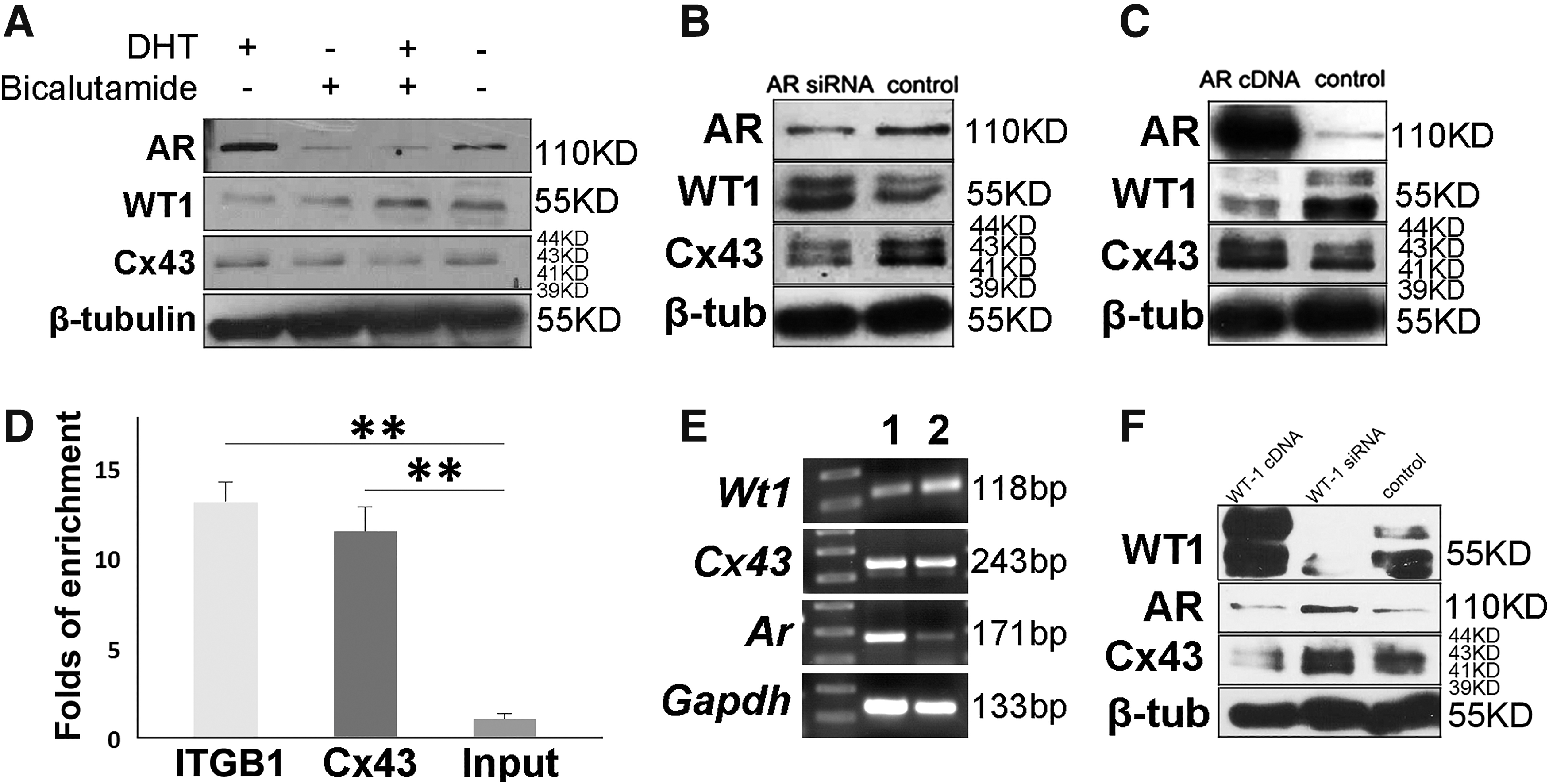
The regulatory effect of androgen on the expression of Cx43. Androgen treatment or deprivation in Sertoli cells enhanced or suppressed expression levels of Cx43
Cx43 gene is a direct target of WT1 in mouse Sertoli cells
Based on the positive correlation of androgen signal and Cx43 from above data, we wonder whether AR or WT1 could directly bind to the promoter region of Cx43. After screening ChIP-seq data in previous study [3], we noticed that WT1 has a direct binding site on the promoter region of Cx43 (Supplementary Table S2, which is available at
Androgen deprivation influenced the expression of Cx43 in SSC niche
Busulfan is a cell cycle nonspecific alkylation agent with germ cell toxicity, but has limited impact on surrounding somatic cells. After treatment with busulfan, transplanted SSCs can still survive, localize, and restore spermatogenesis in the remaining microenvironment of busulfan recipient mice testes [17]. Thus, we intraperitoneally injected busulfan to 6-week-old male mice, which already form mature and stable niche, to establish an in vivo model to study the impact of androgen on Cx43 in SSC niche. Compared with normal testicular niche, which is occupied by spermatogonia (Fig. 4A), busulfan recipient exhibited the intact structures of Sertoli cells and Leydig cells, but the empty cavities in the seminiferous tubules (Fig. 4C). Interestingly, IHC staining demonstrated that the expression of Cx43 remained on the surfaces of Sertoli cells in the empty niches, even if spermatogonia were absent (Fig. 4D), since Cx43 should be distributed between germ cells and Sertoli cells in wild type control (Fig. 4B). It's not clear why Sertoli cells keep on expressing Cx43 in the absence of spermatogonia, but this explains why