
Abstract
In this study, the roles of exosomes (Exo) from bone marrow mesenchymal stem cells (BMSCs) in attenuating early brain injury (EBI) in rat brain after subarachnoid hemorrhage (SAH) had been investigated. The male Sprague-Dawley rats (300–350 g) were used to establish the SAH model using endovascular perforation method. The animals were randomly divided into three groups: sham (n = 25), SAH+PBS (n = 42), and SAH+Exo groups (n = 33). At 1 h after SAH, Exo or phosphate-buffered saline (PBS) was administered by femoral vein injection. The effects of Exo on the mortality, neurological function, brain water content, and blood–brain barrier (BBB) were explored. Furthermore, the expressions of miRNA129-5p and high-mobility group box 1 protein (HMGB1) after Exo treatment were also detected. In addition, immunohistochemistry and western blot were applied to investigate the mechanism of Exo's effects. The results indicated that Exo could improve the neurological functions, reduce brain water content and maintain BBB integrity after SAH. After Exo treatment, the expression of miRNA129-5p was significantly increased, whereas the RNA level of HMGB1 was decreased. The protein levels of proinflammatory and proapoptosis factors, such as HMGB1, Toll-like receptor-4 (TLR4), tumor necrosis factor-α, and p53, were increased after SAH, which were significantly declined after Exo application. The results indicated that Exo from BMSCs could alleviate EBI after SAH through miRNA129-5p's anti-inflammation and antiapoptosis effects through quenching the activity of HMGB1-TLR4 pathway.
Introduction
Subarachnoid hemorrhage (SAH), a type of hemorrhagic stroke, has higher mortality and disability [1]. Early brain injury (EBI) occurs within the first 72 h after SAH, which consist of neuronal apoptosis [2 –4], blood–brain barrier (BBB) disruption [5,6], cerebral edema [7,8], and microcirculatory disturbance [9,10]. EBI has been considered as a main cause of poor outcome after SAH [1,11]. It is believed that the injury of endothelial cells, such as apoptosis or loss of intercellular junction, will lead to BBB disruption and brain edema after SAH [12,13].
The bone marrow mesenchymal stem cells (BMSCs) are promising therapeutics for some diseases, such as SAH [14,15], myocardial infarction, and renal I/R injury [16,17]. Previous results reveal that the protective role of BMSCs might be mainly through their paracrine effects, for example, secreting exosomes (Exo) [18,19]. Exo are nanovesicles derived from the multivesicular bodies in most cells [20]. Accumulating evidence indicates that, as the mediators, Exo facilitate intercellular communication through delivering their contents, such as proteins, mRNAs, and microRNAs (miRNAs) [19,21]. The miRNAs are small RNA molecules, which can regulate gene expression through degrading the corresponding mRNAs or suppressing their translation [22]. The miRNAs within Exo play pivotal roles in modulating the functions of target cells [23].
High-mobility group box 1 protein (HMGB1), a potent mediator of inflammation, is one of nonhistone DNA binding protein family member. It plays critical roles in establishment of autoimmunity or self-maintaining tissue damage [24]. In some diseases, HMGB1 is released from the injured cells to initiate the inflammation through its receptors such as advanced glycation end products or Toll-like receptors (TLRs) through producing chemokines or cytokines [25 –28]. Moreover, the hyperfunction of HMGB1 has been detected before the production of tumor necrosis factor-α (TNF-α) and interleukin (IL-1β) [25]. HMGB1 can also facilitate the activation of p53 and initiate the apoptotic cascades in endothelial cells and neurons in some brain diseases [29 –31]. It has been reported that some miRNAs, for example, miRNA129-5p, can suppress the activity of HMGB1 and play their beneficial roles in lung cancer [32].
Therefore, we hypothesized that the Exo derived from MSCs can alleviate the EBI after SAH through miRNA129-5p-HMGB1 pathway and provide the novel therapeutic approaches for several brain diseases.
Materials and Methods
The methods in this study were approved by the Animal Care and Use Committee at Sichuan University and with the Guidelines for the Use of Animals in Neuroscience Research by the Society for Neuroscience.
Exosomes preparation and identification
MSC culture
The animals (Sprague-Dawley rats, 4 weeks old) were anesthetized with 10% chloral hydrate and sacrificed with cervical dislocation method. The femur and tibia were harvested and rinsed with normal saline (0.9% NaCl). The marrow cavity of the bone was rinsed with Dulbecco's modified Eagle's medium/nutrient mixture F-12 (DMEM/F12) medium by using a 10 mL syringe. The single cell suspension was centrifuged at 1,000 r/min for 10 min. The cells were seeded (106/mL) into a dish and cultured into an incubator (37°C, 5% CO2). Sprague-Dawley rat BMSCs were cultured in DMEM/F12 supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin (Gibco-BRL, Grand Island, NY). After 24 h, the cells were rinsed using phosphate buffer saline (0.1 M phosphate-buffered saline [PBS]) to remove the unattached cells. After 3 days, the cells were digested using 0.25% trypsin while fusing for 80%, passaged 1:2 (inoculation density 1 × 105/cm2), and then passaged once every 3–5 days. The third-generation cells were applied for further experiments.
