
Abstract
Urodeles and some fishes possess a remarkable capacity to regenerate their limbs/fins, a property that correlates with their additional ability to regenerate large areas of the brain and/or produce a variety of new neurons during adulthood. In contrast, neurogenesis in adult mammals is apparently restricted to two main regions, the subventricular zone of lateral ventricles and the subgranular zone of the hippocampus. There, astrocyte-like neural stem cells (NSCs) reside and derive into new neurons. Although it is becoming apparent that other brain regions carry out neurogenesis, in many cases, its functional significance is controversial, particularly, because very few putative NSCs capable of deriving into new neurons have been found. Hence, is renewal of certain neurons a requirement for a healthy brain? Are there specific physiological conditions that stimulate neurogenesis in a particular region? Does the complexity of the brain demand reduced neurogenesis? In this study, we review the production of new neurons in the vertebrate adult brain in the context of a possible functional relevance. In addition, we consider the intrinsic properties of potential cellular sources of new neurons, as well as the contribution of the milieu surrounding them to estimate the reparative capacity of the brain upon injury or a neurodegenerative condition. The conclusion of this review should bring into debate the potential and convenience of promoting neuronal regeneration in the adult human brain.
Introduction
The function of the brain is based on neuronal interactions occurring locally in specific regions or within defined network circuits between different regions. The input fibers make synapses with the cell body and/or branched processes (dendrites) originated from the nerve cell. Thus, disrupting a neuronal network should have dramatic functional consequences. The general neuronal composition of the brain includes two different neuron types that can be classified as follows: principal or projection neurons, those that send an axon to other regions, and the so-called intrinsic neurons or interneurons, involved in local processing (Table 1). Together with the incoming input fibers, local interneurons and projection neurons define the specific functional operations of each brain region [1]. Any replacement due to neuronal renewal should comply with the requirement of inserting into the circuit new elements without a major perturbation and restoring the input and output connections. Therefore, a functionally relevant neurogenic process should involve the birth of new neurons and, specially, their integration into the correct neuronal circuit. Actually, circuit integration might limit the impact of new neurons produced. Also, any artificial induction of neurogenesis should assume the existence of requirements for integration into a circuit.
The Major Neuronal Classes in the Adult Brain
In general, the brain is composed of two markedly different types of neurons: principal or projection neurons that establish communication between relatively distant regions, and intrinsic neurons or interneurons that regulate and synchronize the local network. These two types of neurons can be distinguished not only by the length and distribution of their axons but also by other morphological and physiological properties.
During a typical neurogenic process (eg, that occurring in the embryo; Fig. 1), new neurons are generated from neural stem cells (NSCs). In the embryo, their fate is determined during gastrulation and early neurulation such that they differentiate into specific neurons according to their position [2,3]. NSCs have the ability to self-renew and, upon differentiation, commonly produce transient neural progenitor cells (NPCs), some with high proliferative activity, which turn into immature neurons (neuroblasts). Within the embryo, terminal neuronal differentiation occurs as neuroblasts/immature neurons migrate and mature through receiving and sending short and long projections, creating a functional circuit. Due to intrinsic and extrinsic influences, the fate of the neural precursors derived from those early NSCs significantly change and mostly produce astrocytes at later developmental stages [4,5]; whether they retain neurogenic capacity in vivo is presently unclear. Nowadays, there is evidence that adult NSCs are generated during embryogenesis, mainly from slowly dividing NPCs that remain quiescent to participate in adult neurogenesis [6,7]. The rate of adult neurogenesis can be modulated by environmental or hormonal cues (see below).

Mechanisms to produce new neurons in the adult brain. Based on the cell of origin, adult neurogenesis can be classified as typical and atypical. The typical process (bottom) involves NSCs that proliferate and differentiate into neural progenitors/neuroblasts, which then become mature neurons. Alternatively, a non-neuronal differentiated cell, commonly astrocytes, can also be converted into morphological- and functional-like neurons by extrinsic inducers (eg, brain damage, EB transplantation, or angiogenic factors) or by intrinsic inducers, usually master proneural genes (eg, NeuroD, Ascl1, and Sox2), which establish a neuronal genetic network. In these two latter cases, dedifferentiation, transdifferentiation, and reprogramming are possible processes involved. Extrinsic inducers usually convert astrocytes into NSCs or neuroblasts (left), whereas intrinsic inducers can directly convert astrocytes into neuroblasts or neurons (right). Color images are available online.
