
Abstract
Over the past few decades, our knowledge about the function of the central nervous system (CNS) and astrocytes has improved, and research has confirmed the key roles that astrocytes play in the physiology and pathology of the CNS. Here, we reviewed the intrinsic and extrinsic mechanisms that regulate the development of astrocytes, which are generated from radial glial cells. These regulatory systems modulate various signaling pathways and transcription factors. In this review, four stages of astrocyte development—specification (patterning and switch), migration, proliferation, and maturation, are discussed. In astrocyte patterning, VA1–VA3 domains create the astrocyte subtypes by differential expression of Slit1 and Reelin in the spinal cord. In the brain, patterning creates several astrocyte subtypes by different organizing centers. At the switch step, the janus kinase-signal transducer and activator of transcription pathway governs the transition of neurogenesis to gliogenesis. Bone marrow protein and Notch pathways are also important players of the progliogenic switch. Intrinsic regulation is mediated by DNA methylation transferases, and polycomb group complexes can intrinsically affect the development of astrocytes. In the next stage, these cells proliferate and migrate to their final location. Astrocyte maturation is accomplished through the development of cellular processes, molecular markers, and functions.
Introduction
The structural and functional complexity of the central nervous system (CNS) can be explained by the presence of different specialized cell types that comprised neuronal and non-neuronal cell populations [1]. In 1856, the German pathologist Virchow confirmed the presence of non-neuronal cells in the CNS [1] and named them “neuroglia” [2].
Glial cells are composed of macroglia (astrocytes and oligodendrocytes) and microglia, which constitute at least 50% of the cells in the human brain [3]. Several studies in rodents and humans have shown that the ratio of glial cells to neurons may reflect the complexity of the CNS during evolution [4].
Astrocytes are the most abundant macroglia in the brain and spinal cord [5]. The results of recent studies have confirmed that astrocytes are as important as neurons for the physiological function of the CNS. Astrocytes can sense and respond to neuronal activity at synapses because of their unique phenotypical characteristics. Astrocytes appear to release gliotransmitters to modify the intensity of inhibitory and excitatory synaptic transmissions [6].
Astrocytes develop from radial glial cells (RGCs). RGCs are bipolar-shaped, early multipotent precursors generated from neuroepithelial cells of the neural tube that create both neurons and glia [7 –9]. RGCs generate quiescent neural stem cells (B1 cells) on the walls of the embryonic neural tube (future brain and spinal cord) [10 –12]. B1 cells divide into intermediate progenitors (IPs) (also known as C cells) and produce numerous immature neurons (or A cells) (Fig. 1A) [13]. Time-lapse imaging has shown that RGCs in the brain often undergo asymmetric self-renewal division to produce neurons or IP cells [14].
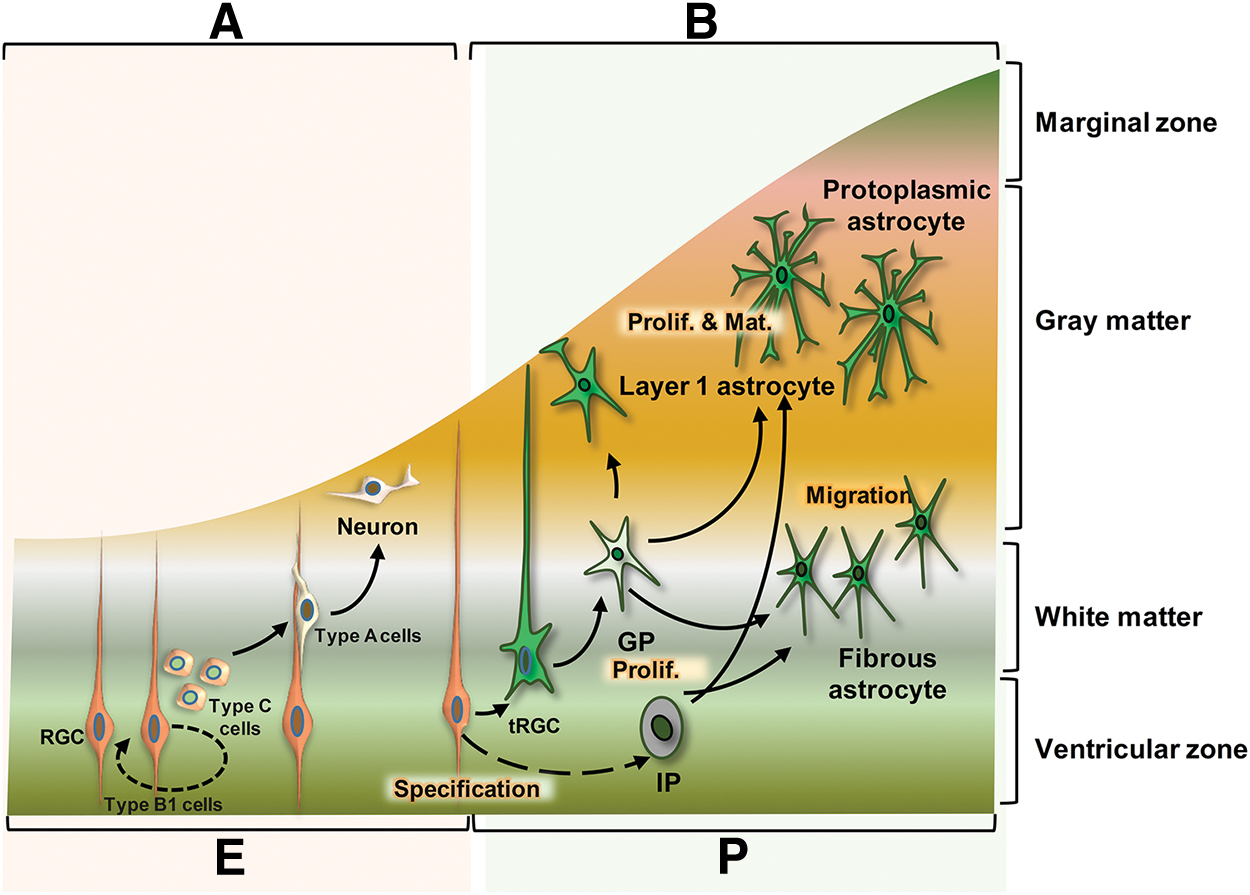
Neurogenesis and gliogenesis in the embryonic and postnatal stages.
