
Abstract
Preclinical medical research has historically depended on either traditional two-dimensional in vitro cell culture or animal models for the purposes of disease modeling, including cancer pathophysiology, immunology studies, drug testing, and toxicity assays. However, both of these models have intrinsic flaws. Two-dimensional cell culture systems do not capture the heterogeneity of in vivo disease, being originally derived from a limited set of cell lines. They also differ from the physiology encountered in vivo, with the majority of pharmaceutical agents, including oncology drugs which are effective in vitro failing at clinical trials. Animal models have issues of cost, associated ethical concerns, and xenogeneity comparative to human systems. Organoids are three-dimensional (3D) cell culture models derived from human tissue, which have the potential to overcome the issues with the traditional models discussed above. They self-organize into 3D structures resembling their tissue of origin and recapitulate some of its functions. They have been shown to be excellent models for the purposes of disease modeling and high-throughput drug screening, among others. Despite these benefits, some challenges yet remain in organoid research. It is hoped that the combination of organoid culture with bioengineering approaches may successfully overcome these.
Background
Despite forms of medicine and even individual physicians being described as long ago as 2700 BC, the history of modern medicine is relatively recent. The first successful human organ transplant, a kidney, was performed by Joseph Murray and colleagues in 1954. Meanwhile, the first long-term remission and cure of hematologic malignancies were described in the postwar period after observing the leukopenic effects of mustard gas on soldiers in World War II [1,2]. Since then the development of multiple classes of immunosuppressants has revolutionized the field of organ transplant, targeted oncologic therapies such as cetuximab and trastuzumab are routinely used alongside traditional chemotherapy, and specific drugs have been developed for monogenic conditions such as hereditary hypercholesterolemia on the basis of the underlying molecular pathology of the disease [3,4].
Much of the advances in these fields have traditionally relied on the use of in vitro two-dimensional (2D) cell culture and animal models, whether for modeling of particular disease states, including cancer, genetic manipulation, or drug efficacy, and safety assays. However, translating effective drug therapies from cell or murine models to in vivo systems historically has an overwhelmingly poor success rate, with up to 90% of drugs successful in preclinical models failing at clinical trials [5,6]. Animal models have specific limitations with respect to greater cost, ethical issues, and xenogeneity for the purposes of physiology and disease modeling. Meanwhile, isolated 2D cell cultures, although derived from human tissue, can provide only a greatly simplified representation of the complex architecture and interactions inherent to in vivo tissue structures [7].
The recent emergence of three-dimensional (3D) cell cultures, or organoids, has the potential to overcome some of the deficiencies encountered with the traditional models described above. Organoids are defined as 3D structures derived from either pluripotent, neonatal, or adult stem cells, which spontaneously self-organize and undergo a degree of differentiation, producing functional cell types, and which have the capacity to undertake some functions of the relevant organ [8 –11]. Although rat organoid cultures were first described in 1992, they had a limited capacity to divide and self-maintain, with a culture lifespan of approximately only a month [12,13]. However, in 2009, a method of producing self-renewing intestinal organoids by culturing them in a Matrigel protein matrix was published by Sato et al [14]. This group had identified cycling crypt base stem cells by the unique Lgr5 genetic marker and confirmed that these intestinal stem cells had the capacity to propagate all cell lineages derived from the gut. Culture of crypts containing Lgr5+ intestinal stem cells, dependent on the presence or withdrawal of particular medium components, such as Wnt, Noggin, and R-spondin, continued to bud and undergo crypt fission. These were expanded into structures with differentiated crypt-villus type architecture, named organoids [14]. Following this, in vitro culture of an optic cup derived from mouse embryonic stem cells was described by Eiraku et al [15].
The successful culture of healthy organoids directly derived from human tissue in 2011 by Clevers' group and subsequently replicated by Batlle's group in 2015 represented a significant breakthrough [16,17]. Human intestinal crypts were isolated from small intestinal and colonic tissue as for murine organoids and cultured in a modified medium containing Wnt, gastrin, and nicotinamide, as well as inhibitors of Alk and p38. Clever's group in 2011 also successfully cultured organoids from human metaplastic (Barrett's) epithelium, mouse colonic adenomatous tissue, and human colorectal neoplastic tissue, having previously demonstrated that driver mutations in the Lgr5+ crypt stem cell lineage were responsible for initiation of neoplasia [16,18]. Since then human organoid cultures have been developed from multiple tissue types, including colorectal, gastric, pancreatic, liver, breast, prostate, cervical, neural, as well as culture of tumor-derived tissue-specific organoids, known as tumoroids [19 –27] (Fig. 1).
