
Abstract
Mesenchymal stromal cells (MSCs) are attractive candidates for immunomodulatory cell therapy. However, it remains unknown how far therapeutic efficacy and potency are dependent on the dosage and activity of the MSCs. We previously observed that infusion of MSCs leads to rapid and transient changes in cytokine expression in blood, lung, and liver. In the present study, increasing doses of syngeneic adipose tissue-derived MSCs were infused in healthy mice and systemic changes in G-CSF, IL6, IL-10, and CXCL5 were detected 2 h after administration of 3 × 105 MSCs per animal, but not at lower doses. In lung and liver tissue, dose-dependent effects of MSCs on cytokine mRNA expression levels were detected from doses as low as 3 × 103 MSCs. Infusion of secretome-deficient or IFNγ-activated MSCs in healthy mice had similar effects on systemic cytokine levels as control MSCs, suggesting that in vivo at least the initial systemic effect of MSC administration is independent of the level of activity of MSCs, but depends on the response of host cells to MSCs. The results of this study reveal a rapid dose-dependent effect of MSCs and stress the important role of host cells in MSC treatment. This knowledge contributes to the design of rational MSC trials and to the quest for clinical efficacy of MSC therapy.
Introduction
The use of mesenchymal stromal cells (MSCs) has gathered significant momentum due to recent studies demonstrating their potential to treat diseases currently without effective treatment, as well as an alternative for therapies with severe side effects [1 –3]. MSCs have demonstrated promising results in the treatment of (experimental) diseases associated with an aberrant immune system, including Graft vs. Host disease, Crohn's disease, systemic lupus, rheumatoid arthritis, and in immunosuppressed organ transplant recipients [4 –10].
MSCs have been proposed to be capable of a broad range of immunomodulatory and regenerative effects, including the production of trophic factors that may allow them to induce regenerative processes at the sites of injury by stimulation of resident progenitor cells [11 –13]. MSCs may furthermore influence nearby cells through direct contact, or through transfer of organelles, enzymes, and vesicles [14 –17]. In addition, MSCs have the capacity to control immune responses through metabolic regulation, such as by controlling L-tryptophan and adenosine levels through indolamine 2,3-dioxygenase (IDO) and ecto-nucleotidase activity [18 –20]. Several of the immunomodulatory mechanisms are potentiated upon challenge of MSCs by inflammatory stimuli such as interferon gamma (IFNγ), which upregulates the expression of anti-inflammatory molecules by MSCs, including IDO, program death ligand 1, and transforming growth factor beta (TGFβ). In vitro assays demonstrate that pretreatment of MSC with IFNγ enhances their inhibitory effect on T cell proliferation and plasmablast differentiation of B cells [21].
An increasing number of studies, however, reveal that infused MSCs have a short lifespan and suggest that at least some of the effects of MSCs upon infusion are mediated through host cells [22 –24]. Previous studies demonstrated that changes in serum cytokine and chemokine levels, including increases in IL-6 and G-CSF, are detectable as early as 2 h after MSC infusion [25] and it is likely that host cells play a role in these fast responses. By tracking human cord blood-derived MSCs after intravenous infusion, we showed that MSCs accumulate in the lungs immediately after infusion and that after 24 h MSCs are phagocytosed by host monocytes.
In vitro experiments further demonstrated that monocytes shift from a CD14++CD16− classical phenotype toward a regulatory CD14++CD16+CD206+ phenotype after phagocytosis of MSCs [22,26]. These monocytes subsequently relocate from the lung through the bloodstream to other sites, thereby redistributing the effect of MSCs [22]. These findings suggest that MSCs are rather passive players in their immunoregulatory effect. Indeed, inactivated MSCs that lost the capacity to secrete cytokines and growth factors induce the same cytokine responses in septic mice as living MSCs [27], indicating that the effect of MSCs is mediated through recognition of MSCs by host cells [27,28].
While in recent years more has been learned about the physiological effects of MSC administration, and the implementation of MSC therapy is still hampered by insufficient understanding of the fundamental mechanisms of action. Despite numerous studies with MSCs in preclinical models and clinical trials, it is not clear whether MSC therapy acts through a regular dose/response pattern. Due to the intrinsic heterogeneity of MSCs and their complex mechanisms of action, which involve numerous intricate inter- and intracellular pathways, a clear dose/response may be blurred.
Although it is well established that pretreatment of MSCs with inflammatory factors can enhance their inhibiting effects on T and B cell proliferation and cytokine and immunoglobulin production using in vitro assays [25], there is limited evidence that pretreated MSCs have improved therapeutic effects in (pre)clinical models. Monocytes have been indicated as key mediators of the effect of infused MSCs but it is unknown whether pretreated MSCs have an enhanced effect on monocyte polarization.
The present study answers some of the fundamental questions on the association between MSC dosage and their immunomodulatory potency, the timing of onset of MSC effects and whether pretreatment of MSCs will enhance their therapeutic efficacy.