the transplanted SSCs can survive in new microenvironment, since the empty niche keeps on being hospitable to SSCs. Moreover, these results suggested the successfully established in vivo model for the study of Cx43 in the niche. Subsequently, 6-week-old male mice were intraperitoneally injected with busulfan to eliminate germ cells and then were intraperitoneally injected with bicalutamide or vehicle (DMSO) for four times 30 days post busulfan injection. The sizes of both busulfan recipient testes and busulfan/bicalutamide recipient testes were significantly reduced than control group (Fig. 4E). Western blot analysis demonstrated that busulfan injection upregulated the expression levels of AR and Cx43, since deletion of germ cells led to an increased proportion of Sertoli cells in testes (Fig. 4F, G). Compared with busulfan recipient testes, busulfan/bicalutamide recipient testes exhibited relatively low expression levels of AR and Cx43 (Fig. 4F, G), implying that deprivation of androgen in germ cell-deleted niche suppressed Cx43 expression, which is consistent with the above results. Finally, to further verify this regulatory effect of WT1 on Cx43 expression, Sertoli cells treated with bicalutamide for androgen deprivation were transfected with control or Wt-1 siRNA, respectively. Expression of Cx43 was upregulated by bicalutamide, but was negatively related to WT1 (Fig. 4H), which was consistent with aforementioned observations. Combined with previous study which revealed that androgen regulates expression of WT1 through GATA2, here we proposed a new regulatory pattern on Cx43 expression in Sertoli cells: androgen signal regulates WT1 expression through GATA2, and WT1 subsequently binds to Cx43 promoter as a transcription suppressor to regulate gap junction (Fig. 4I).