Exosome extraction (SBI Exo-Quick)
MSCs that reached 80% confluency were washed with PBS three times and then were cultured in DMEM/F12 supplemented with 1% penicillin–streptomycin for 48 h, which were harvested and centrifuged for 15 min (3,000 g, 4°C). The supernatant was centrifuged for 10 min (16,000 g, 4°C). MSCs-derived Exo were isolated using SBI Exo-Quick according to the manufacturer's protocol. The Exo-Quick reagent was added into the supernatant at 4°C overnights (sample volume:reagent volume = 5:1). The supernatant was aspirated after centrifuging (1,500 g, 30 min), the pellet was resuspended by using 0.1 M PBS.
Exosome identification
Nanosight particle size analysis
The Exo particle size and concentration were measured by using nanoparticle tracking analysis (NTA) with ZetaView PMX 110 (Particle Metrix, Meerbusch, Germany). NTA measurement parameters were analyzed at 11 positions. The ZetaView system was calibrated by applying 110 nm polystyrene particles. The temperature was maintained at 23°C–37°C in this study.
Electron microscopy
The Exo sample (20 μL) was dropped into a copper grid and kept on the copper mesh for 1 h, and then fixed using 2% acetic acid oxychloride followed by uranium aqueous solution for 10 min. The sample was observed under a 80 kV biotransmission electron microscope (JEM-100S; JEOL, Japan).
Western blot
The western blot assay was performed to detect the Exo marker protein, including positive markers, such as CD63, CD9, and TSG101 (Abcam, Cambridge, UK) and negative marker, for example, GM-130 (Novus Biologicals, Littleton, CO). The detailed method is described as follows.
Establishment of SAH rat model
The male Sprague-Dawley rats (300–350 g) were randomly divided into three groups: sham group (n = 25), SAH+PBS group (n = 42), and SAH+Exo group (n = 33).
The rat SAH model was established using endovascular perforation method as reported by others [13]. The animal was anesthetized using 4% isoflurane in a mixture of 60% medical air and 40% oxygen and maintained anesthesia with 2% isoflurane. The body temperature of rat was maintained at 37°C ± 0.5°C. A 4-0 monofilament nylon suture was introduced into internal carotid artery to perforate the bifurcation of anterior and middle cerebral artery. In the sham group, the nylon suture was withdrawn once the resistance was felt.
The rat limbs were fixed on the surgical table with tape, and a 6 mm incision was performed from the right groin to the distal end. The adipose tissue on the blood vessel was removed, and the right femoral vein was exposed. The mixture of 200 μg of MSCs-Exo and PBS to final volume of 200 μL (200 μL PBS in control group) was injected into the femoral vein within 5 min.
SAH grade assessment
The SAH severity was assessed using previous method [33]. The basal brain of rat was divided into six regions, and each was allocated a score (0–3): grade 0, no blood; grade 1, minimal blood in subarachnoid space; grade 2, moderate blood with recognizable arteries; grade 3, blood covering all the arteries. The total score was 0–18. The rats underwent SAH surgery with a score <7 were excluded in this study.
Brain water content
The brain sample was weighed immediately after removal (wet weight) and weighed again after drying in an oven at 105°C for 48 h (dry weight). The water content was calculated as [(wet weight − dry weight)/wet weight] × 100% [34,35].
Neurological function evaluation
Neurological functions were evaluated at 24 h after SAH by using an 18-point scoring system [36], which consisted of six tests evaluating spontaneous activity, movement of the limbs, forelimbs outstretching, climbing, body proprioception, and response to whisker stimulation. This test was performed by two independent observers.
BBB permeability assessment
BBB permeability was examined as previous reports [37]. The anesthetized animal was intravenously injected Evans blue dye (4%, 2.5 mL/kg) from the tail vein. At 1 h after injection, the rat was perfused by using 0.1 M PBS through the left ventricle. The brain sample was homogenized and centrifuged for 30 min (15,000 g, 4°C). The supernatant was mixed with equal amount of 50% trichloroacetic acid. The mixture was centrifuged for 30 min (15,000 g, 4°C). The absorbance of the brain sample was analyzed by using spectrophotometer at 615 nm (Thermo Fisher Scientific, Waltham, MA). The amount of Evans blue dye in the sample was calculated based on the standard curve.
Immunofluorescence staining
The anesthetized rat was perfused by using ice-cold 0.1 M PBS and 4% paraformaldehyde through the left ventricles (pH = 7.4). The brain was postfixed in the same fixative overnight. The sample was embedded in the paraffin, and the coronal sections (thickness = 10 μm) were obtained by using a cryostat (Leica Microsystems, Bannockburn, IL).