Materials and Methods
Isolation and culture of human MSCs for in vitro experiments
Human MSCs were isolated from subcutaneous adipose tissue, collected after written informed consent, as approved by the Medical Ethics Committee of the Erasmus University Medical Center, Rotterdam (protocol No. MEC-2006-190). Adipose tissue was surgically removed from the abdominal incision of healthy kidney donors. MSCs were isolated as described previously [29]. In short, adipose tissue was mechanically disrupted, washed with phosphate-buffered saline (PBS), and digested enzymatically with 0.5 mg/mL collagenase type IV (Life Technologies, Paisley, United Kingdom) in RPMI 1640 Medium with GlutaMAX (Life Technologies) for 30 min at 37°C under continuous shaking. The stromal vascular fraction was separated by centrifugation and resuspended in Minimum Essential Medium Eagle Alpha (MEM-α; Sigma-Aldrich, St. Louis, MO) containing 2 mM
MSCs were cultured expanded in MEM-α supplemented with 2 mM L-glutamine, P/S, and 15% fetal bovine serum (FBS; Lonza) and kept at 37°C, 5% CO2, and 20% O2. The cells showed a spindle-shaped morphology and expressed the MSC markers CD13, CD73, CD90, and CD105 and were negative for CD31 and CD45. MSCs were passaged at ∼80% confluency using 0.05% trypsin-EDTA (Life Technologies, Bleiswijk, the Netherlands). Cells were cryopreserved and thawed and cultured before experiments. Human MSCs used in experiments were between passage 2 and 8.
Isolation and culture of mouse MSCs for in vivo experiments
Mouse adipose-derived MSCs were isolated from omental adipose tissue of healthy male C57BL/6 mice and cultured as human MSCs as described above and previously [25]. Adipose tissue of five animals was pooled for the generation of a large batch of MSCs, which was aliquoted and frozen until use. Mouse MSCs showed a typical spindle-shaped morphology and expressed CD44 and Sca-1 and were negative for CD11b and CD45. The cells were frozen at passage 1 in 10% DMSO at −150°C until use. Mouse MSCs were thawed in MEM-α supplemented with 2 mM L-glutamine, P/S, and 15% FBS and expanded in 175-cm2culture flasks. Mouse MSCs between passages 2 and 6 were used for experiments.
Inactivation of MSCs
MSCs were inactivated by 30 min incubation at 50°C in a temperature-regulated water bath as described before[27]. MSCs were suspended in PBS, contained in parafilm sealed tubes throughout the duration of heat inactivation. Heat-inactivated MSCs (HI-MSCs) were washed and used immediately for downstream experiments or resuspended in MEM-α supplemented with 2 mM
Pretreatment of MSCs
MSCs were expanded and treated for 4 days with 50 ng/mL of IFNγ (Life Technologies). Immediately before injection, MSCs were washed with PBS and detached by incubation with 0.05% trypsin-EDTA. MSCs were subsequently suspended in PBS and administered immediately as described below.
In vivo experiments
Eight-week-old healthy female C57BL/6 mice were purchased from Charles River (Lyon, France). The mice had free access to food and water and housed in a facility with a 12h light cycle. All animal studies were approved by the national committee on animal care and experimentation. Syngeneic mouse MSCs were used in this series of studies to avoid xenogeneic and allogeneic responses. C57BL/6 adipose tissue-derived MSCs were trypsinized, washed two times in PBS, and resuspended in PBS, and one batch was inactivated by heating as described above. Single cell suspension was obtained by filtration through a 40 mm sieve and 3 × 103, 3 × 104, 3 × 105 cells and subsequently resuspended in 200 mL PBS and infused in the tail vein. Control mice received 200 mL of PBS. Two hours postadministration, mice were sacrificed by cervical dislocation and blood was collected in serum separation tubes (Minicollect; Greiner Bio-One, Alphen a/d Rijn, The Netherlands) and centrifuged at 3,000 rpm for 10 min.
Lungs and livers were collected and snap frozen in liquid nitrogen. Tissues were stored at −80°C until use. All experiments were approved by the Central Committee Animal Experiments Ethics Committee (license number AVD101002016635) and complied with the 1986 Directive 86/609/EC of the Council of Europe.
mRNA expression analysis
RNA was isolated from frozen mouse lung and kidney tissues using TRIzol reagent (Invitrogen, Life Technologies, Carlsbad, CA) and cDNA was synthesized from 1,000 ng RNA with random primers (Promega, Leiden, the Netherlands). Quantitative gene expression was determined using TaqMan gene expression master mix (Life Technologies) and assay-on-demand primer/probes for CCL2 (Mm00441242_m1), CCL3 (Mm00441258_m1), IL-10 (Mm00439614_m1), TGF-b (Mm01178820_m1), IL-1b (Mm01336189_m1), and housekeeping gene hypoxanthine guanine phosphoribosyltransferase (HPRT) (Mm01545399_m1); all assay on demand primers are from Applied Biosystems, Foster City, CA). Results were expressed as copy numbers (efficiency-DCT) ratio to HPRT.
Cytokine and chemokine measurements in mouse serum
In mouse serum samples, levels of IL-10, CXCL5, G-CSF, IL-6, CCL2, and CXCL1 were measured. Cytokine levels were quantified using a Mouse cytokine/chemokine magnetic bead panel multiplex assay (Merck Millipore). Cytokine levels were measured by a Luminex 100/200 cytometer (Luminex, Austin, TX) using Xponent software.
Analysis of human peripheral blood mononuclear cells—MSC coculture conditioned medium
Conditioned medium was generated by coculturing human MSCs or HI-MSCs with peripheral blood mononuclear cells (PBMC) at doses of 1:5, 1:10, and 1:20 for 24 h in 24-well flat-bottom plates in 500 μL of RPMI supplemented with 2 mM
Ex vivo addition of MSCs to whole blood
All human blood samples were collected from healthy volunteers. Whole blood samples were anticoagulated with lithium heparin. Around 4 × 105 MSCs and HI-MSCs were added per 1 mL of blood and samples were collected after 5 min, 15 min, 1, and 2 h of MSC addition. Samples were centrifuged at 500 g for five minutes for cell depletion, then at 2,000 g for 10 min at 5°C for platelets removal. Collected plasma samples were stored at −80°C until use.