Transformed B1 cells that generate glial progenitor (GP) cells are produced following removal of their apical processes and migration of B1 cells to the surface of the pia mater (Fig. 1B) [11]. The asymmetric division of B1 cells produces IPs in the subventricular zone (SVZ) or in the germinal zone of the spinal cord. Division of GPs and IPs produces immature proliferative astrocytes (the first proliferation wave) (Fig. 1B). These immature cells migrate to their final location to generate astrocyte subtypes—layer 1 astrocytes and protoplasmic astrocytes in the gray matter and fibrous astrocytes in the white matter. At this stage, immature astrocytes start a second proliferation wave and eventually mature (Fig. 1B). It has been reported that in the developing spinal cord, radial glia also settle in the ventricular zone (VZ) and generate neurons and astrocytes [15].
In astrocyte development, temporal and spatial mechanisms play very important roles similar to those observed during neurodevelopment [11]. Here, we discuss the first, types, and functions of astrocytes in the mature CNS and review these mechanisms during four stages: (1) specification, (2) proliferation, (3) migration, and (4) functional maturation (Fig. 2).

Summary of mechanisms of astrocyte development. Temporal and spatial intrinsic and extrinsic involved in the development of astrocytes These mechanisms are categorized in four stages: specification, proliferation, migration, and functional maturation. Color images are available online.
Astrocyte Heterogeneity
Astrocytes are categorized into two broad subtypes, protoplasmic and fibrous, based on cell morphology (“morphological diversity”) and their anatomical location within the CNS (Fig. 3) [16]. This morphological diversity is accompanied by “molecular diversity” that can serve as “functional diversity” in these subtypes. Protoplasmic astrocytes have highly branched bushy processes located in the gray matter closely associated with neuronal cell bodies and synapses (Fig. 3A) [17]. Protoplasmic astrocytes typically express S100β, a calcium-binding protein, whereas glial fibrotic-associated protein (GFAP), an intermediate filament protein, is hardly expressed by these cells (Fig. 3A) [17,18].
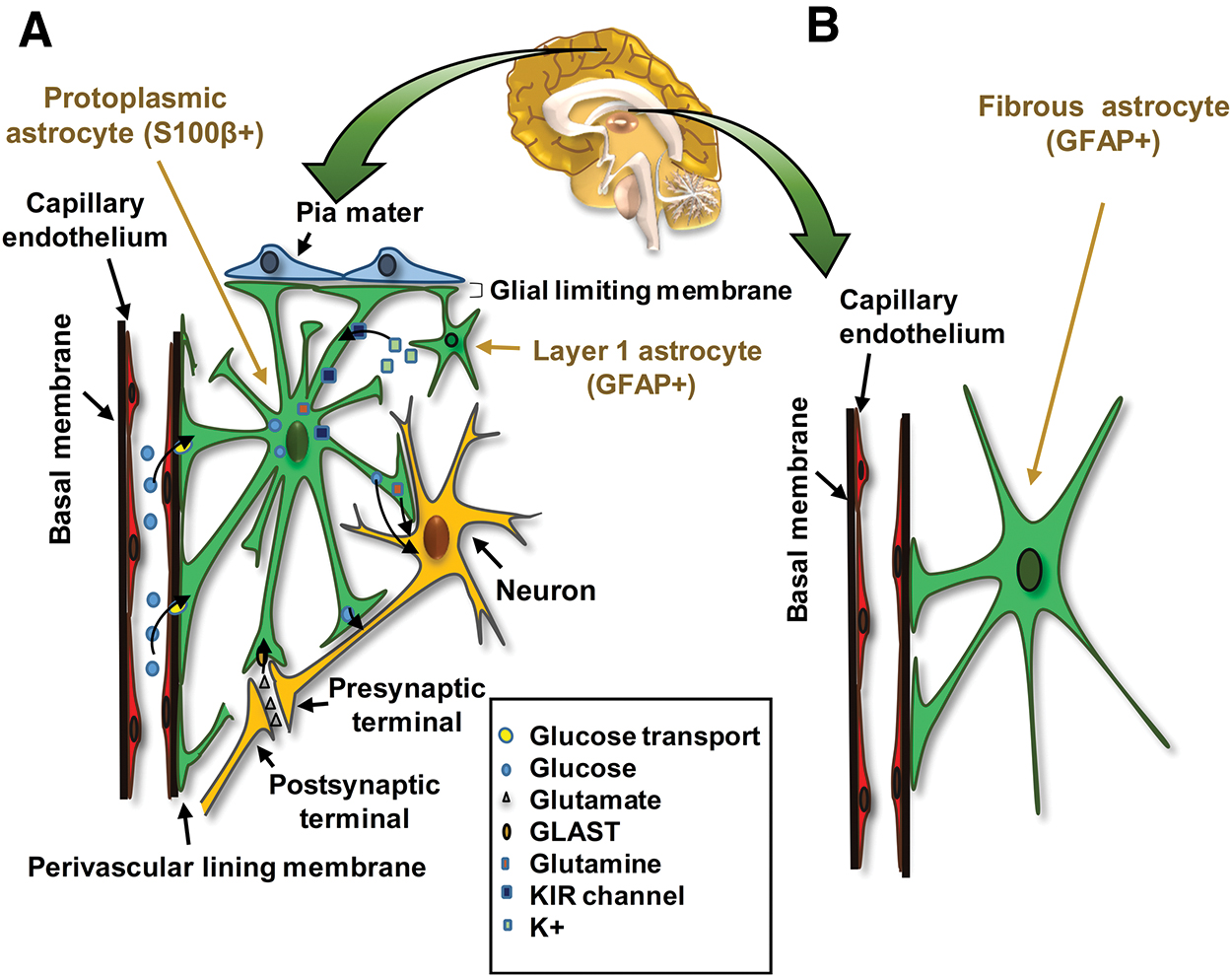
Two basic subtypes of astrocytes.
Protoplasmic astrocytes participate in the generation of the blood–brain barrier (Fig. 4) [17 –19]. The processes of these astrocytes extend onto endothelial cells, pericytes, and neurons. Astrocytes and vascular cells are mainly connected through integrins and connexins (CX40 and CX23) (Fig. 4). On the contrary, astrocytes are connected together through gap junctions (CX43, CX30). These interactions are very important for their normal function and for other cells in the CNS. Abnormalities in connexin expression can cause disorders in this system [20]. The results of several studies illustrate changes in astrocyte connexin expressions in Alzheimer's disease (AD), gliomas, autism, and brain ischemia [21]. Protoplasmic astrocytes regulate synaptic functions through their junctions with synapses (neuroligin/neurexin), usually forming tripartite synapses in the synaptic cleft (Fig. 4) [22,23].

The perisynaptic processes and perivascular processes of astrocytes. Processes of astrocytes are extended on endothelial cells, pericytes, and neurons. Integrins and connexins (CX40 and CX23) are the main attachments to the vascular cells. Astrocytes connect together through gap junctions (CX43 and CX30). Astrocytes surround the neuronal synaptic cleft with perisynaptic processes and make a tripartite synapse. CX, connexin. Color images are available online.