The sections were incubated with primary antibodies (1:200, Santa Cruz Biotechnology, CA) overnight at 4°C, and then treated with the corresponding fluorescence labeled second antibodies (1:400; Santa Cruz Biotechnology). The results were observed under Olympus BX51 microscope (Olympus Co., Tokyo, Japan) [38,39].
Western blot
The bilateral hippocampi were obtained by homogenizing with RIPA lysis buffer (Santa Cruz Biotechnology) and centrifuged (14,000 g, 4°C) for 30 min. The protein concentration of the supernatant was calculated by using a detergent compatible assay (DC protein assay; Bio-Rad Laboratories, Shanghai, China). The sample (50 μg) was loaded on an SDS-PAGE gel and electrophoresed, and then transferred onto the nitrocellulose membrane, which was incubated with primary antibody (1:200; Santa Cruz Biotechnology) overnight at 4°C. The membrane was incubated using corresponding secondary antibodies (1:1,000; Santa Cruz Biotechnology) at room temperature for 1 h. The immunoblots were observed with the ECL Plus chemiluminescence reagent kit (Amersham Biosciences, Arlington Heights, IL). The result was analyzed using Image J software (National Institutes of Health, Bethesda, MD) [40].
Quantitative real-time polymerase chain reaction
The RNA in hippocampus each group was extracted using miRNeasy micro kit (QIAGEN, GmbH, Germany). Reverse transcription of total RNA was performed using TaqMan MicroRNA Reverse Transcription kit (Applied Biosystem, CA). The total reaction system was 15 μL, and reaction conditions were as follows: 30 min at 16°C, 30 min at 42°C, and 5 min at 85°C. Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using TaqMan Universal PCR kit (Applied Biosystem). The reaction conditions were as follows: warm start for 10 min at 95°C, 15 s at 95°C, and 1 min at 60°C (40 cycles). Each sample was added to three wells. Expression levels of the miRNA were obtained using U6 as a reference gene. The expression of HMGB1 was determined by using β-actin as a reference gene. Relative quantification was performed by 2−ΔΔCt method [41]. Each experiment was performed three times to obtain the average value.