Results
High dose of MSCs needed for systemic immune effects
To examine whether the in vivo immunomodulatory effect of MSCs is dose dependent, healthy C57BL6 mice were infused through the tail vein with 0, 3 × 103, 3 × 104, or 3 × 105 syngeneic MSCs per animal and serum collected after 2 h for cytokine and chemokine analysis. Doses of 3 × 103 and 3 × 104 MSCs were ineffective in inducing detectable systemic effects (Fig. 1). Doses of 3 × 105 MScs induced significant elevations in the cytokines, IL-10, G-CSF, IL-6, CCL2, and CXCL1 (Fig. 1A, C–F), and a significant drop in the neutrophil chemoattractant CXCL5 in the serum (Fig. 1B), demonstrating that relatively high doses of MSCs are needed to induce systemic immunomodulatory effects.
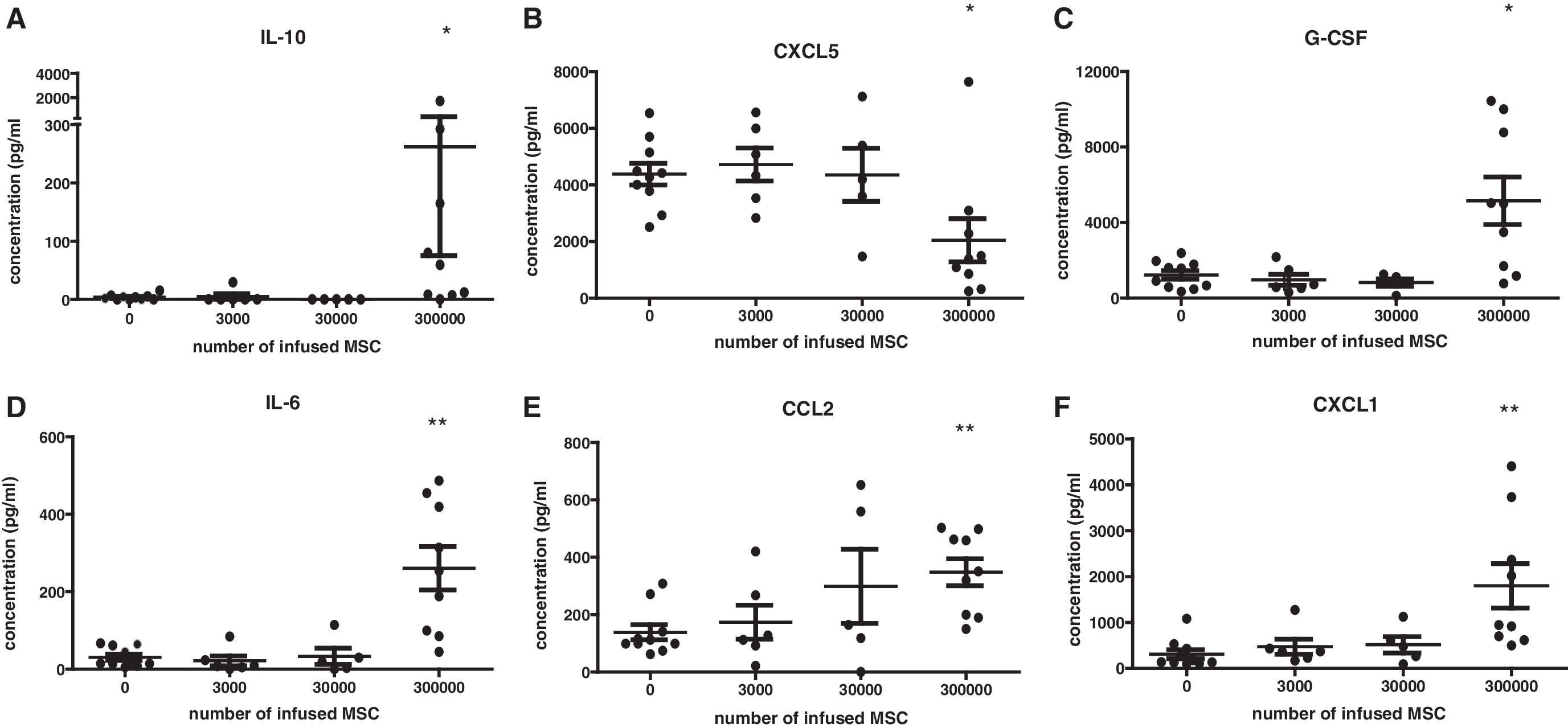
The effect of mouse MSCs on serum cytokine levels. Different doses of syngeneic MSCs were infused through the tail vein in healthy mice and at 2-h serum cytokine levels were determined by Luminex Magnetic Bead Assay.
Dose-dependent cytokine response in lungs and liver after infusion of MSCs
Intravenously infused MSCs largely accumulate in the lungs because they are unable to pass the pulmonary capillary network due to their large size. The lung environment may, therefore, respond to MSC infusions by changes in cytokine and chemokine levels and to detect whether the magnitude of this response is dependent on the dose of MSCs, 0, 3 × 103, 3 × 104, and 3 × 105 MSCs were injected in the tail vein of healthy mice. After 2 h, dose-dependent effects on chemokine and cytokine mRNA levels in lung tissue were measured. The monocyte chemoattractant CCL2 increased after infusion of only 3 × 103 MSCs and this increase became more significant upon infusion of 3 × 104 and 3 × 105 MSCs (Fig. 2C). IL-10 levels were increased from 3 × 104 MSCs (Fig. 2A). TGFβ and CCL3 mRNA levels showed a small but significant increase from 3 × 104 MSCs and was further elevated upon infusion of 3 × 105 MSCs (Fig. 2B, D).