Recently, it has been revealed that protoplasmic astrocytes can be converted to “reactive” astrocytes during an injury [24]. These reactive astrocytes also express protein markers of mature “resting” astrocytes that include GFAP, S100β, aquaporin-4, connexins, and Aldh1L1 proteins [24,25]. It appears that astrocytes isolated from the injured brain can re-enter the cell cycle. Protoplasmic astrocytes in the injured tissue can dedifferentiate [26]. These reactive astrocytes are found in AD, amyotrophic lateral sclerosis (ALS), Parkinson's disease (PD), multiple sclerosis [20], and spinal cord injuries [27].
Fibrous astrocytes are the second subtype of astrocytes. They have straight and long processes, are located in the white matter, and associated with neuronal axons [8,28,29]. Fibrous astrocytes express higher levels of GFAP compared with protoplasmic astrocytes (molecular diversity) [16]. This astrocyte subtype is associated with blood vessels and acts via processes similar to those of protoplasmic astrocytes, but its exact function is not fully understood (Fig. 3B) [28]. Recent studies have shown that a mutation in the GFAP gene can cause abnormal white matter in Alexander's disease [30].
A specialized astrocyte subtype is present in the first layer of the cerebral cortex. Although it has a morphology similar to that of protoplasmic astrocytes, its GFAP expression is similar to fibrous astrocytes. The cellular processes of these astrocytes and end feet of protoplasmic astrocytes construct the glial limiting membrane to cover the cerebral parenchyma (Fig. 3A) [8].
Different morphological classes of astrocytes have been described in mammals [2,17]. Although the mechanisms that control this diversity have not been exactly determined, studies have recently shown that the loss of connexins or other attachment proteins in protoplasmic astrocytes alters the morphology of these cells, directly influences synapse formation, and generates different morphological and functional astrocytes [16,31,32]. Abnormal alterations in astrocyte morphology appear to be widely correlated with pathophysiology, as they are detectable in numerous brain disorders [16].
Astrocyte Development
In this review, astrocyte development is explained in four stages—specification, proliferation, migration, and maturation. We intend to describe the signaling pathways and transcription factors involved in specification, and explain the main pathways in immature astrocyte proliferation, migration, and maturation (summarized in Table 1 and Fig. 2).
The Different Steps of Astrocyte Development with Main Signaling Pathways, Specific Proteins, and Some Abnormalities or Neurodegenerative Disorders that Occurred in Each Step
BMP, bone marrow protein; EGFR, epidermal growth factor receptor; FGF, fibroblast growth factor; GFAP, glial fibrotic-associated protein; GLAST, GLutamate ASpartate Transporter; GluR, glutamate receptor; HBEGF, heparin-binding EGF-like growth factor; JAK-STAT, janus kinase-signal transducer and activator of transcription; NGN, neurogenin; NOTCH, a single-pass transmembrane receptor protein; RAS/MAPK, MEK/ERK (mitogen-activated protein kinase-extracellular signal-regulated kinases, also known as the RAS/MAPK pathway), or MEK/ERK pathway; Shh, sonic hedgehog; TGF-α, transforming growth factor-α.
Astrocyte specification
Several spatial and temporal mechanisms are involved in the generation of neuronal and glial cell types in the neural tube. These regulatory systems were thoroughly described by Rowitch and Kriegstein in 2010 [11]. Although these mechanisms in the brain and spinal cord are similar, they occur at different times. The neural tube is divided into different domains, patterned during the neurogenic period, and preserved in gliogenesis [11]. The gliogenic switch is tightly regulated and occurs in the developing spinal cord and cortex at E12.5 and E16–18, respectively [33]. Recently, studies have reported that patterning and the gliogenic switch are controlled by both extrinsic environmental cues that advance astrogenesis in neural progenitor cells (NPCs) and intrinsic mechanisms to decrease neurogenic capacity and increase astrogenic competence during development [33].
Embryonic pattern formation and gliogenesis
Interestingly, during CNS development, neurons and glial cell subtypes are only generated from the same progenitor cells in the embryonic neural tube [34]. There are several mechanisms that can control patterning and specification in the CNS that lead to the emergence of multiple cell fates [35]. Different combinations of transcription factors (named “progenitor proteins”) [36] are expressed depending on the location of a cell along the anterior/posterior (AP), rostral/caudal, or dorsal/ventral (DV) axes of the neural tube. Thus, the positional identity of cells can regulate molecular diversity and the generation of neuronal and glial subtypes.
Combined expression of these transcription factors generates diverse cell types at different positions that determine the early step of neural tube patterning [35]. According to research, Pax6, Nk2 homeobox 2 (Nkx2.2), and oligodendrocyte transcription factor 2 (Olig2) are key factors that play important roles in the creation of distinctive domains (Fig. 5). Differential expression of these progenitor proteins leads to the generation of p3, pMN, and p2–p0 domains along the DV axis of the neural tube in the spinal cord. The p1–p3 domains generate VA1–VA3 in the white matter [29,37].