Brain regeneration, in contrast to typical adult neurogenesis, should imply the production of neurons and other cell types associated (eg, astrocytes and oligodendroglia, microglia, and endothelial cells) after a traumatic injury, as well as, the correct structural organization, and connectivity, which usually depend on extracellular matrix, a cellular scaffold, or guiding molecules (Fig. 2). However, it is common that authors refer as brain regeneration to the high neurogenic activity observed in some urodeles and teleost fishes upon injury [8 –10]. Although, in this case, reconstruction of large regions is apparent, it has been observed, for example, that the regenerated pallium in axolotls has abnormalities in its architecture and new neurons failed to reconnect properly with long-distance targets [11]. Also, it is important to mention another regenerative process that implies mostly axon outgrowth (ie, axogenesis/axon regeneration), which is remarkably effective after a lesion in the peripheral nervous system, in contrast to the central nervous system (CNS) [12] (Fig. 2). On the other hand, recent evidence shows the birth of new neurons whose origin is not defined, but presumably involves differentiated cells instead of precursor cells and, thus, a dedifferentiation and/or a transdifferentiation process could be involved in the generation of new neurons (see details below); we denominate this process atypical neurogenesis (Fig. 1).
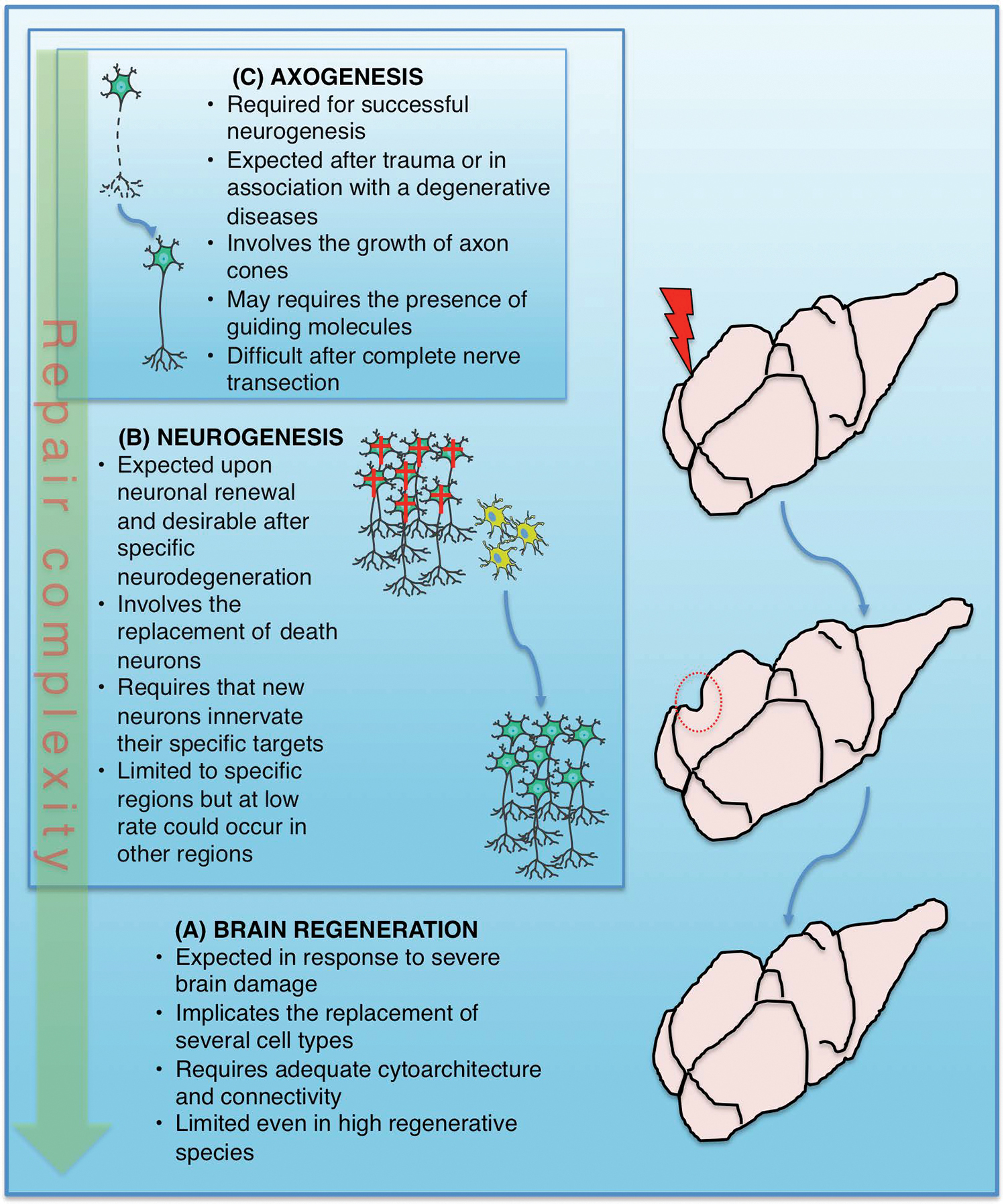
The extents of brain repair and maintenance.