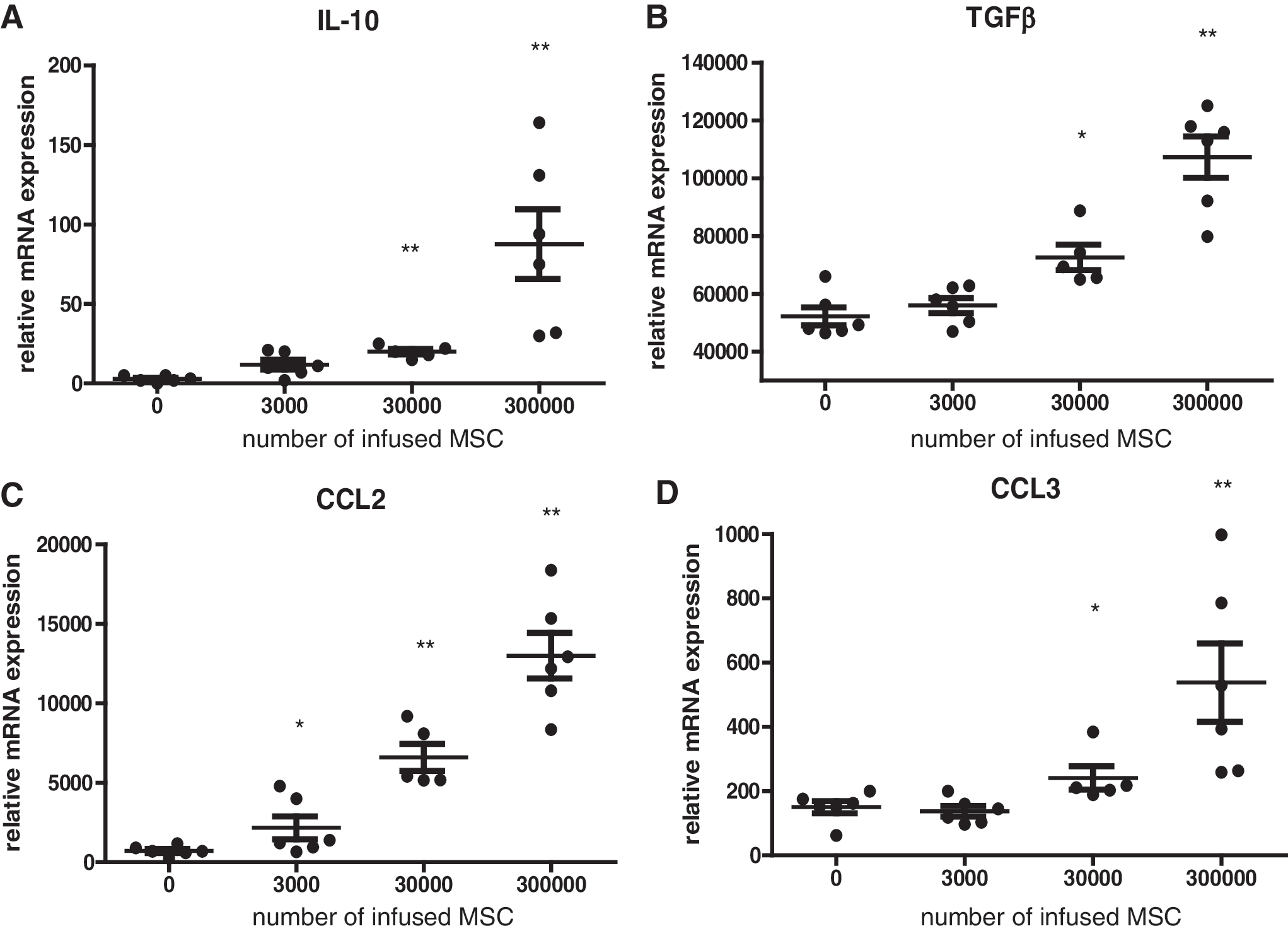
mRNA expression in mouse lung tissue 2 h after infusion of different doses of syngeneic MSCs detected by RT-PCR. Data expressed relative to HPRT.
We previously reported that MSCs are phagocytosed by monocytic cells, which distribute through the bloodstream to other sites, in particular, the liver [22]. To examine whether a dose-dependent effect of MSCs could be detected in liver tissue, we measured mRNA levels in liver tissue 2 h after administration of the abovementioned dosages of MSCs. mRNA levels of CCL2 became elevated even at the lowest dosage administered (Fig. 3C). Changes in TGFβ and CCL3 levels became significant at the dosage of 3 × 104 (Fig. 3B, D). A significant elevation of IL-10 was detected at the 3 × 105 dosage (Fig. 3A).