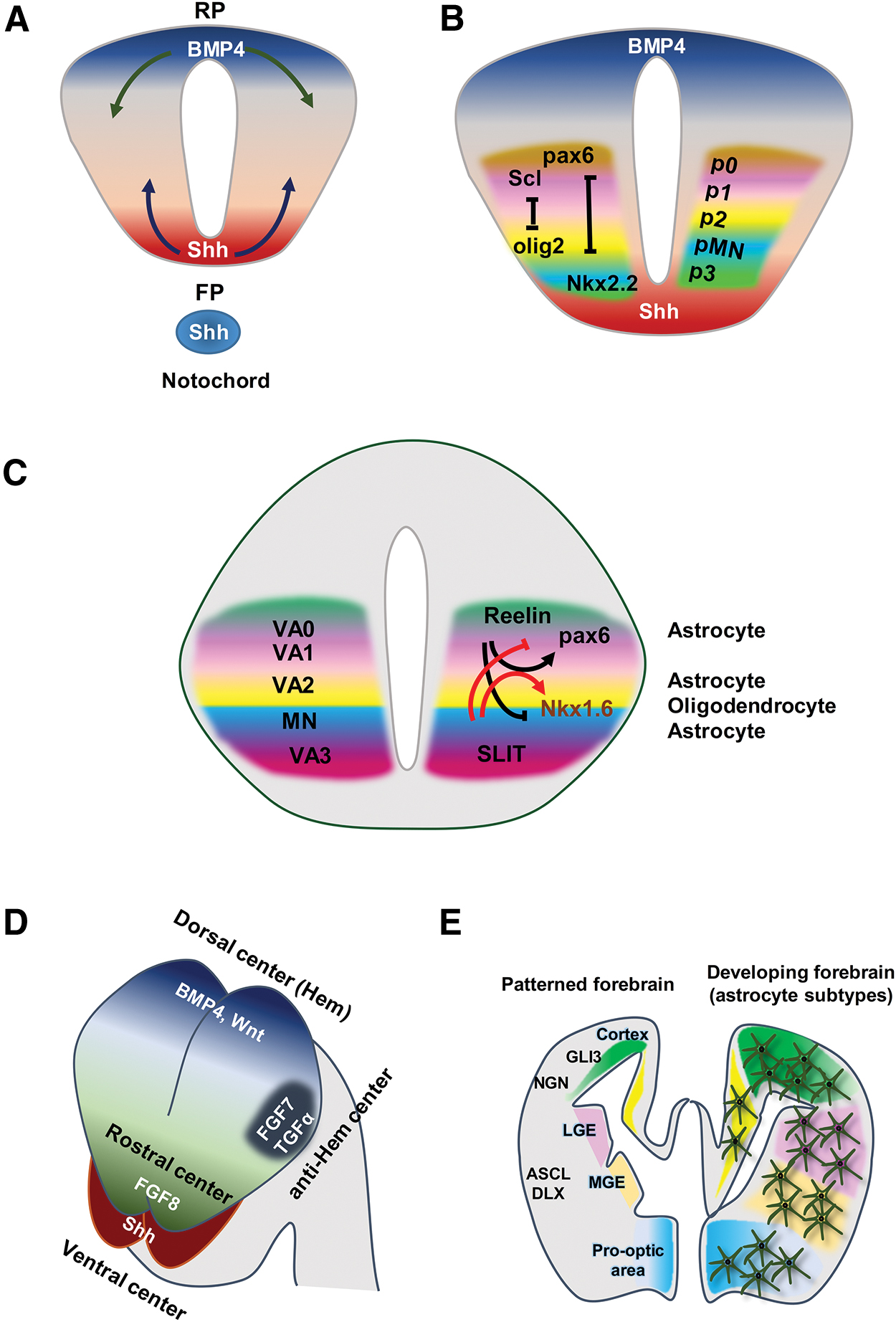
Patterning of the neural tube generates unique domains for astrocyte progenitors in the spinal cord and brain.
Studies have shown that differential expressions of progenitor proteins in the spinal cord depend on organizing tissue signals. One of the most important signaling pathways in patterning is sonic hedgehog (Shh), which is secreted by the notochord and floor plate. The roof plate secretes bone marrow protein 4 (BMP4), another important mediator in patterning (Fig. 5A) [38]. Together, these patterning molecules establish a gradient of morphogenic activities that result in the positional identity of the DV axis of the spinal cord. They order the expressions of progenitor proteins that will further cross-repress each other and form the tight boundaries or domains made by diverse subtypes of neurons (Fig. 5B). These domains are conserved during gliogenesis and regulate the generation of astrocyte subtypes [30,38,39].
These domains should generate subtypes of astrocytes after the switch from neurogenesis to astrogenesis. Previous studies have found that astrocytes are derived from three regions of the white matter in the spinal cord, from dorsal to ventral along the DV axis (VA1, VA2, and VA3), as a “segmental model” for glial cell development (Fig. 5C) [30]. Functional analyses of transcription-factor-encoding genes have also shown the presence of gliogenic domains in the spinal cord. Although the absence of Olig2 ventrally extends the p2 domain and generates astrocytes rather than oligodendrocytes, the loss of Scl (Tal1) causes dorsal Olig2 expression and increases oligodendrocyte production [40]. Differential expressions of Slit1 and Reelin in the VA1–VA3 domains cause molecular diversity (Fig. 5C).
In the VA1 domain, the expression of Pax6/Reelin produces more dorsally located astrocytes. Expressions of Nkx6.1 and Slit1 in the VA3 domain induce the generation of astrocytes in the ventral spinal cord, and astrocytes located between the ventral/lateral white matter are produced from the VA2 domain (Pax6+, Nkx6.1+, Reelin+, and Slit1+) (Fig. 5C). Pax6 is essential for Reelin expression and production of astrocytes from the VA1 and VA2 domains [40]. Reelin inhibits the expression of Nkx6.1, and Slit1 can suppress Pax6 expression (Fig. 5C). These interactions dictate the pattern of diverse domains in the neural tube and generate astrocyte subtypes.
Similar to the spinal cord, spatial domains are generated by patterning molecules and homeodomain protein expression within the forebrain [16]. There are four main organizing centers in the forebrain: (1) a dorsal center, the cortical hem, which secrets BMPs and Wnts; (2) a ventral center source of Shh; (3) a rostral center that is a source of fibroblast growth factor 8 (FGF8); and (4) anti-hem region that secretes transforming growth factor-α (TGF-α) and FGF7 (Fig. 5D). These patterning molecules create a gradient of morphogenic proteins and the positional identity of the DV axis (Fig. 5E). The gradient of morphogenic proteins induces the expression of progenitor proteins (GLI3, NGNs, ASCL, and DLXs) in different domains and produces diverse subtypes of glia (Fig. 5E).
Research is ongoing to determine the detailed mechanisms of region-specific patterning in other regions of the CNS, which are unknown. It should be kept in mind that these domains generate both protoplasmic and fibrous astrocytes. Thus, astrocytes by diverse morphological features have a common developmental origin [16]. Different subtypes of astrocytes appear to be produced by the local environment where they reside [41].
The exact regulation of these domains is very important. For example, abnormal expression of Olig2 leads to premature gliogenesis that is seen in Down's syndrome patients (Table 1).
Switch from neurogenesis to astrogenesis
The switch from neurogenesis to astrogenesis in the spinal cord and brain results in inhibited expression of neuronal markers and, on the contrary, initiation of astrocyte marker expressions [42].
Neurogenesis stage
The expression of neuronal markers is initiated by recruiting several pathways, and followed by the scheduling of neurogenesis. The signaling pathway that triggers neuronal gene expressions is the receptor tyrosine kinase (RTK) pathway, which promotes direct activation of neuronal genes such as a-tubulin and math2, and causes phosphorylation of the C/EBP family (Fig. 6A-1) [43,44]. Activation of the RTK pathway directly inhibits the janus kinase-signal transducer and activator of transcription (gp130-JAK-STAT) pathway, the main signaling pathway in astrogenesis (Fig. 6A-2). Thus, activation of mitogen-activated protein kinase-extracellular signal-regulated kinases (MEK/ERK) in the RTK pathway promotes neurogenesis and blocks astrogenesis during the neurogenesis stage. Phosphorylated C/EBP is linked to P300 and this complex cannot bind the GFAP promoter and inhibit the expression of this astrocyte marker (Fig. 6A-3) [42,45].