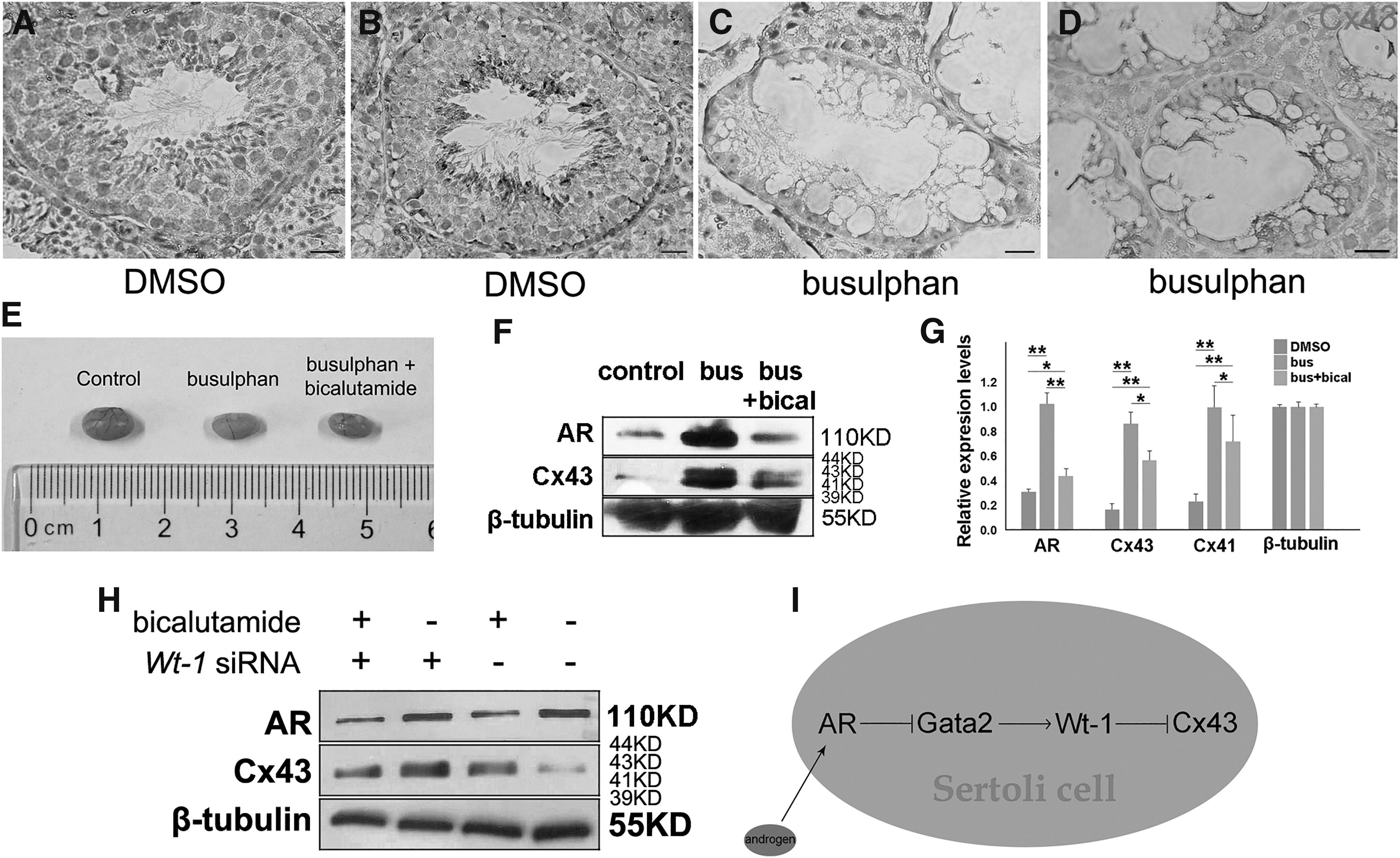
Androgen regulates expression of Cx43 in SSC niche. Compared to wild-type control group
Discussion
The role of Cx43 and gap junction in Sertoli cells has been demonstrated in previous studies. Our results revealed that SSCs from neonatal testis endogenously express Cx43, suggesting a potential function of gap junction between SSCs and Sertoli cells. However, it is not clear whether spermatogonia in other differentiation stages express Cx43. Following experiments showed that Cx43 signals were mainly detected in spermatogonia and Sertoli cells at the basal membrane, in primary SPCs, but not in long-term cultured SPCs, probably because SPCs changed their expression profiles and intercellular interaction pattern after leaving microenvironment. This observation highlights one of the differences between primary SPCs and SPC cell line. Based on our experience of SPC culture, primary SPCs are not easy to be maintained in vitro, and the average doubling time of primary SPCs usually takes about 6–8 days in vitro, whereas that of long-term SPC cell lines is only 2–3 days under identical culture condition, which is consistent with reported study [18]. This implies that the dependence of intercellular interaction, that is, gap junction, is one of potential reasons to hamper the maintenance and proliferation of primary SPCs, and cells which became independent of intercellular interaction may survive under in vitro culture and finally grow rapidly.