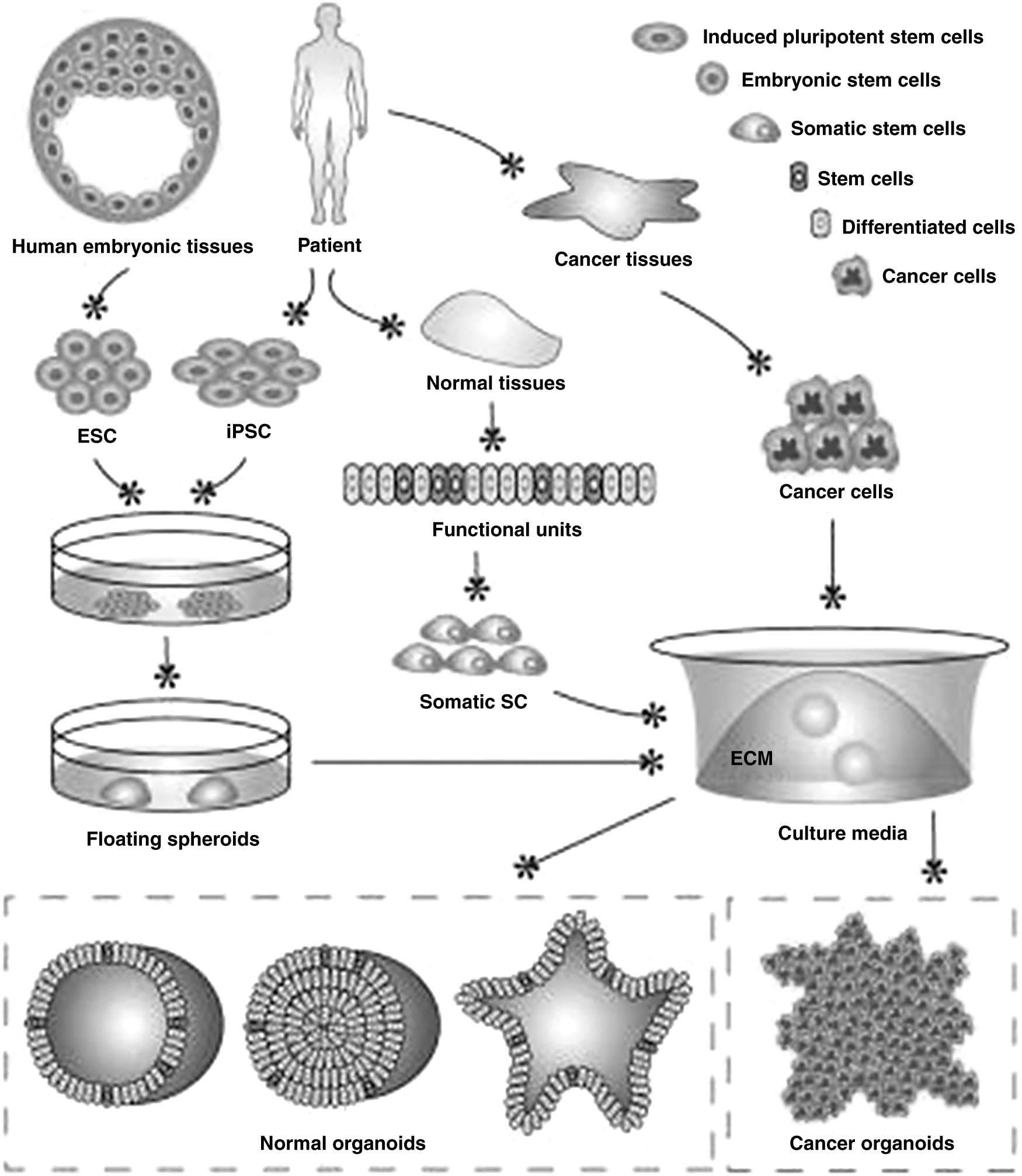
Schematic of organoid development [47]. Reused under Creative Commons License
Advantages of Organoids over Traditional Models
Comparative to conventional 2D in vitro models, 3D organoids achieve a greater level of structural and functional complexity more closely analogous to in vivo physiology. This is due to the presence of tissue-specific architecture, mechanical biochemical cues, cell–cell and crude cell–extracellular matrix (ECM) interactions, which are lacking in 2D monolayer culture systems [28,29]. However, they still allow experimentation under controlled conditions that are not possible to achieve in human clinical studies. They also remain genomically stable, with one study demonstrating genetic stability of hepatocellular carcinoma organoids over a period of a year [30,31]. Importantly, tumoroids also retain individual genetic and proteomic diversity related to the patient-derived tissue of origin [32 –34]. Some of the specific advantages of organoids over animal models or 2D monolayer cell cultures are described below (Table 1).