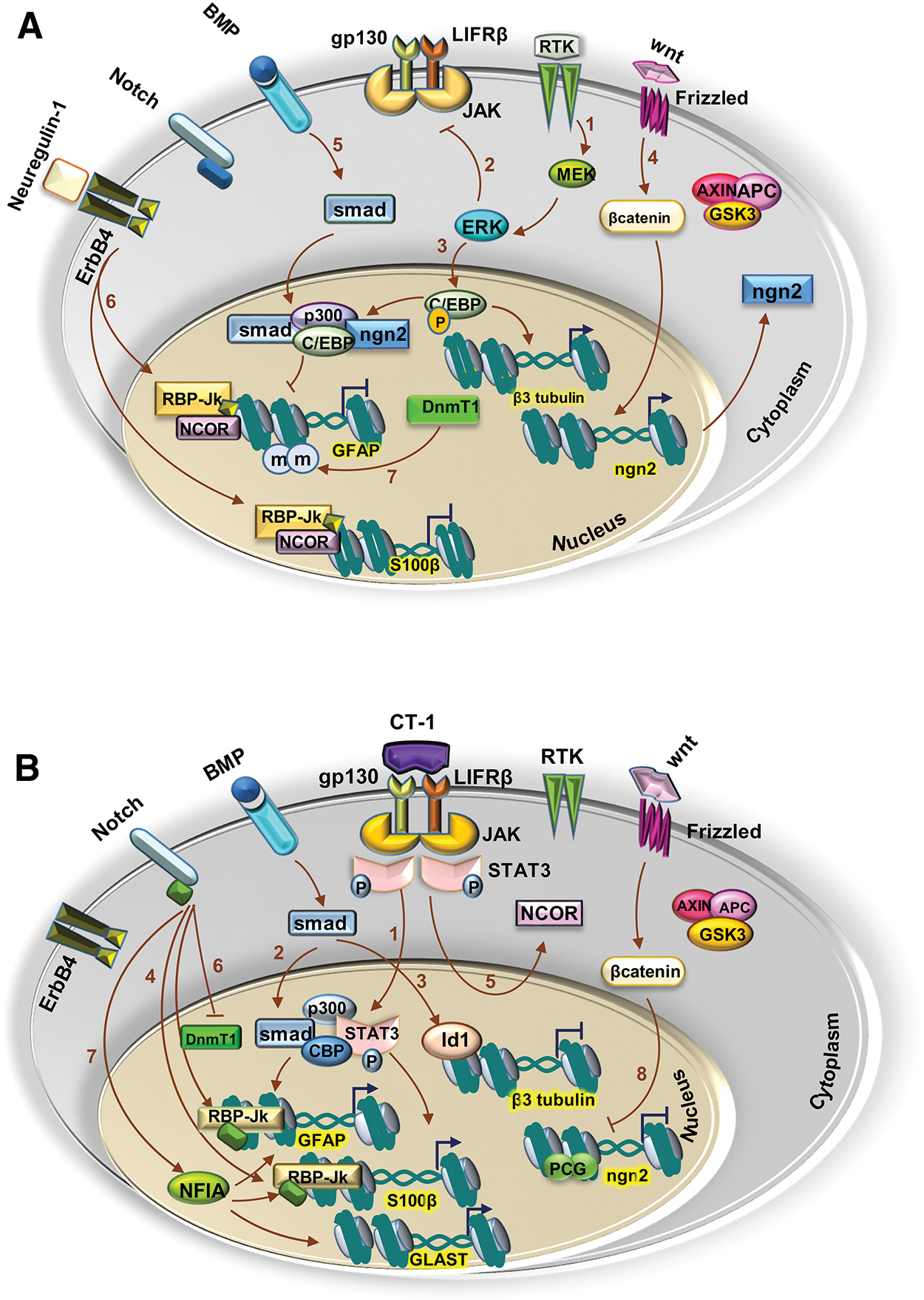
Signaling pathways involved in the induction of neurogenesis and astrogenesis.
Many studies have reported the critical role of neurogenin 1 (NGN1) and NGN2 in neurogenesis, neuronal differentiation, and regulation of gliogenesis [46]. Expressions of NGN1 and NGN2 are activated by Wnt signaling in NPCs and induce neuronal differentiation (Fig. 6A-4). Like C/EBP, NGN links to P300 and inhibits attachment of STAT3 (a downstream protein in the gp130-JAK-STAT pathway) to the GFAP promoter and suppresses expression of the astrocyte-specific genes. Smad1 attaches to the C/EBP/P300/NGN complex and reinforces the prevention of GFAP expression in the presence of NGNs, following activation of BMP signaling (Fig. 6A-5) [46].
Another factor that inhibits gliogenesis during the neurogenic period is neuregulin-1 [33]. Neurogulin-1 interacts with protein N-CoR, a corepressor. The importance of neurogulin-1 in the inhibition of gliogenesis was demonstrated in an embryonic forebrain from N-CoR−/− mice. It appears that the inhibitory effects of neuregulin-1 depend on the interaction between N-CoR and a Notch effector, RBP-Jk. Binding of neuregulin-1 to its ErbB4 receptor leads to cleavage, release of the intracellular domain of the ErbB4 receptor, and interaction with the intracellular domain by the N-CoR protein. Translocation of the intracellular domain/N-CoR complex to the nucleus and generation of the intracellular domain/N-CoR/RBP-Jk complex suppress GFAP and S100β expression (Fig. 6A-6) [33].
Furthermore, methylation of the GFAP promoter in the STAT3 binding site by DNA methylation transferase 1 (DnmT1) inhibits the attachment of STAT3 to this site (Fig. 6A-7). In 2005, Fan et al. reported that conditional knockout of the DnmT1 gene in neural precursor cells decreased the number of neurons and promoted astrogenesis. DnmT1 also regulates the gliogenic potential of neural precursor cells through the methylation of genes in the gp130-JAK-STAT pathway [47].
Overall, in the neurogenic stage, several elements work together to regulate the neuronal potential of neural stem cells (NSCs) by ensuring the inhibition of astrocyte-specific genes and genes of the gp130-JAK-STAT pathway.
Astrogenesis stage
In this stage, the neurogenic pathways must be inhibited and astrogenic pathways are activated. The JAK-STAT pathway is critical for the gliogenic switch. This pathway is activated by attachment of a particular subfamily of cytokines to their receptors [46] such as ciliary neurotrophic factor, leukemia inhibitory factor, and cardiotrophin-1 (CT-1), which results in heterodimerization of LIFRβ and gp130. Barnabé-Heider et al. have indicated that CT-1 is a key ligand for LIFRβ and gp130 [48]. The onset of gliogenesis is determined by neurons that produce CT-1, a major extrinsic element in the regulation of gliogenesis initiation [46].