Data analysis
In this study, the data were expressed as mean ± standard deviation or median with interquartile range. The analyses were performed by using SPSS (version 24.0; SPSS, Inc., Chicago, IL). One-way analysis of variance followed by Tukey's multiple comparison test was applied to analyze the differences among the groups. For those non-normal distribution and unequal variance parameters, Kruskal–Wallis test was used to analyze the data. The neurological function and SAH grade were analyzed by using the Mann–Whitney U-test. Statistical significance was defined as P < 0.05.
Results
Isolation and characterization of Exo
In this study, Exo was isolated from the culture medium of BMSCs by multiple centrifugations. Then, we characterized the Exo using electron microscope, western blot, and NTA. The electron microscope results showed that the particle pellets were round-shaped vesicles with the membrane bounded (Fig. 1A). The western blot results showed that the isolated Exo contained the exosomal markers CD9 and CD63 (Fig. 1B). Furthermore, as shown in NTA results, the diameter of Exo was ∼110 nm (Fig. 1C). These results indicated that the Exo had been successfully isolated from the BMSCs in this study.
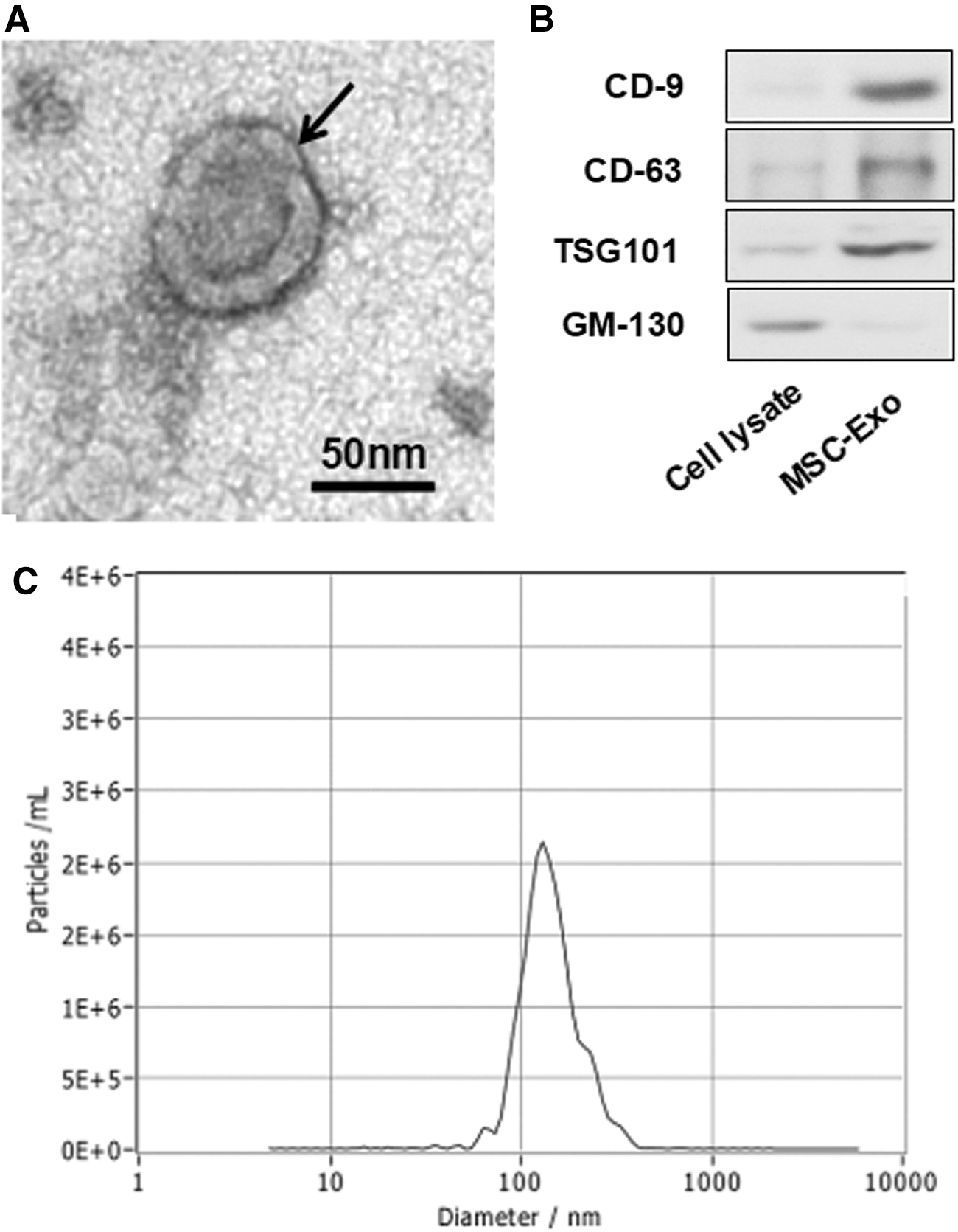
Isolation and characterization of Exo. The electron microscope results indicated that the particle pellets were round-shaped vesicles (arrow) with the membrane bounded
Effects of Exo on EBI after SAH
The mortality of animals in SAH+PBS group (40.47%, 17 rats died in total 42 animals, described as 17/42) was higher compared with that of sham group (0, 0/25, P < 0.05). After Exo treatment, the mortality after SAH was significantly reduced (24.24%, 8/33, P < 0.05 compared with that of SAH+PBS group) (Fig. 2A).