Classical Neurogenic Zones in the Adult Mammalian Brain
Santiago Ramón y Cajal in his book “Estudios sobre la degeneración y regeneración del sistema nervioso” described the feasibility of axogenesis/axon regeneration and the lack of newly generated neurons in adulthood [13]. As many studies reinforced the network-wiring model of brain function, it is difficult to imagine the integration of new neurons into the adult brain circuits. This difficulty may be exemplified by the poor impact, particularly from the functional point of view, of early reports showing the generation of new neurons in the adult brain [14 –16]. A second wave of evidence in the last decade of the 20th century, exposing the formation of new neurons in the adult brain, was required to attract more interest in adult neurogenesis and, overall, question the significance of these observations [17,18].
Nowadays, two neurogenic regions have shown abundant production of new neurons in the brain of adult mammals, including humans [19,20], even though a recent report states that neurogenesis in the monkey and human hippocampus declines with age to undetectable levels [21]. Specific niches in the subventricular zone (SVZ) and subgranular zone (SGZ) of the anterior telencephalon and hippocampus, respectively, show dividing NSCs, whose progeny can be traced into new neurons. Those neurons born in the SVZ derive from NSCs that differentiate into neuroblasts, which migrate a long distance throughout the rostral migratory stream to reach the olfactory bulb (OB) and become new periglomerular or granule interneurons [22,23]. On the other hand, those born in the SGZ differentiate into intermediate neuroblasts that, during several days, migrate until reaching the granule cell layer of the dentate gyrus, where they become excitatory neurons that participate in the local circuitry [24]. It is apparent that neurogenesis in these two highly vascularized niches follows a similar differentiation process starting with NSCs (B or type I cells), which differentiate into transit-amplifying cells (C or type II cells) that then become intermediate nondividing neuroblast (A or type III cells). NSCs in these two regions mostly give rise to neurons in vivo, but in vitro also show the ability to produce astrocytes and oligodendrocytes [25]. The potential of SVZ NSCs to generate oligodendrocytes and astrocytes in vivo has been shown to depend on their location in the lateral ventricle [26] and upon injury [27,28].
Functional Significance of New Neurons in the Adult
In the main neurogenic regions, neurogenesis ablation has physiological consequences. In the hippocampus, newborn neurons correspond to granule cells, which are principal neurons that connect with the CA3 area. Therefore, hippocampal neurogenesis has a crucial role in long-term potentiation [29] and contributes to learning and memory processes [30]. Thus, when neurogenesis is blocked, behavioral deficiencies have been reported in tests such as the Morris water task, open field activity, and object/context mismatch [31]. Three hypotheses have been proposed by Aimone et al. [32] regarding the participation of new granule cells: (1) newborn neurons are the primary recipients of learning and, as a consequence, they help to distinguish highly similar events in composition and configuration; (2) the role of new neurons is encoding a temporal context to separate two temporally close events; and (3) new neurons are capable of encoding novel events, while old neurons encode past events, allowing memory resolution. It is relevant to mention that despite the significant neurogenesis observed in the hippocampus, only few neurons integrate into the circuits, while the apparent superfluous neurons produced die around the first weeks of their birth [24]. However, their proliferation and survival could be enhanced, for example, by exercise (eg, running) or exposure to enriched environments, respectively [33].
In the case of the OB, loss of new interneurons disrupts its electrophysiological properties (eg, reduction of miniature spontaneous inhibitory currents) through a decrease in the number of inhibitory synapses on the mitral cell lateral dendrites that, consequently, decrease synchronized neuronal activity [34]. The mitral cells are the OB projection (ie, principal) neurons; they establish dendro-dendritic synapses with granule neurons, an outcome of adult neurogenesis. Therefore, efficient interpretation of odors may require OB neurogenesis. The execution of the rodent sexual behavior is one example in which the olfactory systems, main OB (MOB) and accessory OB (AOB), are important and, thus, the role of adult neurogenesis could be evaluated [35 –38]. Accordingly, an increase in newborn neurons in the MOB is observed in female mice exposed to pheromones of dominant males [39]. Also, it has been reported that repeated paced mating (ie, sexual interaction regulated by the female at a frequency and intensity manner that enhance fertility) promotes the arrival of more new granule cells in the OB of adult male [40] and female rats [41]. Furthermore, newborn granule cells in the AOB appear essential for mating partner recognition in the female mouse [42] and offspring care [43]; indeed, disruption of OB neurogenesis by focal radiation of the SVZ in adult female mice alters social interactions with males, but not with females [44]. On the other hand, Kermen et al. [45] propose that olfactory learning consolidation, a process requiring protein synthesis, determines the survival of newborn neurons, which are helpful for long-term memory formation.