Studies reported that the dynamic expression pattern of Cx43 is determined by spermatogenic cycle [5], and the life expectancy of Cx43 is as short as several hours [19]. However, in this study testes were harvested about 40 days post busulfan/bicalutamide injection, but Cx43 expression was still detected, and expression level of Cx43 in empty niche could be inhibited by bicalutamide. It indicates that the niche is still “alive” and keeps on being hospitable and compatible to SSCs, even if SSCs are absent for a long term. Moreover, the expression of Cx43 on Sertoli cells seems to be independent to spermatogonia, which is consistent to the study which revealed that busulfan injection does limited harm to niches: only expression of Sox9 was declined, whereas expression of WT1, Claudin-11, and platelet derived growth factor receptor alpha (PDGFRA) was unaffected [17]. This confirms that the remaining niches after germ cell depletion by busulfan are suitable for SSC transplantation model.
It's of importance to understand the regulatory pattern of SSCs in the niche for reproductive and clinic research, since testis is a vulnerable target of environmental toxicants [20]. Pharmacologically germ cell depleted mice provide a valuable model for studying the regulatory mechanism in the niche. Removal of germ cells with busulfan injection showed no impact on the expression of Claudin-11, the key component of tight junction between Sertoli cells and closely associating with germ cell migration in the niche [17]. On the contrary, Claudin-3, another claudin protein expressed restricted to newly formed tight junctions, is responsive to androgen signal [13], which is similar with regulatory pattern of androgen signal on the gap junction key component Cx43 revealed in this study. This evidence suggests that cell junction associated members are more likely to be subjected to androgen signal, rather than signal from germ cells. However, considering that Claudin-3 in newly formed tight junctions will be replaced by Claudin-11 during germ cell migration [21], the impact of germ cell interaction on Sertoli cells is interesting and complicated, which needs to be further investigated in further study.
The regulatory mechanism of androgen in testicular niche is complicated. Research revealed the link of androgen signal and BTB [13], and our data from previous studies and this study demonstrated that androgen promotes SPC differentiation through indirect pattern [3] and upregulates Cx43 expression in the niches, respectively. Based on these observations, we hypothesize that androgen may also promote SPC differentiation through regulating gap junction during intercellular interaction (Fig. 5). Evidence from Cx43-Sertoli cell condition knockout mice exhibited that spermatogenesis was blocked, differentiated spermatocytes and spermatids were deficient, but spermatogonia remained in seminiferous tubules [22], which is consistent with our hypothesis above, namely Cx43 functions in regulating spermatogonia differentiation, and this process is regulated by androgen. However, further experiments are required to verify this hypothesis.