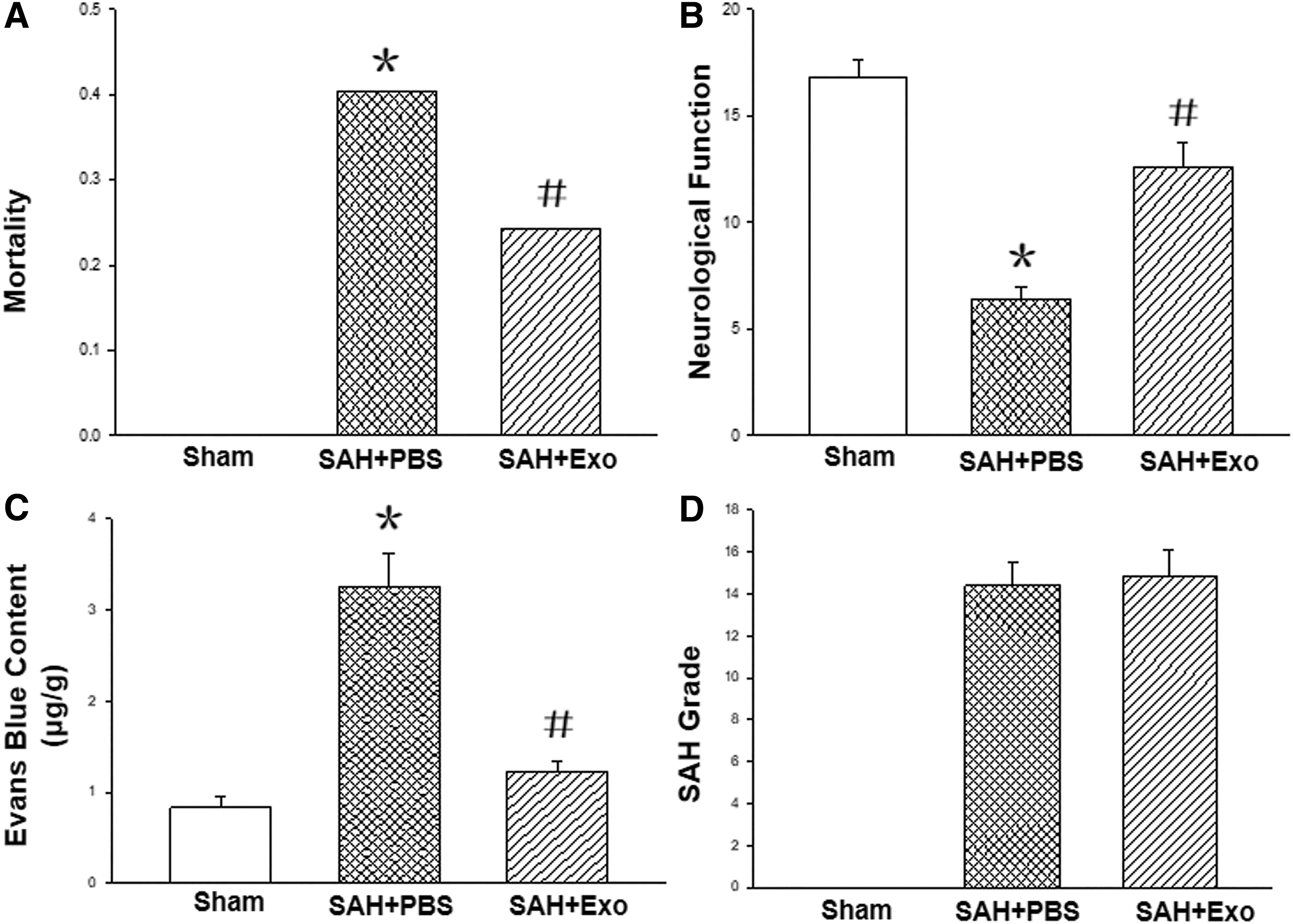
Effects of Exo on EBI after SAH. The mortality of animals in SAH+PBS group was higher compared with that of sham group. After Exo treatment, the mortality after SAH was significantly reduced
The neurological functions of the rats in SAH+PBS groups (6.40 ± 0.55) were impaired compared with those of sham group (16.80 ± 0.84, P < 0.05); after Exo application, the neurological deficits were significantly alleviated (12.60 ± 1.14, P < 0.05 compared with that of SAH+PBS group, n = 5 each group) (Fig. 2B).
The Evans blue in the brain of SAH+PBS group was markedly increased compared with that of sham group (3.26 ± 0.36 μg/g vs. 0.83 ± 0.12 μg/g, P < 0.05, n = 5 each group), which was reduced after Exo treatment (1.22 ± 0.13 μg/g, P < 0.05, n = 5) (Fig. 2C).
In addition, there was no blood in the subarachnoid space of the animals in sham group, and there was no statistical significance between the SAH grades of SAH+PBS (14.40 + 1.14) and SAH+Exo groups (14.80 + 1.30). These results indicated that the SAH severity was not affected by the Exo treatment; furthermore, the effects of Exo were independent of the SAH severity among the groups (Fig. 2D, n = 5).
The potential mechanism of Exo's effects
Involvement of miRNA129-5p in EBI
By using qRT-PCR assay, we found that the miRNA129-5p level in the hippocampus of sham animal was markedly elevated after Exo injection into hippocampus. At 24 h after SAH, the miRNA129-5p level was significantly reduced, which was increased after Exo treatment compared with that of SAH+PBS group (P < 0.05, n = 5). These results implied that the reduction of miRNA129-5p might be involved into the development of EBI after SAH, and replenishment of Exo might provide protective effects after SAH (Fig. 3B).