Timeline of Advances in Organoid Technology
Cell Morphology and Organization
Traditional in vitro cell cultures are arranged as a flat monolayer. The morphology of cells cultured in this manner differs greatly from cell morphology in the 3D in vivo state, thus potentially producing differing effects on cellular function and interactions [35]. Cells in organoid cultures self-organize in a spatial configuration similar to the tissue of origin. This confers differing properties and behavior dependent on the cell's spatial location in the unit, with cells at the outer margin of the structure exhibiting greater proliferation and motility and those at the center displaying more quiescent behavior [36]. These patterns do not occur in 2D cell culture. In addition, differential protein expression, gene expression, cell migration, cell organization, survival, and cell signaling have been observed in 3D comparative to 2D cell cultures, providing a more accurate representation of the processes and physiology in vivo [37].
Tumor Microenvironment and Interactions, Coculture
In the past years, the vital role of the tumor microenvironment in initiation of tumorigenesis, tumor progression, and metastasis has become increasingly apparent [38]. The multiple complex interactions and cross talk of tumor epithelial cells with the surrounding stroma and mesenchymal cells can create either a facilitatory or inhibitory landscape, and therapeutic strategies targeting such interactions, for example, immunomodulators and antiangiogenic agents, have been developed with good clinical effect [39,40]. Organoids recapitulate the key cellular interactions, microenvironment, and stromal compartments present in vivo, thus more accurately replicating cancer pathophysiology and interactions with the microenvironment which cannot be achieved in traditional in vitro cell culture [41,42]. In addition, it is also possible to further simulate the immune microenvironment and examine cell–cell interactions in a 3D setting by coculturing heterotopic organoid cell lines of interest, such as colorectal carcinoma organoids with fibroblast or lymphocyte cell lines [43 –45].
Retention of Genetic Diversity
In 2D cancer cell culture systems, loss of the genetic heterogeneity observed in the tumor tissue of origin consistently occurs after multiple passages [35]. In contrast, organoids retain the individual genetic and proteomic profiles of the tumor tissue of origin, while maintaining the common features of the disease. This has been demonstrated in organoids derived from a number of diverse tumor types, including hepatocellular, colorectal, gastric, and breast [32 –34,46]. Colorectal cancer organoids successfully cultured from different anatomical sites (right/left/rectal) and uncommon histological subtypes (mucinous, neuroendocrine) maintain a resemblance to the parent tumor with respect to histological subtype, mutational, and proteomic profile [34]. Breast cancer organoids also retain the hormonal profile and HER2 status of the original tumor [47]. This offers great potential for assessing tumor behavior associated with particular mutational profiles and precision individualized drug efficacy assays.
Drug Metabolism
As discussed above, the success rate of clinical drug development remains exceedingly low, with compounds demonstrating effectiveness in preclinical models often failing to translate this to clinical trials [5,6,48]. Oncology drugs have a significantly higher failure rate but simultaneously rapidly rising development costs [48,49]. One of the significant factors affecting this is the applicability (or lack thereof) of preclinical drug discovery models, including cancer models, to in vivo patient physiology. However, the ability of organoids to recapitulate cell–cell interactions and cell–ECM interactions allows them to more accurately reflect drug responses and metabolism than 2D in vitro models, while bypassing the xenogeneity and cost limitations associated with patient-derived xenografts. Successful use of patient-specific cystic fibrosis organoids for drug discovery of novel compounds and determining effectiveness of preexisting compounds on an individual basis, especially those with rare or uncharacterized cystic fibrosis transmembrane conductance regulator (CFTR) mutations has been demonstrated [50]. Other studies suggest that similar results can be achieved in the field of oncology, with patient-derived tumor organoids behaving as excellent models for assessing individual response to therapeutic agents [32,34,51]. Vlachogiannis' 2018 study in particular demonstrated extremely high sensitivity (100%), specificity (93%), positive (88%), and negative (100%) predictive value of patient-derived organoids response to chemotherapeutic agents respective to in vivo efficacy [52].