When CT-1 is linked to its receptors, the JAK protein activates and phosphorylates the STAT3 protein (Fig. 6B-1). Reduction of astrocyte formation in gp130−/− or LIFRβ−/− neural precursors has been reported by in vitro and in vivo studies [48,49]. GF, AP, and S100β expressions depend on the binding of STATs to p300/CBP, the formation of the STATs/p300/CBP complex, and the binding of this complex to its promoters [43,44].
On the contrary, at low NGN1 expression levels and in the presence of BMP and gliogenic cytokines, the Smad/p300/CBP/STAT complex induces the expression of gliogenic genes (Fig. 6B-2) [46]. Under these circumstances, BMPs induce the expression of inhibitory factors such as Id1, as antagonists of neurogenic genes, to ensure repression of neuronal expression (Fig. 6B-3) [43,46].
By activation of Notch, its intracellular domain is cleaved and translocated to the nucleus, where it interacts with RBP-Jk to form a transcriptionally active complex [43]. When the JAK-STAT pathway is not activated, RBP-Jk binds to N-CoR and cannot bind the GFAP promoter. Following activation of the JAK-STAT pathway, RBP-Jk can bind to the GFAP promoter and GFAP is expressed (Fig. 6B-4). At this point, activation of the JAK-STAT pathway leads to the translocation of N-CoR to the cytoplasm, where it determines the response of glial genes to environmental stimuli (Fig. 6B-5). Dephosphorylation of STAT3 in this stage leads to excessive neurogenesis and decreased astrogenesis, present in human neurodevelopmental disorder Noonan syndrome (Table 1) [45].
Notch signaling can regulate gliogenesis by epigenetic changes in the GFAP promoter. Demethylation occurs when Notch dissociates DnmT1 from the GFAP promoter (Fig. 6B-6). It has been reported that Notch signaling can induce the expression of nuclear factor IA (NFIA). NFIA is a very important regulator of the neuron-to-glia switch. NFIA is an essential, sufficient factor that promotes glial specification [38]. This factor is necessary for the stimulation of expression of the early glial marker, GLAST (Fig. 6B-7) [50]. NFIA can suppress neurogenesis via the preservation of Notch signaling [38]. NFIA, in conjunction with another important factor, SOX9, creates a complex that regulates the expression of a set of genes that promote gliogenesis [38]. On the contrary, results from several studies have indicated that NFIA expression induced the expressions of GFAP and S100β (Fig. 6B-7) [38,50,51].
As previously mentioned, NGNs inhibit the binding of STAT to the GFAP promoter [35]. In the absence of NGN2, the STAT/p300/CBP complex is formed and GFAP is expressed. Moreover, NGN expression is induced by Wnt signaling in the neurogenic stage. After the neuron-to-glia transition, NGN is still present in neuroepithelial cells but it cannot activate NGNs in these cells [35]. After neurogenesis, intrinsic changes occur in histone H3 of the NGN1 and NGN2 promoters of NPCs. At this stage, hypoacetylation and hypermethylation occur in histone H3. These epigenetic changes in NPCs are mediated by polycomb group complexes (PcG) (Fig. 6B-8). Knockout of key components of the PcG (eg, Ring1B and Ezh2) delays the production of astrocytes [35]. Thus, despite the continuous presence of Wnts, the expression of NGNs is inhibited due to intrinsic epigenetic changes (Fig. 6B-8) [35].
In summary, at this stage, the neural tube is patterned, astrogenesis occurs, and astrocytes are formed from the different domains. Then, these cells proliferate and migrate to their final location in the spinal cord and the brain.
Astrocyte proliferation
Two waves of astrocyte proliferation have been detected in both the developing spinal cord and cortex: before migration in the lateral VZs and after migration in their final location (local division, Fig. 7A) [8,16,17,52]. In the lateral VZs, radial glia divide asymmetrically and proliferate to produce astrocyte progenitors before migration (Fig. 7A-1). These astrocyte-restricted precursors migrate to their final location where they divide symmetrically to generate more astrocytes (Fig. 7A-2) [42]. RGCs may transform into GP after their terminal division. Astrocyte proliferation occurs in the spinal cord earlier than the forebrain (caudal to rostral patterning) [42]. In 2012, scientists used green fluorescent protein to label the SVZ RGCs of mice at postnatal days (P) 0–2, and showed that only a very small percentage of cortical astrocytes were generated from SVZ RGCs [42]. Bandeira et al. demonstrated a six- to eightfold increase in the astrocyte population in the brain during the first 3 postnatal weeks [53]. Local division generates about half of the mature cortical astrocytes; however, the exact timing of this process is unclear [54].

Astrocyte proliferation and migration.
Some studies have revealed that the major pathway in astrocyte precursor proliferation is the RAS/MAPK pathway or MEK/ERK pathway in the spinal cord and cortex [1,55]. Activation of the epidermal growth factor receptor (EGFR) induces the RAS/MAPK pathway (Fig. 7B). Astrocytes are one of the cells expressed by EGFR and its ligands (epidermal growth factor [EGF] or heparin-binding EGF-like growth factor [HBEGF]) [56]. Results of studies have shown that HBEGF has an autocrine function on astrocytes and can improve the expressions of HBEGF and EGFR (Fig. 7B). HBEGF binds its receptors (EGFR, Erbb2, and Erbb4) and induces the RAS/MAPK pathway (Fig. 7B) [56]. In this pathway, BRAF, a member of the Raf kinase family, activated and activees ERK1/2. These factors are critical for astrocyte proliferation (Fig. 7B) [30,55]. Loss of BRAF function results in a decrease of 20% in the astrocyte population, while ectopic expression of this factor leads to astrocyte hyperproliferation [55,57]. In some neurodegenerative disorders and in gliomas, increased expressions of HBEGF and EGFR in the astrocytes may cause a reactive state with the properties of immature astrocytes (Table 1) [56].
Astrocyte migration
Astrocyte progenitors migrate into gray matter and white matter to differentiate into protoplasmic and fibrous astrocytes (anatomical diversity), respectively. Although astrocyte precursors migrate along with the radial glial processes, their migration is limited. Since RGCs have lost their processes during the early postnatal stages, a secondary migration may occur. Nevertheless, the exact molecular and cellular mechanisms that regulate astrocyte precursor migration remain poorly defined [37].
Migration usually includes cytoskeleton rearrangement, the extension of the cell membrane in front of the cells, and the formation of protrusions or filopodia. Then, new adhesion sites are recognized at the front, followed by contraction of the cell body and detachment of the posterior portion of the cell. Migrating astrocytes are very flat, firmly adherent, and polarized. Astrocyte polarization is characterized by the formation of a protrusion of ∼100 μm in the direction of the migration. This protrusion is full of elongated microtubules. During microtubule rearrangement, reorientation of centrosomes in front of the nucleus occurs and forms the microtubule organizing center (MTOC) that consequently determines the direction of cell migration. Centrosome position in astrocytes appears to be a good indicator of the orientation of the protrusion; therefore, it reflects the orientation of migration in the polarized cells [58].