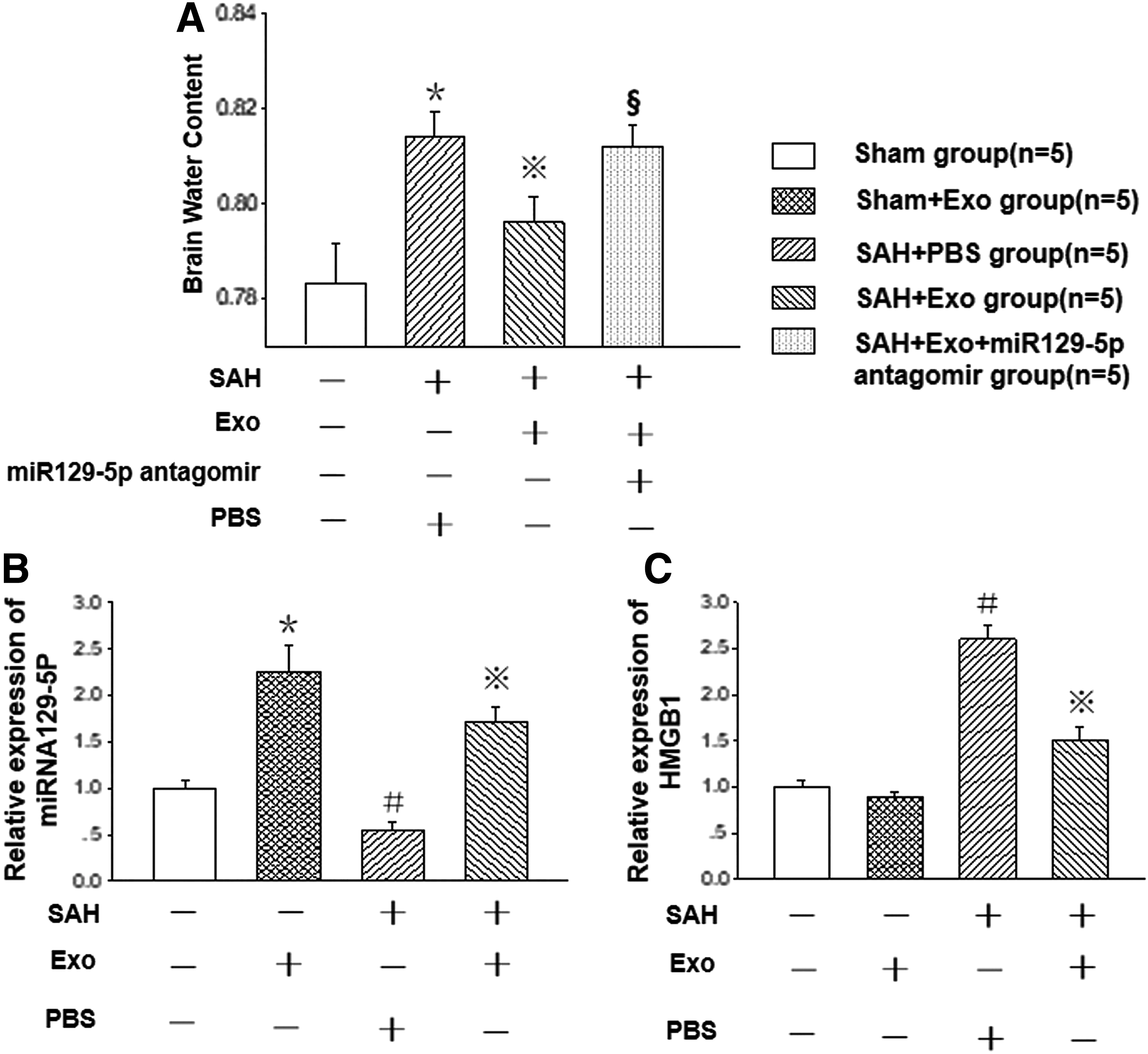
Involvement of miRNA129-5p in EBI. The miRNA129-5p level of sham animal was markedly elevated after Exo injection into hippocampus (P < 0.05). At 24 h after SAH, the miRNA129-5p level was reduced (P < 0.05), which was significantly increased compared with that of SAH+PBS group after Exo treatment
Protective role of miRNA129-5p in brain edema
The brain water content of animals in the SAH+PBS group (81.40% ± 0.55%) was significantly higher than that of sham group (78.32% ± 0.84%) (P < 0.05). After Exo treatment, the brain water content after SAH was markedly decreased (79.60% ± 0.55%). To confirm the role of miRNA129-5p in the alleviating BBB disruption and development of brain edema, we applied the miR129-5p antagomir (miR129-5p inhibitor; GenePharma Co. Shanghai, China). The results revealed that the brain water content was significantly increased again in the SAH+Exo group with the presence of miR129-5p antagomir (Fig. 3A, n = 5 each group).
Suppression of HMGB1-TLR4 pathway activity
The qRT-PCR assay results showed that the mRNA expression level of HMGB1 was significantly increased after SAH, which was markedly suppressed after the Exo treatment (Fig. 3C, n = 5).
To explore the protective mechanism of Exo's effects, the double immunofluorescence staining was performed. The results revealed that, after SAH, the expression levels of HMGB1 and TLR4 were simultaneously increased and colocalized in the endothelial cells (Fig. 4A1–B4), which were significantly reduced in the endothelial cells after Exo treatment (Fig. 4C1–C4). These results revealed that the HMGB1-TLR4 pathway was activated and involved into the brain injury after SAH, and Exo might play protective roles through suppressing this pathway activity.