Organ Transplantation
A further application of organoid technology is for the purpose of developing transplantable functional tissues in place of human donor organ transplants. Most work in this field is with respect to development of transplantable functional hepatocyte tissue due to the potent regenerative capacity of the liver. Organoid cultures with the capacity to differentiate into functional hepatocytes and cholangiocytes have been developed from human pluripotent and embryonic cell lines [53 –55]. The functional capacity of these transplantable hepatic organoids has been assessed in animal models, including murine models of acute liver failure, with promising results. Transplantation of intestinal stem cells in experimental animal models of acute colitis has also been investigated, with the aim of promoting mucosal healing [56 –58]. Accelerated mucosal healing in acute inflammatory bowel disease (IBD) is associated with a more favorable prognosis, including decreased rates of hospital admission and need for surgical intervention [59,60]. As Matrigel, the original matrix utilized for organoid culture, is derived from nonhuman (murine) tissue, alternative matrices have been developed to provide a suitable nonimmunogenic culture medium for organoids intended for in vivo transplantation [61].
Challenges
Despite these benefits, challenges still remain in organoid research. These include the failure rate of cultures, the time taken to develop cultures to maturity, difficulties with scaling-up of organoid systems into larger and more complex tissue constructs, lack of external reproducibility of individual organoid cultures, and a method for systematic large-scale analysis of observed results [62].
Future Directions
Organoid technology thus holds great potential for the future of the fields of disease modeling, genetic engineering, personalized and regenerative medicine, and drug development. The application of targeted bioengineering approaches, computational-aided large-scale data analysis, and recently developed advances in genetic engineering to organoid models is likely to significantly progress their reproducibility and utility. Some of these approaches are described below.
Bioengineering
Bioprinting
Organoid cell lines are currently cultured with no spatial constraints in substrates such as Matrigel, a tissue scaffold formed from proteins secreted by mouse sarcoma cells. Three-dimensional printing technology has been utilized for the purpose of bioprinting, whereby a bioink comprising cell aggregates or organoids is imprinted along with the desired biomaterials (e.g., hydrogels, matrix components) onto a supporting medium via a computer-aided transfer process [28,29]. This renders a highly accurate, reproducible 3D model with organoids, biomaterial, and scaffold spatially aligned at the desired patterns, gradients, and densities set by the modeling software. This method will help to address both reproducibility issues and scaling-up of organoid cultures into larger, microanatomically faithful tissue constructs. It has been successfully used as a 3D bioprinted liver model, in which bioprinting hepatocyte organoids into a 3D model with hexagonal liver-type architecture improved morphological organization and cellular functioning [63]. However, challenges remain with successful vascularization and functionality of the final tissue constructs.
Biofabrication
Currently, mature intestinal organoids, while mimicking the crypt and villous-like structures which occur in the gut, do not reproduce true villi. Biofabrication techniques such as laser ablation allow rapid production of scaffolds which may be created to mimic intestinal topography, including villi and microvilli. These can subsequently be directly seeded with epithelial organoids or alternatively used as moulds to create hydrogel-based porous copies, which reproduce the microanatomy of the gut [62].
Organoid-on-a-chip
Intestinal motility and maintenance of unidirectional luminal flow are among the key physiologic functions of the gut. Flow patterns, mechanical deformation, shear stresses, and a degree of peristaltic motility can be simulated via epithelial cell-lined microfluidic platforms, mimicking gut physiology with greater accuracy [28,64 –66]. Combining gut organoid cultures with such platforms produces greater similarity to the in vivo state with respect to both the physical environment and cellular activity. These are being further utilized to investigate the gut-microbiome relationship in health and disease, particularly IBD, by inoculating the gut-on-a-chip organoid with selected bacterial cultures and examining the effect of these on the model system [67,68].
It is now well recognized that the tumor microenvironment is crucial to progression or inhibition of tumor initiation, growth, invasion, and metastasis. This is a complex environment, with varying chemical and protein transport systems, degrees of vascularization, and osmotic and hydrostatic pressure gradients. These can again be imitated by creating bioengineered microfluidic platforms and semipermeable scaffolds whose selected chemical and physical parameters can be closely controlled [69,70]. Organoid tumor cultures can be combined with these (tumor-on-a-chip technology) to more closely reflect the in vivo tumor pathophysiology and interactions with the microenvironment.