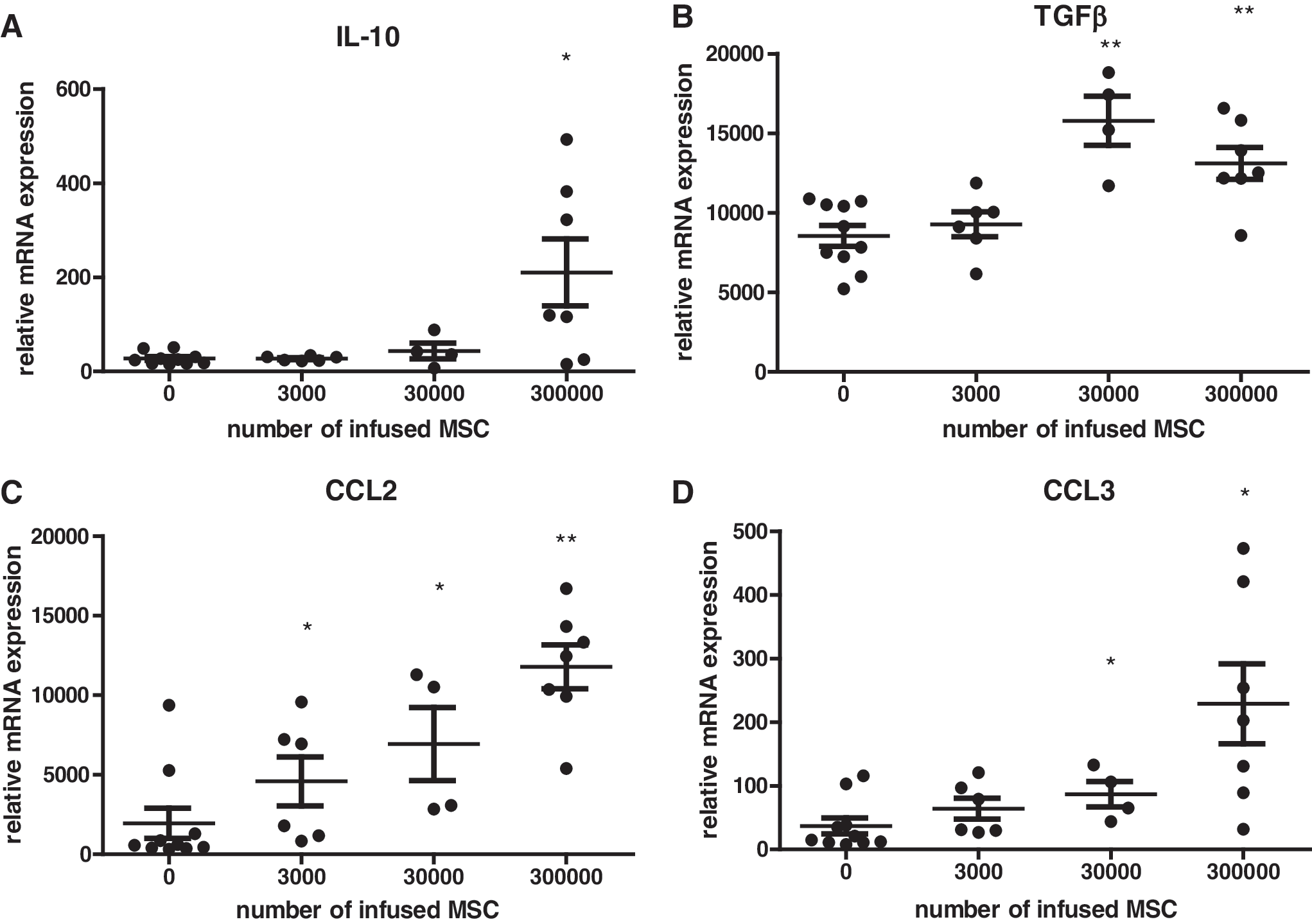
mRNA expression in mouse liver tissue 2 h after infusion of different doses of syngeneic MSCs detected by RT-PCR.
Dose-dependent effects of human MSCs and HI-MSCs in vitro
To explore whether the effects of different doses of MSCs can be confirmed in vitro and to study whether inactivation of MSCs through heating to 50°C, rendering secretome-deficient HI-MSCs maintain their dose-dependent effect, we added human MSCs and HI-MSCs at ratios 1:20, 1:10, and 1:5 to human PBMC and measured cytokine levels after 24 h. MSCs and HI-MSCs by themselves did not produce high levels of the measured cytokines. MSCs elevated levels of IL-6, CXCL5, CXCL1, and CCL2 in PBMC cocultures dose dependently (Fig. 4). Unlike MSCs, HI-MSCs did not induce cytokine secretion. This result demonstrates that the elevated cytokine levels in cocultures with control MSCs come from the MSCs or are derived from PBMC after stimulation by control MSCs and that this effect is abolished by inactivation of MSCs.

Effect of human MSCs and HI-MSCs on cytokine levels in MSC-PBMC coculture supernatant. MSCs and HI-MSCs were cocultured with PBMC at different ratios and supernatant cytokine levels determined after 24 h by Luminex Magnetic Bead Assay. Data shown represents the mean with SEM of MSCs from 10 different donors.
HI-MSCs and IFN-γ pretreated MSCs act in the same way as control MSCs in vivo
We then explored how HI-MSCs affect systemic cytokine levels in vivo. Two hours after administration of 3 × 105 MSCs and HI-MSCs in healthy mice, similar increases in systemic IL-10, G-CSF, and IL-6, and decrease in CXCL5 were detected in both groups, suggesting that MSCs without any activity result in similar cytokine changes as the control MSCs (Fig. 5A–D). To further assess the effect of MSC activity, MSCs were enhanced by pretreatment with IFN-γ (50 ng/mL), which stimulates their antiproliferative effects on immune cells. Similar increases in systemic G-CSF, IL-6, and IL-10 were detected in the IFN-γ-stimulated groups and non-IFNγ-treated groups (Fig. 5A, C, D). All types of MSCs decreased CXCL5 levels (Fig. 5B). Overall, heat inactivation and IFN-γ pretreatment had no major additional effect on cytokine levels compared with control MSCs. These data suggest that the changes in serum cytokine levels upon infusion of MSCs are derived from host cells and that MSCs act as a trigger for the response.
