
Abstract
Stress urinary incontinence (SUI) after prostate surgery is primarily caused by urethral sphincter damage. There are few effective therapeutic approaches for male SUI due to both insufficient study of the structure of the external urethral sphincter (EUS) and incomplete understanding of the resident EUS stem/progenitor cells. The goals of this study were to localize and to determine the distribution of tissue-resident stem/progenitor cells in the male EUS throughout EUS development and to understand the anatomic temporal patterns of the EUS. Newborn Sprague Dawley rats were intraperitoneally injected with the thymidine analogue, 5-ethynyl-2-deoxyuridine (EdU), and the EUS was harvested at five time points (1, 2, 3, 4, and 8 weeks postinjection). The tissue was then processed for EdU staining and immunofluorescence staining for stem cell markers Ki67 and proliferating cell nuclear antigen. We counted the EdU+ label-retaining cells (LRCs) at each time point and colocalized with each stem cell marker, also we isolated and cultured the cells in vitro. The results revealed that the number of EdU+ LRCs in each EUS cross-section decreased over time and that the LRCs were located immediately under the basal membrane of laminin, densely adherent to the muscle fibers. In addition, the thickness of the striated muscle layer developed much faster than the smooth muscle layer during EUS development. By 4 weeks, the structure of the EUS layers was well differentiated. The EUS resident stem/progenitor cells were isolated with MACS® MicroBeads system, and myogenesis was confirmed. In this study, we defined both the time-course development of the EUS and the distribution of resident stem/progenitor cells. This information is crucial for forthcoming studies regarding male micturition and for development of novel therapeutic approaches for postoperative male SUI.
Introduction
Stress urinary incontinence (SUI) after prostate surgery may cause emotional distress by delaying a patient's return to pleasurable activities and by inhibiting relationships and financial distress by delaying return to the labor force [1]. For prostate cancer patients, robot-assisted radical prostatectomy (RAP) has become the standard surgical procedure for extirpation of prostate cancer [2]. Even with the improved visibility that this surgical modality provides the surgeon, many patients develop moderate-to-severe SUI after RAP. Glazener et al. reported that the incidence of SUI after RAP is as high as 74.3%, dropping to 46.8% after 1 month, 21.4% after 3 months, 13.6% after 6 months, and 9% after 12 months [3]. For patients with benign prostatic hyperplasia (BPH), despite the introduction of novel modalities for prostate obstruction, transurethral resection of the prostate (TURP) remains the gold standard for surgical management of BPH. Early incontinence occurs in up to 30%–40% of patients, and the ejaculatory ducts may also been injured during TURP [4,5]. Postoperative SUI is primarily caused by injury to or deficiency of the urethral sphincter after prostatectomy [6]. There are several currently available treatments for SUI, including pelvic floor muscle training, urethral bulking agents, male sling, and artificial urinary sphincter. However, an ideal treatment option for SUI has not yet been developed [7].
Stem cell-based therapies have received considerable attention as a potential therapy for regenerating defective tissues or organs [8]. In recent years, multiple preclinical studies have explored the use of stem cells in animal models as a potential treatment option for SUI. Because of their multidifferentiation potential, stem cells can be induced to differentiate into myoblasts with the objective of alleviating urethral sphincter dysfunction. Stem cells derived from striated muscle (STM), adipose tissue, bone marrow, and urine have been investigated. Thus far, however, there has been an insufficient therapeutic response to various stem cell therapies and the risks of these procedures have not been clearly established [9]. Recent studies have investigated the potential for activation of tissue-resident endogenous stem cells as a therapeutic approach for tissue regeneration. This avoids the need for complex procedures to harvest, cultivate, and transplant stem cells. In situ activation of stem cells has the potential to provide a direct strategy for regenerative medicine to overcome current transformational disadvantages associated with stem cell transplantation [10].
The male external urethral sphincter (EUS), a layer of the urethra composed of striated musculature, plays a crucial role in male urinary continence. The basal tonic activity of the EUS during bladder filling contributes to urethral closure. EUS dysfunction or damage may cause both SUI and ejaculatory dysfunction [11,12].
In 1994 Southgate et al. reported the technique for the isolation of and long-term culture of human urothelial cells using high calcium medium [13]. Larsson et al. found that isolated cloned human urothelial cells can self-renew in vitro and fully differentiate in vivo [14]. In addition, there have been multiple studies investigating the effects of mesenchymal stem cells on neovascularization and the recovery of urinary incontinence [15,16]. In 2019, we successfully localized and isolated female STM stem cells [17]. Few studies, however, have researched the location or distribution of the resident stem/progenitor cells in the male urethra.
Tissue-resident stem/progenitor cells exist in every organ. Owing to a lack of specific markers, identification of these cells is accomplished through a combination of immune staining for putative stem cell markers with the “label-retaining cell (LRC)” strategy [18]. A complete understanding of the morphological features of the male EUS including localization and characterization of the resident stem/progenitor cells will provide an anatomical basis for pathophysiological and therapeutic studies into male SUI. The aims of this study were to determine how the EUS develops, to describe the musculature of EUS, and to define and localize the tissue-resident stem/progenitor cells in the male EUS.