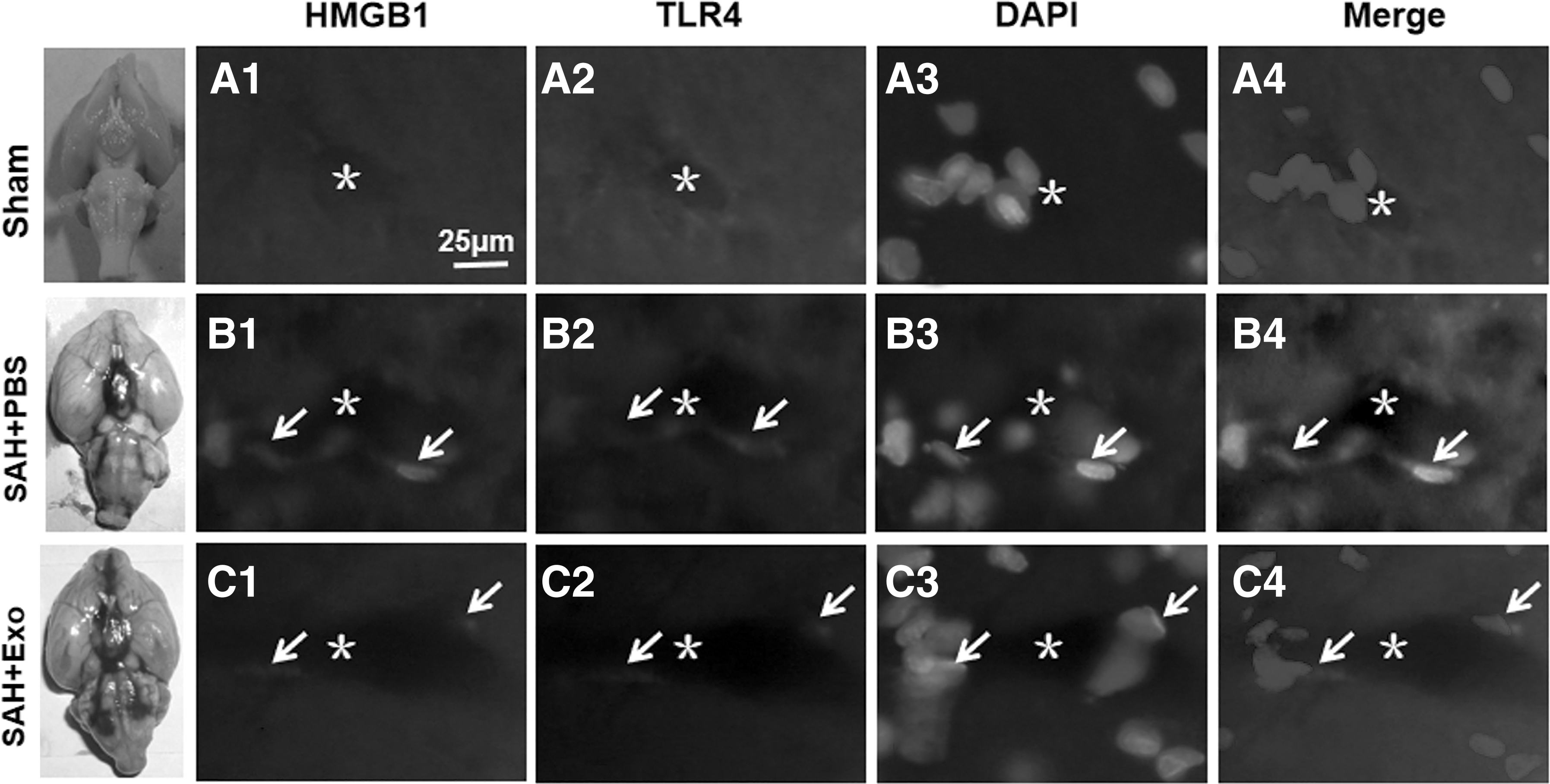
Exo suppressing HMGB1-TLR4 pathway activity. After SAH, the expression levels of HMGB1 and TLR4 were simultaneously increased and colocalized in the endothelial cells
In addition, the western blot results showed that, after SAH, the expression levels of HMGB1, TLR4, and the proinflammatory factor TNF-α, the proapoptosis factors phosphorylated-p53 were significantly elevated, which were markedly reduced after Exo treatment (Fig. 5A–D, n = 5).

Potential mechanism of Exo's beneficial effects. After SAH, the expression levels of HMGB1, TLR4, and the proinflammatory factor TNF-α, the proapoptosis factors phosphorylated-p53 were significantly elevated, which were markedly decreased after Exo treatment
Discussion
In this study, we explored the protective roles of Exo in attenuating EBI after SAH through its anti-inflammation and antiapoptotic effects through miRNA129-5p-HMGB1-TLR4 pathway.
After SAH, the increased intracranial pressure and decreased cerebral blood flow result in the severe global cerebral ischemia, which leads to the EBI, such as BBB disruption, brain edema, and cell death due to apoptosis, inflammation, and oxidative stress [1,2]. The previous studies revealed that the inflammation and apoptosis cascades in brain were involved in BBB disruption after SAH [2,42], which consequently leads to the leakage of liquid and the macromolecule from plasma into brain parenchyma, for example, IgG, which can be detected after combining with Evans blue dye [13]. In this study, the Evans blue content and brain water content were markedly elevated after SAH, which indicated that there was an obvious BBB disruption after SAH.