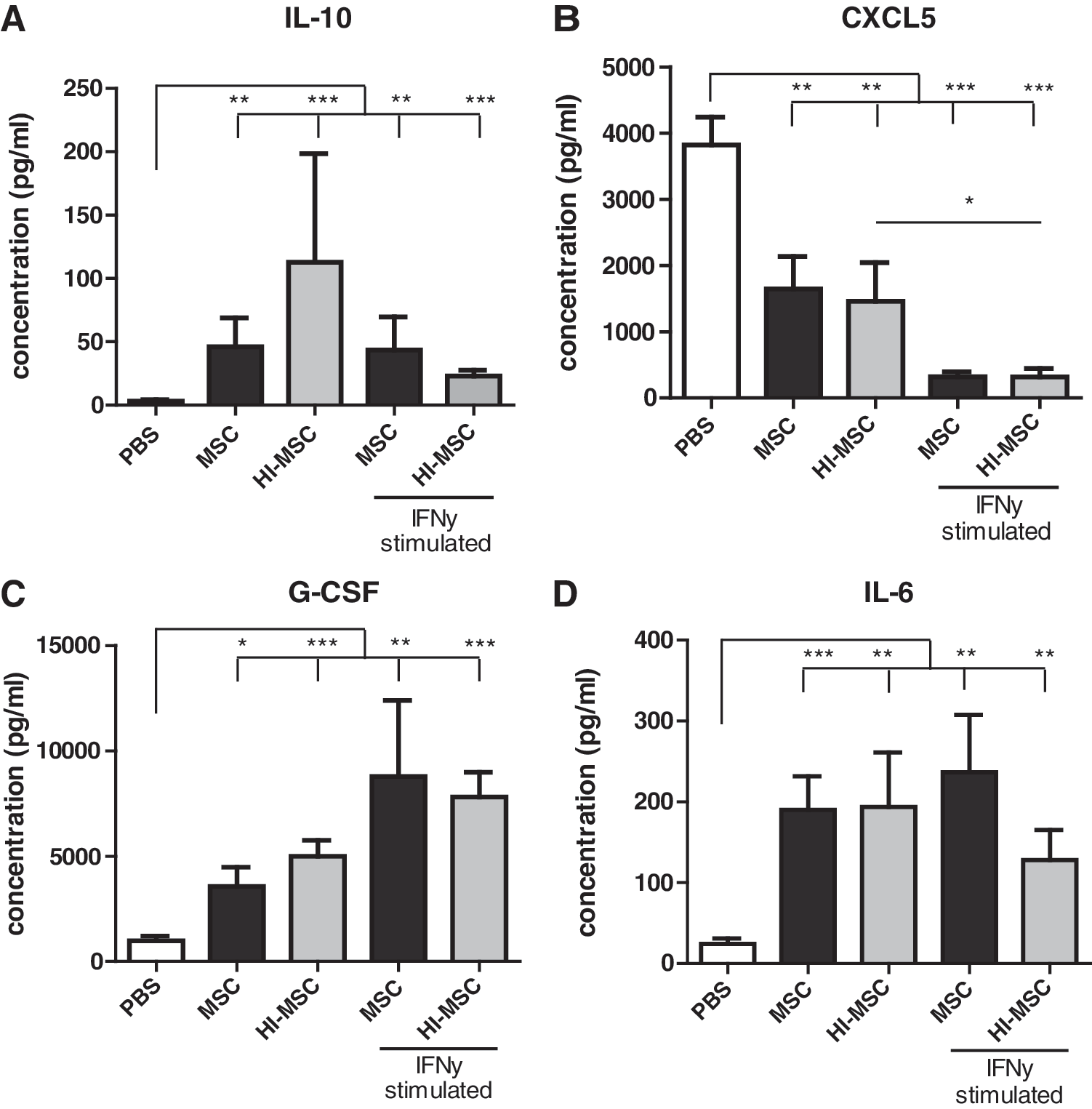
Effect of IFNγ treatment and heat inactivation of mouse MSCs on serum cytokine levels 2 h after infusion of 300,000 syngeneic MSCs detected by Luminex Magnetic Bead Assay.
Effects of MSCs and HI-MSCs on cytokine levels in human whole blood
To examine whether MSCs and HI-MSCs would induce similar fast cytokine responses in human whole blood as observed in mice, we added human MSCs to heparinized whole blood. As soon as 5 min after the addition of MSCs, we observed a small but significant increase in the level of CXCL5 (Fig. 6B). IL-10 levels showed a tendency to increase upon the addition of MSCs and HI-MSCs but were only significantly elevated at 2 h after addition of HI-MSCs (Fig. 6A). Along with CXCL5 and IL-10, a rise in IL-6 levels was observed after 5 min of adding MSCs, and further increased after 1 and 2 h. This increase was less with the addition of HI-MSCs, suggesting that IL-6 may be secreted by control MSCs (Fig. 6D). There were no significant changes in CCL2 (Fig. 6C).