Interestingly, early investigations of tumor metastasis from the primary site to distant solid organs using such platforms have already occurred, with migration of colorectal cancer cells from colon to liver organoids linked by a metastasis-on-a-chip device described [69].
Genetic Engineering
Genetic manipulation of organoids and other preclinical research models has previously been performed via transfection of siRNA. However, this method has undergone recent significant advances with the development of the CRISPR-Cas9 gene editing system, whereby specific predesigned genetic mutations may be introduced in a controlled manner into live cells with a low rate of off-target mutations [71]. Studies in organoid cultures derived from patients with cystic fibrosis have successfully utilized CRISPR-Cas9 delivery methods to replace the defective CFTR gene with a functional copy, demonstrating the feasibility of the method [72,73]. Unlike cystic fibrosis, sporadic cancer is a polygenetic disease. However, the role of particular driver mutations as key events in the initiation and progression of tumor growth and invasion is amenable to study via tumor organoids genetically edited using CRISPR-Cas9 to contain selected mutational landscapes, for example, knockdown of APC, TP53 with activating mutations of KRAS, and PIK3CA in colorectal cancer, and loss of the BRCA1 tumor suppressor gene in breast cancer [74 –76].
Biobanking
Organoids represent excellent models for assembling biobanks of patient-derived tissue. With respect to cancer modeling, matched healthy and tumor-derived organoid cultures can be derived from an individual patient. Highly invasive procedures are not necessary to acquire sufficient tissue for successful organoid culture; tumor organoids derived from colorectal liver metastases have been cultured from single 18G needle biopsy specimens with a success rate of 71%, with a subsequent series confirming the feasibility of the method with a success rate of 60% [77,78]. In addition, the successful culture of metastatic prostate cancer organoids from liquid biopsy, that is, blood, has been described, although this technique is not well established [79]. The significance of organoid biobanks relates to the volume of data that can be established. For current in vitro cancer drug screening, even where large number of cell lines are utilized, the numbers for each individual cancer type are low, and uncommon subtypes exponentially more so. The success of detecting significant drug effect patterns is therefore correspondingly low. However, large-scale biobanking of diverse cancer-derived organoids allows capture of disease heterogeneity, including of rare genetic variants, computational analysis of tumor genetic mutational profiles and potential for identification of predictive biomarkers [46,80]. Patterns of correlation between particular genetic mutational profiles, clinicopathological cancer phenotypes, and drug responses or resistance may be analyzed [81,82], and individualized therapeutic and toxicity drug assays may be performed, facilitating tailored oncologic treatment based on the response [32,34,51].
Drug Sensitivity Assays and Treatment Response
Organoids compare favorably to patient-derived xenografts in terms of both cost and culture cycle length. Although they remain more costly than 2D in vitro models, it has been shown that drugs are metabolized and develop chemoresistance in a more physiologically similar manner in organoid cultures comparative to their 2D model counterparts [83]. Studies examining the utility of organoids as drug screening models have shown responses consistent with the molecular profile of the original tumor, including in colorectal and prostate cancer organoids [76,79], and work comparing drug responses in matched organoid and patient-derived xenograft models has demonstrated concordance between the two [78]. In a similar manner to the discussion of biobanking above, the large volume of molecular profiles for any given cancer which can be derived from organoid culture lends itself to high-throughput drug screening. Organoid cultures can be grown in multiple microwells of up to 96 and 384-well formats, therefore allowing multiple samples and concentrations to be examined while using only small volumes of costly and potentially difficult to access developmental drugs [83]. The use of optical metabolic imaging and mathematical modeling techniques to accurately record and analyze culture growth and drug response will also facilitate drug discovery via organoid models. Finally, a large prospective observational trial, TUMOUROID, (NL49002.031.14) has been recently established to assess if patient-derived tumor organoids accurately reflect drug response in patients receiving treatment for metastatic colorectal, breast, or nonsmall cell lung cancer [76] (Fig. 2).