It is important to note that, because neurons with short axons are the main class of neurons generated in the adult brain, establishment of local connectivity seems to be the only requirement for integration to already established circuits. It is well known that inhibitory and excitatory interneurons, in addition to synchronize principal neurons, help to enhance the sensitivity to fluctuating stimuli and to refine information processing [1]. In the adult OB, new neurons remodel the circuit as old interneurons are replaced. This latter appreciation of neurogenesis supposes that interneurons suffer a functional wear up causing death and, therefore, a replacement is necessary to keep circuits working. In the OB, this replacement could respond to the continual renewal of sensory neurons of the olfactory epithelium, an outside CNS neurogenic region [46], in a process that might require the refreshment of olfactory circuits. However, several experiments have shown that OB neurogenesis ablation affects olfactory discrimination, but not the capacity of smelling [47]. In the hippocampus, neurogenesis ablation impairs spatial memory and contextual fear conditioning [29,48], while physiological induction of neurogenesis in this region (eg, interactive environments and exercise) increases learning and memory abilities. However, a question about the functional requirement of neurogenesis in this region remains opened since it apparently decreases in adult humans [21,49].
From the observations in the two main neurogenic areas, it is possible to consider that neurogenesis can become a necessary process for a specific circuit when its function requires a constant neuronal replacement or recruitment. Thus, can a functional demand under specific physiological conditions turn on neurogenesis in apparently non-neurogenic brain areas? In the hypothalamus, a brain region responsive to metabolic and hormonal activity, recent findings have demonstrated that new neurons integrate to the median eminence and arcuate nucleus during adulthood and this process seems to be influenced by the metabolic and nutritional status of the subject [50].
The new GABAergic interneurons that appear to be generated at slow rate in the cortex and striatum (see below) may provide plasticity to circuits, which seems a safer option than renewal of projection neurons (Table 1). Dopaminergic neurogenesis in the substantia nigra remains controversial because most researchers have not identified newborn dopaminergic neurons using BrdU labeling. However, it has been demonstrated that dopaminergic neurons derived from human embryonic stem cells (ESCs) and transplanted into the mouse substantia nigra have the ability to project to the striatum [51] showing the feasibility of new long-projection neurons to reach their target in the adult mammalian brain.
Neurogenic Potential of the Mammalian Adult Brain
New neurons from NSCs: typical neurogenesis
Regarding the possibility that other regions of the adult mammalian brain hold neurogenic potential, many studies have focused in searching NSCs and their progeny by combining BrdU labeling and the detection of immature and mature neuronal markers. Following a typical neurogenic process (Fig. 1), dividing cells (eg, BrdU+), which could be putative NSCs, and their neuronal progeny identified by NeuN, Tubb3, HuD, and MAP2, among other neuronal markers, have been found in the cortex, striatum, amygdala, piriform cortex, hypothalamus, substantia nigra, and vagus nucleus (reviewed in Chaker et al. [52]). In 2014, Conway and Schaffer [53] found broadly distributed EphB4 receptor (EphB4R)-immunoreactive cells, some of which co-expressed the NSC marker, Sox2. The administration of its ligand ephrin B2, normally produced by hippocampal astrocytes in young rats, increased neurogenesis in the hippocampus of old subjects, and also promoted the emergence of doublecortin-positive (Dcx+) cells in the striatum and the cortex, an indication that resident quiescent NSCs in these regions can initiate neurogenesis [53]; the functional consequences after this treatment remain to be demonstrated.