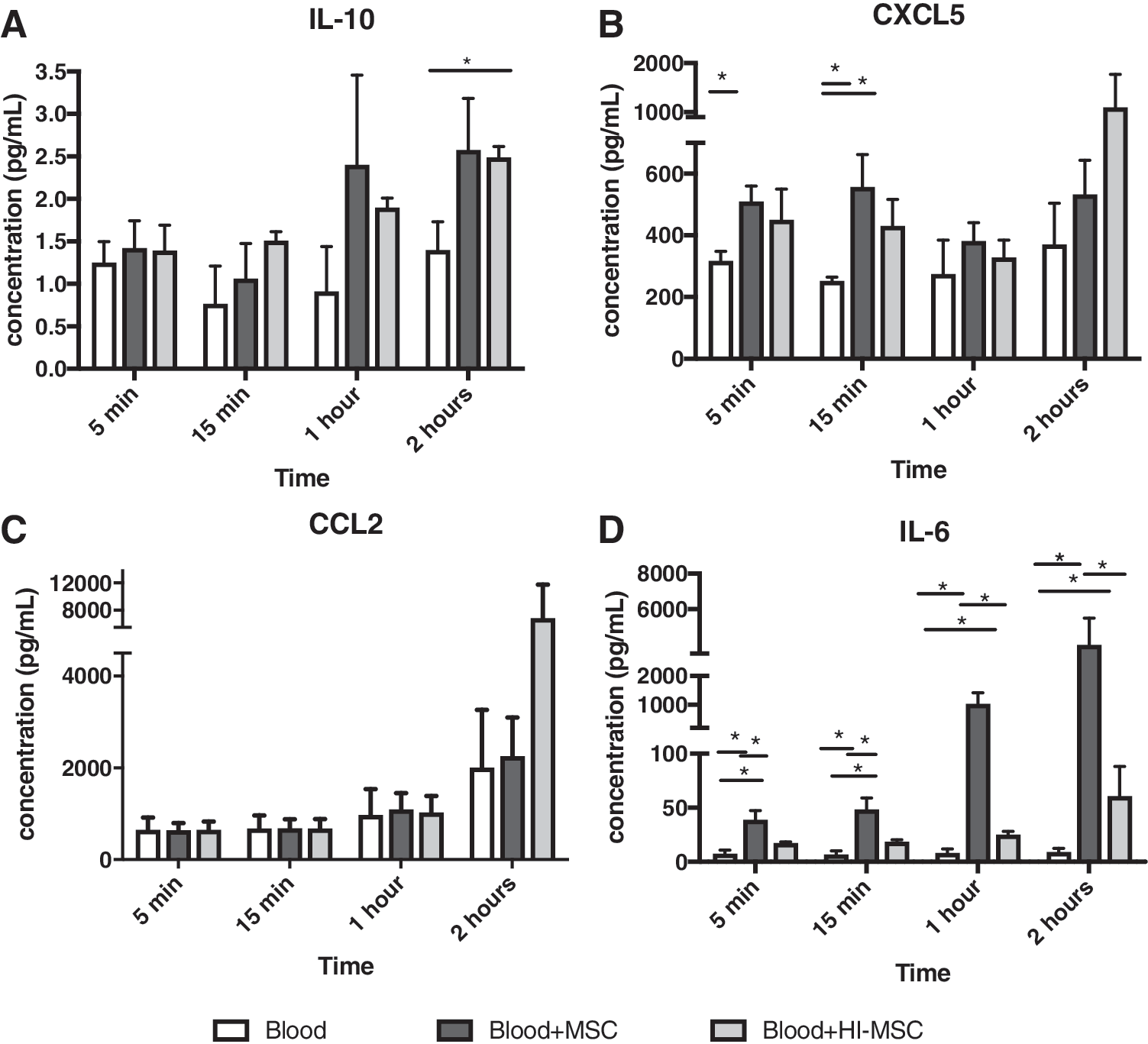
Changes in
Discussion
While the use of MSCs as a therapy for a variety of diseases has been studied extensively, there is a lack of consensus on the optimal dosage for best therapeutic outcomes. Even though the most commonly used route of administration of MSCs in animal models and early clinical trials is through intravenous infusion, most studies elucidating MSC dosage have been conducted using focal injections. The few studies testing different doses of intravenously administered MSCs used highly disease-specific models [30 –33]. Due to the complexity of these models, the association between dose and outcome is rather paradoxical. In the present study, we used healthy mice to unambiguously demonstrate that the effect of intravenously injected MSCs is dose dependent.
A significant change in serum cytokine levels was only detectable upon infusion of 3 × 105 MSCs per mouse. This dose would equal roughly 15 × 106 MSCs per kg body weight in humans, which is a very high and seldom administered dose in clinical studies [32]. We therefore reduced MSC dosage by 10-fold and 100-fold to determine if fewer MSCs can be used to achieve similar effects. Systemically, cytokine changes were not observed at the lower dosages, indicating that the MSC response is dose dependent and that a high dose of MSCs is required for modulation of serum cytokine levels.
This would suggest that high doses of MSCs are required to achieve therapeutic effects of MSCs in clinical trials. However, the most commonly used approach to extrapolate dosage from animal to human in clinical trials is allometric scaling [34]. Based on this method, the human dosage is calculated to be 1.28 × 106 MSCs per kg body weight for a 60 kg individual. In fact, clinical trials commonly use doses of 1–2 × 106 MSCs/kg body weight. Interestingly, a recent study in human treated with LPS demonstrated that infusion of 4 × 106 MSCs per kg body weight was able to modify systemic cytokine levels, whereas lower doses were largely ineffective [35]. Altogether these data indicate that at the commonly used doses of 1–2 × 106 MSCs/kg body weight in clinical trials, systemic effects on cytokine levels are undetectable.