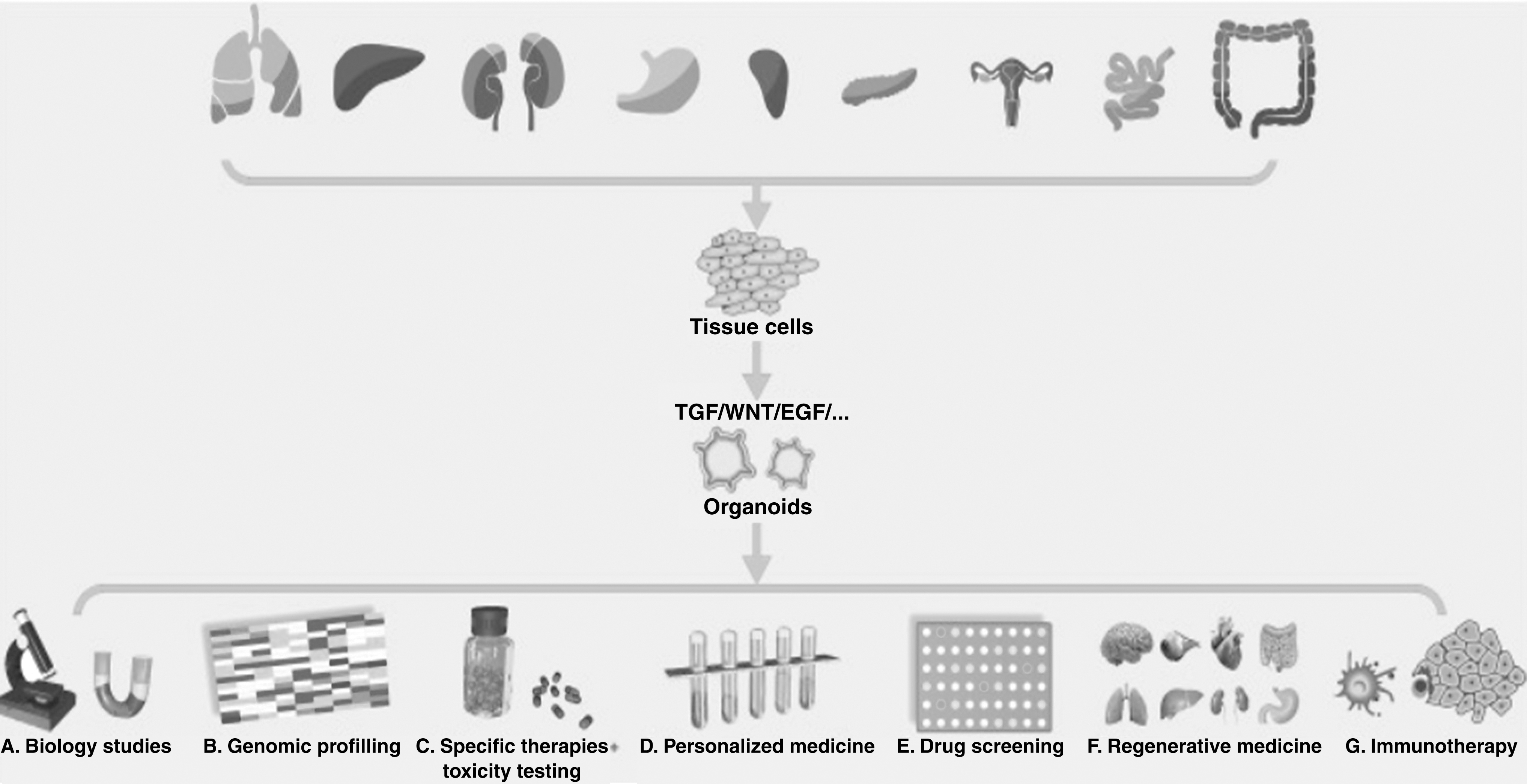
Potential applications of tumor organoids [7]. Reused with permission.
Conclusion
Organoids present an exciting preclinical model with multiple advantages over traditional in vitro cell cultures and animal models in terms of providing genetically faithful, more physiologically accurate representations of cancer biology, development, and metastasis and drug metabolism and resistance. While challenges yet remain with organoid cultures, including reproducibility and scaling-up of organoid systems, these are likely to be addressed by combining organoid culture with a variety of bioengineering approaches such as those described above. The results of ongoing trials and drug discovery via organoid models, such as the TUMOUROID trial described above, are eagerly awaited to demonstrate the further potential of this novel technology.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.