Migration starts in the presence of external cues, including the extracellular matrix (ECM), soluble factors, and adjacent cells. Integrin commonly responds to these inducers. In addition, integrin activates intracellular signaling pathways that lead to cytoskeletal rearrangement, structural reorganization, and morphological polarization [58,59]. During migration, several integrins are activated in different cells. Specifically, β1 integrin is activated in astrocytes [19]. CDC42 and/or RAC (small Rho GTPases) are activated by adaptor proteins after the activation of β1integrin in astrocytes by the ECM (Fig. 7C-1). RAC regulates actin polymerization and membrane protrusion; therefore, it is required at the front of the cell [60].
RAC and CDC42 are important for the regulation of cytoskeleton microtubules. One of the targets of CDC42 is mPar6, the mammalian homologue of the Caenorhabditis elegans PAR6 protein (Fig. 7C-2). This protein is responsible for the polarity of the embryonic C. elegans cells and contains a motif that binds CDC42/RAC. mPar6 plays an important role in cell polarity and acts along with another protein, atypical protein kinase C (aPKC). In astrocytes, mPar6 forms a complex with PKCζ, a subtype of aPKC (Fig. 7C-2). Etienne-Manneville and Hall have reported that the mPar6/PKCζ complex is not essential for protrusion formation, but it is required for the correct formation of MTOC and orientation of the protrusion [61]. This complex interacts with glycogen synthase kinase-3β (GSK-3β) to promote polarization of centrosomes and control the direction of migration. GSK-3β interacts with another protein, adenomatous polyposis coli protein (APC), which stabilizes microtubule arrays and reorients the MTOC using dynein (a motor protein) (Fig. 7C-3) [62]. On the contrary, after the activation of RAC/CDC42, IQGAP1 accumulates at the front and crosslinks actin filaments [62]. Several studies have indicated that IQGAP1 directly interacts with APC; both IQGAP1 and APC are at the leading edge. IQGAP1 appears to anchor APC to actin filaments (Fig. 7C-4) [62].
In astrocytes, S100β interacts with IQGAP1 and controls the rearrangement of actin filaments. The polymerization of actin filaments in the vicinity of the plasma membrane creates pushing forces that lead to the formation of the membrane protrusion (Fig. 7C-5) [63].
After activation of RAC/CDC42, another target, Ser/Thr kinase P65PAK, is activated. In migration, integrin-dependent matrix adhesion is essential at the front and in the body of the cell but becomes disassembled at the rear of the cell to allow cell migration. P65PAK regulates this local adhesion (Fig. 7C-6) [58].
Astrocyte maturation
After proliferation and migration of astrocytes to their final location, the astrocytes become morphologically and molecularly mature. Astrocyte development is followed by the generation of branches and fine processes (termed “peripheral astrocyte processes”) that complement morphologic diversity and the expression of several markers that complete molecular diversity. Peripheral astrocyte processes comprise around 50% of the mature astrocyte volume and consist of various ion channels, ligand receptors, and transporters. These processes interact by synapses and activate several signaling pathways in astrocytes and neurons [64]. Astrocytes, by their processes, occupy a large, nonoverlapping domain that is <50 nm in diameter (Figs. 4 and 8A) [59]. It is now believed that astrocytes “tile” with one another by a mechanism that relies on dendritic tiling to completely cover the space between neurons [59,65]. How these domains are formed remains an intriguing and unexplored issue. These astrocyte domains generate “synaptic islands” around a single astrocyte and they can control the activity of all synapses within the astrocyte's unique spatial domain (Figs. 4 and 8A) [66]. Following an injury, breakdown of unique astrocyte domains may occur in the CNS due to reactive astrocyte overlap [67].

Astrocyte maturation.
The cellular processes of astrocytes develop during the first postnatal week where most of these processes appear as filopodia. During weeks 3 and 4 of postnatal development, astrocyte processes begin to branch. The distal cell processes become thinner and spongiform, and fine astrocyte processes that densely infiltrate the brain tissue to astrocytes display “mature morphology” [59].
In addition, the expressions of several important genes in astrocytes begin in this stage. The expressions of these factors are responsible for normal astrocyte function [59]. These genes include glutamate transporters such as glutamate transporter 1 (GLT-1) [29,59], CX43, and CX30 [21,29,59,68], and inwardly rectifying potassium channel Kir4.1 [22,45,69,70] also detected in astrocytes within 3–4 weeks after birth [59]. This molecular diversity with morphological and anatomical properties generates astrocyte heterogeneity and supports different functions of the astrocyte subpopulations. The expression of these markers and the growth of peripheral astrocyte processes in astrocytes complete the astrocyte development until 3–4 weeks after birth. Moreover, by the expressions of these proteins on the surfaces of the fine processes, astrocytes achieve their proper functions.
Different mechanisms regulate astrocyte maturation. Astrocyte-to-astrocyte contact is a very important mechanism in astrocyte maturation. As previously mentioned, EGFR and its ligands are expressed by astrocytes and induce proliferation in the astrocytes (Fig. 7B). In the maturation stage, BMP is activated and downregulates the expression of HBEGF (Fig. 8A-1). The decrease in HBEGF reduces proliferation. On the contrary, activation of BMP and a decrease in the MEK/ERK pathway induce CX43 expression (Fig. 8A-2). The expression of CX43 in immature astrocytes can induce the expression of another connexin protein, CX30 (Fig. 8A-3). Moreover, results of studies have indicated that the presence of neurons is necessary for the initiation of CX30 expression and improvement of CX43 expression [71]. There is the selective induction of CX30 in cells in close contact with neuronal cell bodies (Fig. 8A-4). Neuronal-specific signals appear to be important for CX30 expression [72]. These neuron-dependent inductions play important roles in the initiation of astrocyte gene expression during development in vivo. It is possible that neuronal extracellular components in perineuronal nets around neurons are involved in the induction of CX30 expression in astrocytes that are close to these perineuronal nets (Fig. 8A-4) [72].
Recent studies of the astrocyte gene profile showed the expressions of many neurotransmitter receptors on the surface of developing astrocytes to receive neuronal signals. During astrocyte maturation, glutamate induces the growth of astrocyte processes via the glutamate receptor (Fig. 8A-5) [59].