Specifically, new neurons have been found in the amygdala, piriform cortex, and temporal cortex of non-human primates [54]; the amygdala and hypothalamus of female prairie voles [55] and mice [50]; and the vagus nucleus of rats [56]. In some instances, the specific neuronal type generated in the adult has been identified. For example, in the cortex and striatum of rats, the new neurons express GABAergic markers (GABA/GAD-67, calbindin, and calretinin) [57], and in the caudate nucleus of rabbits, the new neurons are calretinin positive [58]. Similarly, in the striatum of humans, new GABAergic interneurons appear to be generated from in situ NSCs [59]. However, in all cases, the new neurons identified never correspond to projection neurons and rather seem to be interneurons (Table 1). An exception is a report from Zhao et al. in which they reported the incorporation of new dopaminergic neurons to the medial-rostral part of mice substantia nigra by BrdU and [3H] thymidine incorporation [60]. However, other research groups could not replicate these results [61,62]. Nonetheless, another report suggests also the generation new dopaminergic neurons originated from periaqueductal precursors in mice [63]. Despite these observations and, in contrast with the SVZ- and SGZ-associated neurogenesis, the number of dividing NSCs and/or new neurons is relatively very low in these other regions under normal conditions.
On the other hand, it has been observed that injury stimulates proliferation of putative NSCs and the generation of new neurons in these potential neurogenic regions. For example, newborn neurons generated upon stroke and excitotoxic injury in the striatum express markers of striatal medium-sized spiny neurons, but they represent only about 0.2% of lost neurons; the origin of these neurons appears to be the SVZ, suggesting that this niche holds NSCs with the potential to differentiate into striatal neurons [64,65]. Thus, under injury, it appears that NSCs are recruited from neighboring niches. The emergence of new neurons in the rodent cortex upon damage, which apparently are able to establish corticothalamic connections [66], has also been reported. In the mouse substantia nigra, the number of proliferating cells increases and new neurons emerge upon 6-OHDA administration, a highly specific neurotoxin that targets catecholaminergic neurons [60], but a similar study does not support this conclusion [62]. Also, MPTP-induced dopaminergic neuron death awakes activity in another recently described putative neurogenic niche, the periaqueductal region of the midbrain in mice, which appears to repair the damaged substantia nigra by incorporation of new dopaminergic neurons (TH+, DAT+) [63,67]. Thus, at best, it appears that even under damage, the awakening of neurogenesis in most of the brain is an inefficient process.
The above observations do not exclude the possibility that residing cells capable of generating new neurons exist in different brain regions, but none of the conditions tested have been able to awake them to initiate a neurogenic process. If this were the case, the putative neurogenic region should hold an environment that, at least, is permissive for neurogenesis. Thus, we recently developed a strategy to assess the neurogenic potential of a specific adult brain region by determining its capacity to support neurogenesis of transplanted naive precursor cells with demonstrated neurogenic potential [68]. By transplanting cells derived from ESCs in adult rats (ie, embryoid body cells), we have shown that within the SVZ-rostral migratory stream, 50% of grafted cells reach a neuronal fate after 3 weeks, whereas in the striatum, a poorly neurogenic region, astrocytic differentiation is a favored fate of transplanted cells. In agreement with the increase in neurogenic potential due to brain damage, new neurons derive from transplanted cells in the striatum after a stroke.
Interestingly, the intact substantia nigra pars compacta behaves as a strong promoter of neurogenesis of transplanted cells in our assays since up to 45% of total grafted cells were NeuN+ [69]. However, despite the efficient generation of neurons from transplanted cells, none showed specific markers of the host intact region [70], but evidence of dopaminergic differentiation of transplanted ESC-derived cells could be detected after induced degeneration of substantia nigra dopaminergic neurons [70]. These observations suggest that not only promoters of neurogenesis could emerge upon brain damage but also factors that favor specific differentiation. Therefore, “silent neurogenic niches” awaiting to find the conditions to activate latent NSCs may exist in the adult brain.
New neurons from non-NSCs: atypical neurogenesis
Evidence suggests a neurogenic potential of a variety of cells in the adult brain (astrocytes, oligodendrocytes, and glial precursors) when they are stimulated (eg, by a factor infusion or injury) or transplanted to a known neurogenic environment (Fig. 1). For example, NG2+ progenitor cells were identified in the cortex [57] and the substantia nigra [61,71], reinforcing the existence of locally residing oligodendrocyte precursors. Although it was believed that NG2+ progenitor cells only produce new oligodendrocytes in vivo, they can be reprogrammed into neurons in the injured cortex [72], display a multipotent phenotype in vitro [73], and produce new neurons when transplanted to a neurogenic region such as the hippocampus [61,74]. In the case of differentiated cells such as astrocytes, their conversion into neurons has been accomplished by the forced expression of proneural factors (eg, NeuroD1, Ascl1, Brn2a, and Myt1l) and dopaminergic fate determinants (eg, Lmx1a/b, Foxa2, Otx2, and Nurr1) in vitro and in vivo [75 –77] (Fig. 1, intrinsic inducer). Moreover, parenchymal astrocytes have been shown to gain neurogenic properties in response to damage in the striatum or in the cortex [78,79] and the spinal cord [80] (Fig. 1, extrinsic inducer). Interestingly, Notch1 signaling reduced upon stroke or experimentally reduced in uninjured animals promotes the activation of quiescent ependymal cells [81] and a latent neurogenic program in striatal astrocytes [79]. In this latter case, astrocytes gain a multilineage potential, transiting from Gfap+/S100+ cells (glial type) to Ascl1+/Dcx+ neuroblasts in 3 weeks, which later derive into NeuN+ mature neurons. Similarly, we have reported that 20%–30% of Gfap+/S100b+ cells in the substantia nigra acquire neuroblast markers (Psa-Ncam/Dcx) 15 days after embryoid body cell transplantation, and some of them express Tubb3, a neuronal marker, and show neuronal morphology on day 30 [82].