In the lung tissue, the anti-inflammatory cytokine IL-10 was significantly increased with only 3 × 103 MSCs per mouse, equaling 0.15 × 106 MSCs per kg body weight. Similarly, 3 × 103 and 3 × 104 MSCs per mouse modulated cytokine mRNA expressions in the liver, further supporting our previous observations [22] that infused MSCs are phagocytosed by macrophages and transported to the liver, or that MSC debris in the circulation is phagocytosed by liver resident macrophages. These results demonstrate that MSCs act through a dose-dependent mechanism and provide baseline data for the adjustment of MSC dosage according to specific disease conditions or individuals. The question is whether in addition to dosage, other factors such as the activity of MSCs also influence the effect of MSC therapy.
We generated HI-MSCs and IFN-γ pretreated MSCs to examine whether MSC therapy can be modified by abolishing or enhancing MSC activity and examined their effect on serum cytokine levels. We observed that changes in cytokine levels in response to IFN-γ preconditioned MSCs and secretome-deficient HI-MSCs were similar to control MSCs. While studies have shown that IFN-γ pretreated MSCs have increased expression of immunosuppressive factors, they also show elevated expression of MHC-I and II, which may hasten the clearance of MSCS by host cells [36 –39]. It is possible that the combination of these changes in MSCs in response to IFN-γ would result in an effect of IFN-γ-MSC that is not different from control MSCs. It is also possible that IFN-γ-MSC have other, nonmeasured, effects after administration different from control MSCs.
However, the syngeneic model employed for this study allows an unmasked analysis of pretreated MSC activity independent of rejection mechanisms. Our results demonstrate that enhancement of MSC immunosuppressive activity does not affect the role of MSCs on modifying systematic cytokine levels, indicates that the serum cytokine changes in response to MSC infusion are derived from host cells and that MSCs merely act as a trigger for the response. Whether the similar effects of IFN-γ preconditioned MSC, HI-MSC, and control MSCs on serum cytokine levels shortly after infusion lead to similar immunomodulatory effects in disease models requires further investigation.
To further elucidate the origin of the cytokine changes observed in vivo, we cocultured human MSCs and HI-MSCs with PBMC in various ratios in vitro. While MSC:PBMC cocultures showed clear dose-dependent changes in cytokine levels with increasing ratios of MSCs, HI-MSC:PBMC coculture cytokine levels were similar to PBMC alone. This observation suggests that the cytokine changes observed in MSC-PBMC cocultures came predominantly from MSCs. However, the present and previous [27] in vivo studies have shown that HI-MSCs do have a significant effect on serum cytokine levels. The discrepancy between the in vivo and in vitro models could be attributed to the differences in the immune milieu between the two models. The mechanism of action of HI-MSCs is contact dependent and appears to rely on the phagocytosis by phagocytic cells. The number of monocytes present in the PBMC used in the in vitro model is limited and they may not survive long enough to produce sufficient cytokines after phagocytosis of MSCs to be detected.
Furthermore, PBMC contain no granulocytes. Interestingly, we found that addition of MSCs and HI-MSCs to whole blood revealed immediate changes in IL-6 and CXCL5, suggesting a role for granulocytes in the rapid response to MSCs. This ex vivo model still could not fully capture all the components in the in vivo environment as the CXCL5 levels demonstrated an increase ex vivo rather than a decrease in vivo. Thus, in addition to granulocytes, other cell types, such as endothelial cells, may work synergistically to elicit MSC-induced effects in vivo. Further studies are needed to determine the cell types responsible for the dose-dependent changes in cytokine levels upon MSC administration.
MSCs express Tissue Factor (TF) and intravascular administration of TF-bearing cells can elicit an instant blood-mediated inflammatory reaction (IBMIR) [40]. IBMIR is an adverse innate immune response triggered by tissue cells and is accompanied by a coagulation response and systemic increases in TNFα, CCL2, IL8, and IL6 at 3 h after initiation [41]. It is possible that the rapid cytokine changes we observed after intravenous administration of adipose tissue-derived MSCs were the result of an IBMIR response. The timing of the cytokine changes we observed 2 h after MSC administration match with the cytokine elevations in IBMIR at 3 h. Therefore, the cytokine responses we detected in serum, lung, and liver tissue may be derived both from IBMIR and from immune cells that interacted with administered MSCs through soluble factors and by phagocytosing MSCs.
To conclude, the effect of MSCs on systemic cytokine levels and mRNA expression in the liver and the lung is directly dose dependent. This knowledge contributes to the design of rational MSC trials and search for clinical efficacy of MSC therapy. Understanding the basic characteristics of MSCs is crucial for the standardization of MSC therapy. Due to the complicated and intricate relationships between MSCs and the immune milieu, as well as the heterogeneous nature of MSCs, the precise dosage of MSCs needed for treatment is yet to be determined and will likely vary between disease type, MSC donors, and recipient. Our data indicate that high doses of MSCs are required to elicit systemic effects; low doses may be sufficient for local effects. This knowledge about the dose-dependent effects of MSCs can be used to adjust the amount of administrated MSCs according to disease type and the desired site of action.
Footnotes
Acknowledgment
The authors wish to acknowledge Nicole Lemmens for her help with the setup and maintenance of the Luminex cytometer.
Author Disclosure Statement
The authors declare that no competing financial interests exist for the work presented in this article.
Funding Information
A Mitacs Globalink research grant was generously provided as funding for this project.