To explore the underlying mechanism of BBB injury after SAH, we detected the expression levels of proinflammatory and proapoptosis pathway. As a potent mediator of inflammation, HMGB1 and its downstream target TLR4 were simultaneously activated in the endothelial cells, and their expression levels were also elevated. It was reported that TLR4 could initiate MAPKs and NF-κB signal pathway and activate the inflammation and apoptosis cascades [43]. In this study, we found the expression levels of proapoptosis protein p53 and proinflammatory protein TNF-α were markedly enhanced after SAH, which might be partially induced by activated HMGB1-TLR4 pathway in the microvessels.
Some studies have inferred that the beneficial effects of MSCs mainly mediated by their paracrine action after transplantation. The Exo, a major form of paracrine components, has been confirmed as a critical mediator for intercellular communications under physiological or pathological conditions. The Exo transfer some specific components, such as proteins, RNAs, and even DNAs [44,45], which might determine the effects of Exo physiologically and pathologically.
miRNAs, 20–22 nucleotide-long noncoding RNAs, are specifically expressed in some cells, and can negatively regulate their target gene expression at the post-transcriptional level [46]. The miR-129-5p, a tumor suppressor [47], has been shown to ameliorate the neuroinflammation and blood–spinal cord barrier damage in the spinal ischemia-reperfusion injury [48]. In addition, miR-129-5p prevents NF-κB transduction and HMGB1-TLR4 inflammatory signaling [28,49]. In this study, qRT-PCR results showed that the level of miR-129-5p was significantly increased after Exo application in sham animal, which indicated that replenishment of Exo could effectively enhance the level of miR-129-5p in the rat brain. After SAH, the level of miR-129-5p in brain was decreased; its specific mechanism was still to be clarified. We speculate that it might be, at least partly, due to the release of miR-129-5p into CSF or plasma like other miRNAs [50,51]. Consequently, the level of its downstream target HMGB1 was significantly enhanced. Meanwhile, western blot results revealed that the inflammation reaction (TNF-α) and apoptotic cascades (p53) in endothelial cells were also activated, which participated in the development of BBB disruption. These results indicated that the dysfunction of miR-129-5p might be responsible for the EBI after SAH, which might be attenuated by replenishment of miR-129-5p. As far as we have known, it is the first time to investigate the role of miR-129-5p within MSCs-Exo in alleviation of EBI after SAH.
After injection of the Exo into hippocampus after SAH, the level of miR-129-5p, other than other miRs (data not shown), was found significantly enhanced. Indeed, MSC-derived Exo can serve as a vehicle to transfer some miRNA to distant recipient cells, altering the miRNA expression of the recipient cells. In this study, the increased expression level of miRNA129-5p after MSC-Exo treatment might, at least partly, originate from the injected MSC-Exo. In contrast, the increased miRNA129-5p might be newly produced by the adjacent tissue under the modulation of the components in the MSC-Exo, such as some RNAs or proteins.
Meanwhile, with the increased level of miR-129-5p, HMGB1, both in RNA and protein levels, was markedly reduced, and the expressions of TNF-α and p53 were also declined. To confirm these results, we applied the miR-129-5p antagomir (inhibitor) in evaluation of Exo's effect on the brain water content. The results showed that the protective role of Exo in alleviating brain edema was reversed in the presence of miR-129-5p antagomir. These results indicated that miR-129-5p could quench the activity of HMGB1-TLR4 signaling and attenuate the inflammation and apoptosis cascades in brain and might be critical beneficial component in Exo on alleviation of EBI after SAH.
In this study, the expression level of HMGB1 is not significantly changed in non-SAH with Exo injection group, as shown in Fig. 3B and C. Normally, HMGB1 is a highly conserved DNA-binding protein that functions as an intracellular regulator of nucleosome formation and gene transcription [52]; therefore, under physical condition, it is comparatively stable and cannot be easily modulated by other regulatory factors, for example, miR-129-5p. However, under pathological conditions, for example, SAH in this study, HMGB1 can be activated and translocate from the nucleus to the cytoplasm [25]. The activity of HMGB1 is enhanced and liable to be regulated by other factors due to its location (in the cytoplasm, even in the extracellular space other than in the nucleus). In this study, the expression level of HMGB1 is not significantly changed in non-SAH with Exo injection group, which might be due to its stability under physical condition. Meanwhile, its comparatively enhanced activity and cytoplasm location might account for its regulation by miR-129-5p after SAH.
Conclusions
In this study, we found that Exo played its protective roles in attenuation of EBI after SAH, which was mainly mediated by miR-129-5p in Exo through its antiapoptosis and anti-inflammation effects through suppressing the activity of HMGB1-TLR4 pathway.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (grant no. 11932014 and 31971239).