The above observations indicate that there are cells that restraint their intrinsic neurogenic potential due to the lack of extrinsic neurogenic-promoting factors. However, hardly this can be considered the general situation as many characteristics of differentiated cells are imposed by intrinsic restrictions on the genome. In particular, reactive astrocytes, which have been considered a potential source of new neurons, generally do not differentiate into neurons when they emerge in damaged brain regions [83]. Previously, this was thought to be an irreversible condition, but, presently, the limits of these epigenetic restrictions can be evaluated by the expression of “master genes,” mostly encoding transcription factors, which can increase genome plasticity or force the conversion into specific neuron types even from lineage-distant cell types such as fibroblasts and brain pericytes (reviewed in An et al. [84]; Fig. 1). Reactive astrocytes have been converted into neurons with a variety proneural transcription factors in vitro and in vivo [83]; for example, reactive astrocytes in the injured cortex of wild-type or 5xFAD transgenic mice, a model of Alzheimer disease, can be reprogrammed into neurons by the expression of NeuroD1 [72]. Also, it has been shown that the in vivo expression of Sox2 in resident astrocytes (ie, driven by the Gfap promoter) promotes neuronal differentiation [85]. Similarly, the combination of Ascl1, Brn2a, and Myt1l in astrocytes can generate neurons in vitro and in vivo [75], and Ascl1, NeuroD1, Lmx1a, and miR218 reprogram human and mouse astrocytes into induced dopaminergic neurons [76].
Nonetheless, despite the remarkable conversion of astrocytes into neurons induced by extrinsic and intrinsic factors, this phenomenon occurs at a relatively low frequency. Interestingly, metabolic and oxidative stress are generated during the phenotypic conversion of reactive astrocytes into neurons induced by transcription factors such as Ascl1 or Neurog2, which when prevented with antioxidants or genes such as Bcl2, neuronal production is significantly increased [86]. These data support the actual possibility of changing the fate of restricted cells by genetic manipulations and/or by the addition of factors that alter metabolic and redox homeostasis. However, although these experimental observations may be useful for the development of therapeutic procedures, in no way does it imply that these processes occur during normal brain function or upon repair.
Evolutionary Perspective of Adult Neurogenesis (Renewal vs. Reparative Neurogenesis)
Neurogenesis has also been observed in non-mammalian adult vertebrates. Actually, the discovery of neurogenesis in songbirds such as zebra finch and canary was inspiring for deep studies in mammalian neurogenesis. The neurogenesis in these birds is seasonal and related to song learning. During the reproductive season, new projection neurons are recruited and, as a consequence, the song control nucleus increases in size in a testosterone-dependent manner. Precursor cells reside in the ventricular zone and newborn neurons migrate through radial glia fibers to reach the high vocal center and integrate into functional neuronal circuits [87,88]. Seasons also appear to modulate hippocampal and SVZ-OB neurogenesis in mammals [89], but the significant change in size of brain of shrews in association with seasons has, however, a minimal, if any, contribution of neurogenesis and, rather, this is a process independent of cell proliferation or death [90].
More recently, pronounced and widespread neurogenesis has been observed in the adult brain of zebrafish, a vertebrate with a high neurogenic capacity. Active neurogenesis is observed in the OB, telencephalon, thalamus, epithalamus, preoptic region, hypothalamus, tectum, rhombencephalon, spinal cord, and cerebellum [91,92]. Its 16 progenitor niches are distributed along the rostrocaudal axis and contain putative NSCs, among them radial glia, which give rise to diverse types of neurons [93]. In another teleost fish, Apteronotus leptorhynchus, a lesion in the cerebellum induces cell proliferation and regeneration of granule neurons [94].