Materials and Methods
Animals
All animal experiments in this study were approved by the Institutional Animal Care and Use Committee at our institution. A total of 30 male neonatal Sprague Dawley rat pups delivered by primiparous rats were used for the 5-ethynyl-2-deoxyuridine (EdU)-retaining study. Each pup received an intraperitoneal injection of EdU (50 mg/kg; Invitrogen, Carlsbad, CA) immediately after birth. Six rats were sacrificed at each time point (1, 2, 3, 4, and 8 weeks postinjection) for EUS tissue harvest.
Isolation of EUS tissue-resident stem/progenitor cells and culture
EUS resident stem/progenitor cells were isolated as previously reported [19]. In brief, the EUS STM from 2-week-old rats was enzymatically dissociated, first with 0.2% collagenase II and then with 0.04 U/mL dispase. Nonmuscle tissue was gently removed under a microscope. The cell suspension was filtered through a 70 μm nylon filter (Falcon). Streptavidin beads (Miltenyi Biotech) were then added to the cells along with antibodies for integrin-α7–phycoerythrin and CD34–Alexa647 (eBioscience). This was followed by magnetic depletion of biotin-positive cells. The isolated stem cells were then cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% fetal bovine serum in a 10-cm dish. The myotube induction medium was DMEM supplemented with 2% horse serum (HS).
Preparation of tissue sections
Rats were anesthetized with 5% isoflurane and sacrificed by thoracotomy. After euthanasia, the rat urethras were harvested and fixed for 4 h with cold 2% formaldehyde and 0.002% picric acid in 0.1 M phosphate buffer. This was followed by overnight immersion in buffer solution containing 30% sucrose. Tissues were frozen in optimum cutting temperature compound (Sakura Finetic USA, Torrance, CA) and stored at −80°C for the further use. Sections were cut by 5 μm and affixed to charged slides for immunofluorescence (IF) staining as previously reported [20].
Phalloidin staining
Tissue sections were incubated with Alexa-488/594-conjugated phalloidin (1:500; Invitrogen), which was diluted 1:200 in 1% bovine serum albumin for 20 min at room temperature. After rinsing with phosphate-buffered saline (PBS), the tissues were stained with 40, 6-diamidino-2-phenylindole (DAPI, 1 mg/mL; Sigma-Aldrich, St. Louis, MO) for nuclear staining.
IF staining
Frozen tissue sections stored at −80°C were placed in cold methanol for 10 min, washed twice in PBS for 5 min and once in PBS/0.3% Triton X-100 for 5 min, then incubated with 3% goat serum at room temperature for 30 min. After draining the solution from the tissue section, the slides were incubated overnight at 4°C with primary antibodies, including antimyosin heavy chain (MHC; 1:500; Abcam, Cambridge, MA), anti-α-smooth muscle actin (αSMA; 1:500; Abcam), anti-Ki67 (1:500; Abcam), antiproliferating cell nuclear antigen (PCNA) (1:500; Abcam), anti-Laminin (1:500; Abcam), CD34 (1:500; Santa Cruz Biotech, Santa Cruz, CA), integrin 7α (1:500; Santa Cruz Biotech), and antimyogenin (1:500; Abcam). Secondary antibodies used included Alexa-488 and Alexa-594 conjugated antibodies (1:500; Invitrogen). Nuclear staining was performed with DAPI. For tracking EdU+ LRCs, tissue sections were incubated with EdU Click-IT reaction cocktail (Invitrogen) for 30 min at room temperature.
Myotube formation assay
The isolated EUS resident stem/progenitor cells were harvested and stained for MHC (1:500; Abcam). This was followed by induction with 2% HS then microscopy and photography at 7 days for the detection of myotube formation.
Image and statistical analysis
Tissue slides were examined with a Nikon Eclipse E600 fluorescence microscope and photographed with Retiga 1300 Q-imaging camera using the ACT-1 software (Nikon Instruments, Inc., Melville, NY). The images were then quantified using Image-Pro Plus image software (6.0; Media Cybernetics). For statistical analysis, five randomly selected fields per slide were examined and analyzed with Prism 8 (GraphPad Software, Inc.). Data were expressed as mean ± standard deviation. One-way analysis of variance followed by Bonferroni post hoc analysis and chi-square test was performed for statistical significance among multiple groups (P < 0.05/0.001).