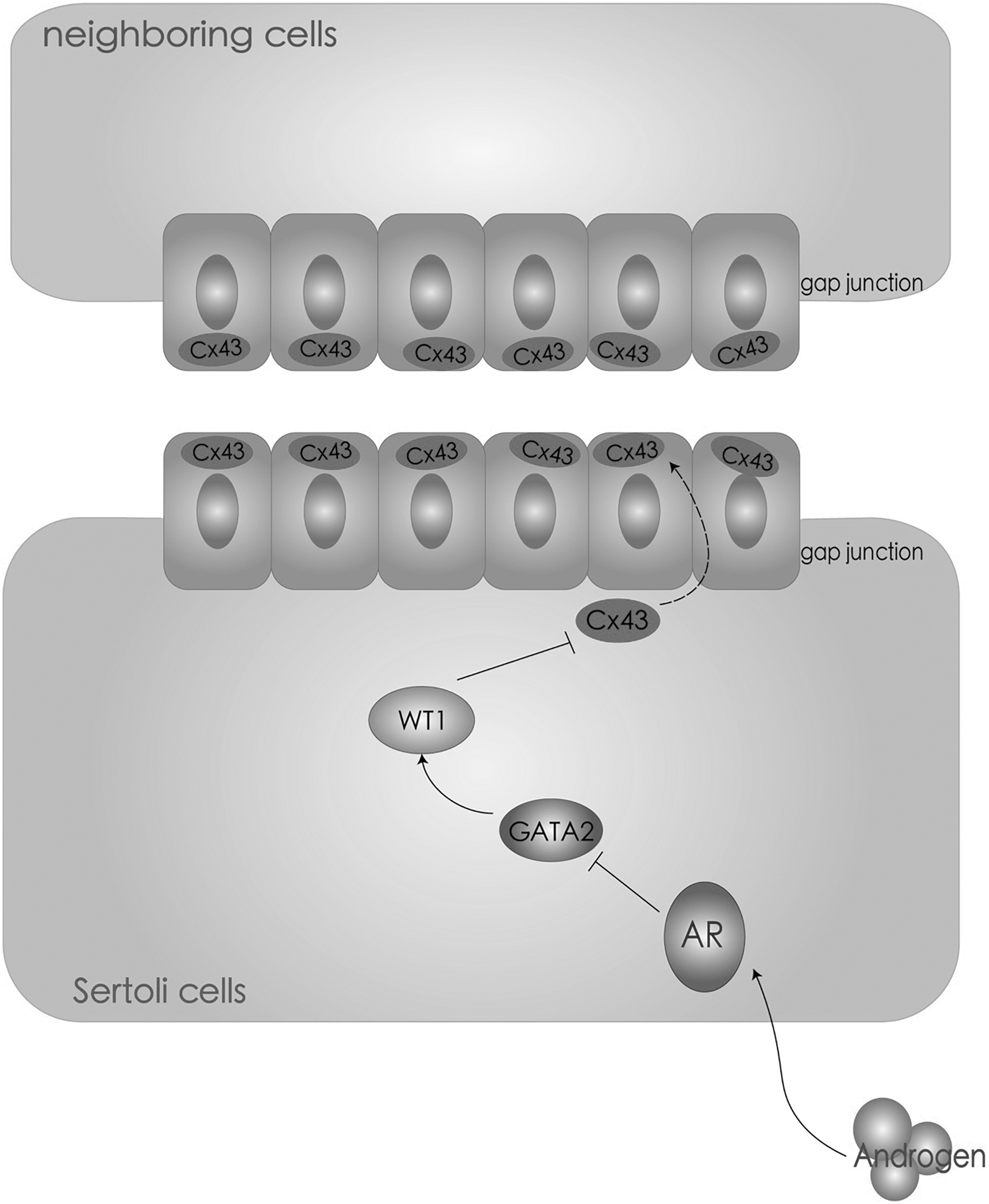
A putative signal pathway of androgen on regulating gap junction in testicular niche. Based on the data of this and previous studies, a regulatory pattern of androgen on gap junction in testicular niche is summarized: androgen signal regulates expression of Cx43 in Sertoli cells through GATA2 and WT1 and, subsequently, affects fates of neighboring cells.
Conclusions
This study investigates the regulatory role of androgen on Cx43 expression in Sertoli cells and confirms that WT1 directly inhibits Cx43 expression by binding on the promoter region of Cx43 gene, which preliminarily reveals the regulatory mechanism of androgen on Sertoli cell–spermatogonia interaction and may provide implications for basic research and clinic study.
Ethics Approval and Consent to Participate
All of the experimental procedures have been approved by the Ethics Committee at Nanjing Agricultural University.
Footnotes
Acknowledgment
The authors thank Professor Xiaodong Zhao (Shanghai Jiao Tong University) for suggestions on AR signal study and ChIP-seq data analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (81501316) and Fundamental Research Funds for the Central Universities in China (KYDS201807, KYTZ201602).