Urodeles and certain lizards, which normally show a restricted proliferation in the telencephalon and the most rostral diencephalon, generate new dopaminergic neurons in the adult newt after injury [95]. Reptiles, in general, add new cells to their telencephalon, while, in particular, some lizards show a broad regeneration of cerebral cortex upon injury, apparently indistinguishable from that of an undamaged brain [8]. However, a recent study performed in adult axolotls has pointed out that brain regeneration after hemisphere resection is an incomplete process and, despite all types of neurons being regenerated, the process fails in recovering the normal tissue architecture and the reconnection of long distance axons [11]. Although authors did not evaluate a behavioral recovery in the regenerated subjects, these results suggest that brain regeneration in highly regenerative species is a limited process (Fig. 2).
From a phylogenetic point of view, regeneration evolution appears to be a metazoan feature that once gained can only be lost for reasons that remain to be elucidated. Nevertheless, a recent report suggests that some species of Nemertea (unsegmented worms), contrary to their ancestor, gained the ability to regenerate their head, including the brain [96]. Commonly, species with the ability to regenerate a part of the body (eg, limb and tail) are also able to regenerate other organs with increased efficiency. Therefore, it is expected to find that urodeles, certain reptiles, and fishes also show increased neurogenesis in their brains and spinal cord during regeneration [97 –99]. Regeneration is a process that requires extensive cell proliferation and differentiation and, thus, it might originate, at least in part, from the mechanisms that control proliferation, differentiation, and maintenance of stem and progenitor cells. Supporting this notion, the gene encoding p19, a cell cycle inhibitor, is missing within the Ink4a locus in urodeles [100]. In the brain, this increased capacity for proliferation and differentiation might be the basis of the apparent added renewal rate of certain neurons in these species, although for repair after injury, the inflammatory response likely has an additional profound influence in the efficiency of regeneration [101].
Despite the similarities, there are distinguishable features between the mammal, fish, and amphibian adult brain that may be a hallmark in regeneration capacity. For example, radial glial cells, the embryonic NSCs, persist into adulthood as ependymoglia in the case of adult fish and amphibians [95], whereas, in mammals, neuroepithelial cells transit into radial glia cells, and finally become free astrocytes in adults [102]. In fact, the most abundant glial cells in adult mammals are parenchymal astrocytes. Another interesting difference is the distribution of the ventricular system. In mammals and other vertebrates, the main neurogenic niches in the adult brain are associated with the ventricular system where astrocyte-like cells act as precursor cells. In contrast, the ventricular system of zebrafish crosses above the telencephalon displaying a T-shaped array from a coronal view [103]. This organization could contribute to spread NSCs and the neurogenic process itself, while in the mammalian brain, the ventricular system wraps up the neurogenic niches. Moreover, a recent study showed that newts share the same neurogenic niches found in mammals, but after ablation within a non-neurogenic region, the specific neuronal type damaged can regenerate trough the activation of ependymoglia cells, a cell type apparently absent in mammals.
The observations described above support the idea that there are quiescent areas in the brain of these species that could be activated to generate new neurons under certain physiological or pathological conditions [95]. Interestingly, parenchymal astrocytes of mammals have the capacity to proliferate and re-express proteins characteristic of radial glia and NSCs, such as vimentin, nestin, and Sox2, during the astroglial reaction occurring upon injury [83]. Reactive astrocytes, in contrast with those derived from the intact brain, acquire the capacity to form neurospheres (ie, aggregates composed of NSCs and their progeny), and although it can be converted into neurons by proneural transcription factors as mentioned above, only under particular conditions in vivo (eg, hypoxia) are they able to generate neurons (reviewed in [104]). In this context, there is a possibility that parenchymal astrocytes directly derived from radial glia cells maintain a memory and behave as precursor cells upon injury or in the presence of signals emerging under certain physiological conditions. The recent reports referred above, showing that parenchymal astrocytes acquire a neuronal identity, support this notion [78,79]. Therefore, it is appealing to think that differentiated glial cells around an injury site can gain multilineage potential and be the source of new neurons, in analogy with the ependymoglia in urodele amphibians. Despite the above, it is interesting that in species evolutionarily distant from mammals, such as the adult crayfish (Procambarus clarkii), it seems that the anterior proliferation center, a hematopoietic organ close to the brain, provides NSCs to sustain neurogenesis [105].
In contrast to jawless vertebrates [106], another feature against nervous system regeneration in mammals is the presence of myelin, an insulating layer around nerves, which is inhibitory for axonal regeneration. Furthermore, the glial scar, composed of extracellular matrix and proteoglycans, which forms after injury, constitutes a barrier for axogenesis/axon regeneration and neurogenesis [107], while in fish and lizard, CNS axons regenerate even in the presence of a glial scar [97].