The dependence of the morphological variety of astrocytes on direct contact with neurons was confirmed by Stogsdill et al. in 2017. They reported that astrocyte neuroligin/neuronal neurexin interactions could affect the maturation of astrocyte morphology [31]. Three members of the neuroligin family (NL1, NL2, and NL3) are expressed on the astrocyte surface [31]. Neuroligins might be involved in the regulation of the growth of astrocyte processes as demonstrated in neuroligin KO mice. These mice had reductions in the branching processes, total branching length, and cell body sizes [73]. Mutations in neuroligins and neurexins have been detected in autism (Table 1) [74]. On the contrary, studies have shown that the expressions of Wnt and β-catenin protein were decreased in autistic brains. Wnt/β-catenin signaling may be due to astrocyte neuroligin/neuronxin interactions [73,75]. This interaction causes the expression of GLT-1 (EAAT2) and glutamate synthetase (GS), and can regulate astrocyte metabolism and the organization of astrocyte processes (Fig. 8A-6) [76]. Moreover, altered expression of GLT-1 in astrocytes is a common feature of the morphology of schizophrenia in the human brain (Table 1) [77]. In schizophrenia, a decrease in astroglial numbers and processes, or dystrophic or swollen astrocytes have been observed in different regions of the brain, cortical and hippocampal structures [77]. GLT-1 expression is significantly decreased in Huntington disease (HD) tissues in humans [77].
In addition, neurons control astrocyte morphology in the healthy adult brain via Shh, which can regulate the molecular and functional diversity of astrocytes (Fig. 8A-7) [78]. Neurons secrete Shh to control the properties of astrocytes that are in contact with them (Fig. 8A-7). Surprisingly, astrocytes respond differently to the Shh gradient across the CNS. In a subtype of astrocytes of the cerebellar cortex, Shh promotes ionotropic glutamate receptors (GluA1 and 4) and GLutamate ASpartate Transporter (GLAST), as well as potassium homeostasis (Kir4.1). Hippocampal astrocytes, in contrast, respond to Shh signaling by expression of Kir4.1 [78]. The decreased expression of Kir4.1, as well as decreased expression of GLT-1 and GS, is detected in epilepsy [77].
In addition to neurons, astrocyte maturation is affected by other cell types. Several studies have shown that vascular signals are key for the expression of Aqp4 in astrocyte end feet (Fig. 7A-8) [56]. Microglia and oligodendrocytes also significantly reduce cell proliferation in the maturation stage (Fig. 8A-9) [56].
Overall, these mechanisms must be highly regulated to generate a mature astrocyte (Fig. 8B).
Conclusion and Future Perspectives
Rapid progress in knowledge that pertains to astrocyte development and pathology has addressed important questions about the role of these cells in human neurological disorders and brain injury [79]. Vertebrate astroglia include a functionally and morphologically heterogeneous group of cells. Patterning relies on the local environment of glial precursors in the neural tube that produce diverse subtypes of astrocytes. These cells become specialized, proliferate, and migrate to specific microenvironments of the CNS and mature.
We have covered astrocyte development in four steps: specification, proliferation, migration, and maturation. Each step is subject to the regulation of extrinsic and intrinsic mechanisms that affect patterning, switch, migration, symmetric division, and maturation. In astrocyte patterning, p0–p3 domains produce VA1–VA3 domains, which make heterogeneous astrocytes by differential expression of Slit1 and Reelin [30,35]. In the brain, patterning is regulated by different organizers and it generates astrocyte subpopulations in different locations. At the neurogenesis stage, extrinsic mechanisms such as activation of RTKs and the Wnt pathway, as well as NGN1, inhibit gliogenesis and activate neurogenesis [45,46]. At this point, the GFAP promoter is methylated by DnmT1 and this intrinsic mechanism inhibits expression of this astrocyte marker [44].
In contrast, during astrogenesis, the JAK-STAT pathway is activated by CT-1 that is secreted by new neurons. Activation of the JAK-STAT pathway induces the expression of proastrocyte genes such as GFAP and S100β [43,46]. BMP and Notch pathways are also important players of the progliogenic switch [43]. Intrinsic regulation of the switch occurs in histone H3 of the NGN1 and NGN2 promoters following activation of PcG [35].
Astrocyte proliferation occurs in two waves in the spinal cord and brain: (1) before migration where progenitors divide asymmetrically in the VZ and (2) after migration where cells divide symmetrically to generate more astrocytes [54]. Migrating astrocytes are extremely flat, firmly adherent, and polarized [58]. By activation of integrin, intracellular signaling leads to cytoskeletal rearrangements, structural reorganization, and morphological polarization [60].
Cellular processes and the expressions of molecular markers in astrocytes develop during the first postnatal weeks [59]. During weeks 3 and 4 of postnatal development, astrocyte processes and the expression of several genes complete astrocyte development and generate diversity in these astrocytes. Branches of these cells infiltrate into brain tissue, and the astrocytes acquire a mature morphology [59]. In particular, the study of astrocyte cellular diversity in conjunction with decoding the related functional heterogeneity would be helpful in providing a general understanding of the development of CNS networks under normal and pathologic conditions.
The developmental processes of astrogenesis are tightly regulated by several factors and pathways. Various studies have been conducted to identify the importance of astrocytes during injuries and neurodegenerative diseases [7,17,28,45,79]. It has been demonstrated that the properties of astrocytes change after an injury, and these new properties are controlled by a series of new regulatory factors, or by the same pathways under different conditions (embryo vs. injured adult) [65]. However, attention to the fundamental pathway of development in normal astrocytes, reactive astrocytes, and regenerative astrocytes may provide insight into their injury-associated responses.
In recent years, the feasibility of deriving functional astrocytes from embryonic stem cells and induced pluripotent stem cells can provide the opportunity to discover the exact roles for astrocytes in human diseases [5,6]. In addition, several studies have reported the high potential of astrocytes in transforming to other cells [59,80 –82]. Therefore, it is of particular interest to study CNS repair through in vivo reprogramming [80,83 –94]. By targeting astrocytes, in vivo reprogramming has been able to successfully generate glutamatergic neurons as well as GABAergic neurons [90,92,93,95]. Stem cell therapy has been used in various experimental settings, and cells were transplanted into an animal model after differentiation. However, in vivo transdifferentiation of astrocytes can provide new therapeutic methods for the repair of this system. This new approach is considered to be an effective regeneration strategy [91,96 –98]. Although this method can be a promising new approach in regenerative medicine, the challenges of improving this technology and translating it into clinical applications should be addressed.
Finally, since astrocytes are much more heterogeneous than previously suspposed, many questions remain unanswered. It is critical to use new genetic approaches to assess the genetic profiles of astrocytes in the developing, mature, and injured CNS.
An understanding of the precise regulatory mechanisms involved in astrocyte development and reactive astrocyte production can lead to the development of novel therapies against different neurodegenerative diseases such as AD, HD, ALS, and PD, and traumas such as spinal cord injuries. Moreover, direct conversion of somatic cells into astrocytes would be more appropriate.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a grant provided by the Royan Institute.