The Potential and Convenience of Promoting Regeneration in the Human Brain
Given the functional relevance of neurogenesis in the OB and the hippocampus of small mammals, many studies in humans have been focused in the same brain structures and found evidence of its occurrence [19,20]. In particular, consistent data support neurogenesis in early human postnatal life, but debatable data have left inconclusive whether new neurons continue to be generated during adulthood [108]. Thus, presently, there is no definitive evidence indicating that a specific area of the human brain demands neuron renewal for a proper neuronal circuit function. However, if it occurs, long-term information should not be kept within these renewable neurons, such that their incorporation into a circuit does not represent a challenge for brain functions. In addition, incorporation of long axon projection neurons would require signals that correctly guide axon growth. In this regard, it is interesting that human ESC-derived dopaminergic neurons transplanted to the rodent substantia nigra have the ability to project to the striatum and other specific target regions [51], showing the feasibility that new long-projection neurons reached their target in the mammalian adult brain. Although it is unlikely the cues that guide the formation of primary circuits during development are present in the adult brain, a scaffold might remain after neurodegeneration and used by new neurons to re-establish the precise projections.
Despite the present controversial appreciation of neurogenesis in the adult human brain, it is worth thinking that new neurons could emerge after a lesion or in association with a specific physiological condition. However, constraints to neurogenesis in the adult brain must exist as cells with a proliferative ability and regenerative properties, like NSCs, oligodendrocyte precursor cells, and astrocytes, are potential targets of oncogenic mutations that would produce their malignant transformation and originate a tumor [109]. Actually, the tight regulation of proliferation through tumor suppressor genes is a likely mechanism that prevents hyperproliferative disorders such as cancer in long-lived species [110]. Accordingly, telomerase activity as well as other positive regulators of cell proliferation are detectable in regenerative tissues such as the male germline and epithelial and hematopoietic tissues, which are also the origin of most human cancers [111]. Therefore, it can be anticipated that the use of regeneration as a treatment for degenerative disorders through the activation of proliferation would increase cancer risk. An extreme example of this possibility in the brain is a report showing the apparent dedifferentiation and transformation into gliomas of mature neurons and astrocytes after downregulating suppressor genes [112,113]. Considering this, it seems safer to generate new neurons by a transdifferentiation process in which cells are converted into neurons without an excessive proliferation stage [114].
Adult humans, as other mammals, have a limited capacity to regenerate and processes of wound repair commonly finish with scar formation and tissue dysfunction [115]. In contrast with regenerating animals that have the faculty to access to embryonic programs, to recover damaged or lost tissue without signs of a scar [116], regeneration capacity in mammals appears influenced by the highly developed immune system that plays a vital role in the switch from regeneration to scar formation [117]. Recently, it has been reported that a microRNA, miR-200a, contributes to define the gene expression pattern after injury in the axolotl. Interestingly, the inhibition of miR-200a during spinal cord regeneration in this animal induces the pairing of c-Fos to c-Jun instead to JunB, as occurs in humans, promoting a reactive gliosis and scar formation [98]. Therefore, reducing scar formation could increase the regeneration capacity of the mammalian CNS.
The high level of complexity of neuronal circuits in the human brain should also be considered a relevant factor that could have limited adult neurogenesis during human evolution. Fortunately, the human brain has many other strategies to contend with injury. These other plastic processes, alternative to neurogenesis, include synaptogenesis and dendritogenesis, as well as reconnection of local areas through rebuilding of axonal contacts. This phenomenon, known as compensatory plasticity, has been reported in blind people where visual cortex responds to auditory and tactile stimulations [118] or in people who have lost a limb [119]. Even though these strategies do not seem enough to compensate lost neurons in neuropathologies, it is also true that regeneration of projection neurons is a complex issue regarding their vast connections (around 6000 synapses per neuron) and distance of targets [1]. In conclusion, although possible, it seems very challenging to generate specific new neurons de novo that have the capacity to integrate correctly into the preexisting and complex neural networks of the adult brain. On the other hand, it seems tempting to search deeper into the relevance of interneuron neurogenesis upon circuitry function and into their contribution to prevent or ameliorate neurodegenerative disorders.
Footnotes
Acknowledgments
D.M.A. is a doctoral student of the Programa de Doctorado en Ciencias Biomédicas, Universidad Nacional Autónoma de México (UNAM), and received the fellowship 355036 from CONACyT. We are grateful with Dr. Jennifer R. Morgan for her comments and careful reading of the article.
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
This work was supported by CONACyT grants CB131031 and FOINS1723.