Results
Development of EUS from 1 to 8 weeks
The average thickness of the STM layer (stained by MHC) developed much faster than the smooth muscle layer (stained by αSMA) (Fig. 1A). The thickness of the STM layer was 0.168 ± 0.050, 0.274 ± 0.065, 0.415 ± 0.091, 0.644 ± 0.028, and 1.136 ± 0.211 mm at 1, 2, 3, 4, and 8 weeks after birth, respectively. The STM layer thickness at every time point differed significantly by group (*P < 0.01) (Fig. 1B). The thickness of the smooth muscle layer was 0.029 ± 0.001, 0.053 ± 0.009, 0.058 ± 0.005, 0.060 ± 0.007, and 0.082 ± 0.015 mm, respectively. The smooth muscle layer thickness at 2, 3, and 8 weeks differed significantly compared with the previous time points (**P < 0.01) (Fig. 1C). The ratio of STM/smooth muscle was 5.79, 6.37, 7.16, 10.73, and 13.85, respectively. The results showed a significant change at 2 and 3 weeks compared with the previous time point; the values were statistically significant (*P < 0.05). At 4 and 8 weeks, the values were more statistically significant (**P < 0.01) (Fig. 1D).
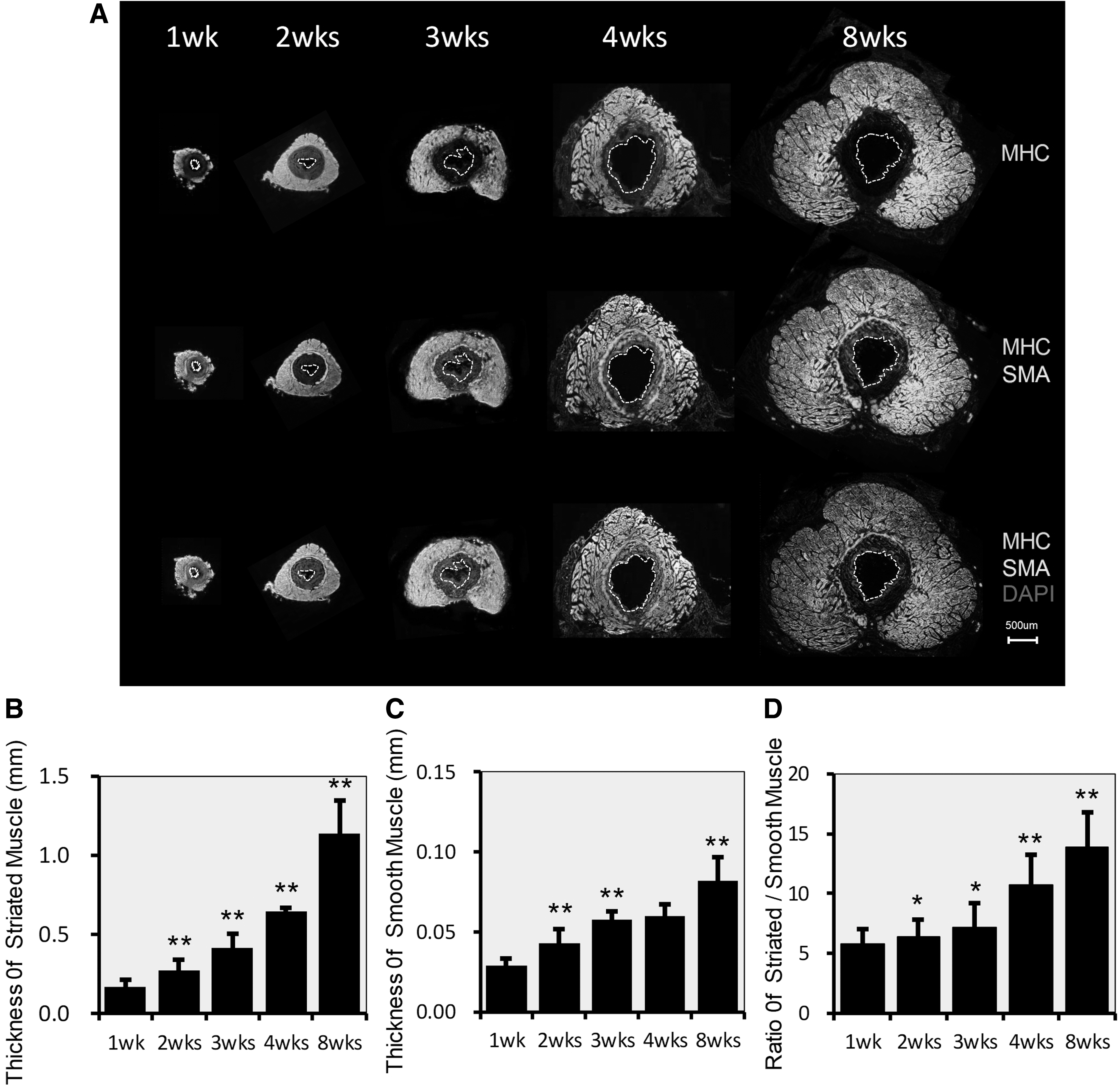
Development of the EUS from 1 to 8 weeks.
Musculature structure of EUS
We found that as the EUS developed from 1 to 8 weeks, the structure of the different layers became clear at 4 weeks and matured by 8 weeks (Fig. 2). These layers include the mucosal layer, submucosal layer, smooth muscle layer, inner longitudinal STM layer, and outer circular STM layer.
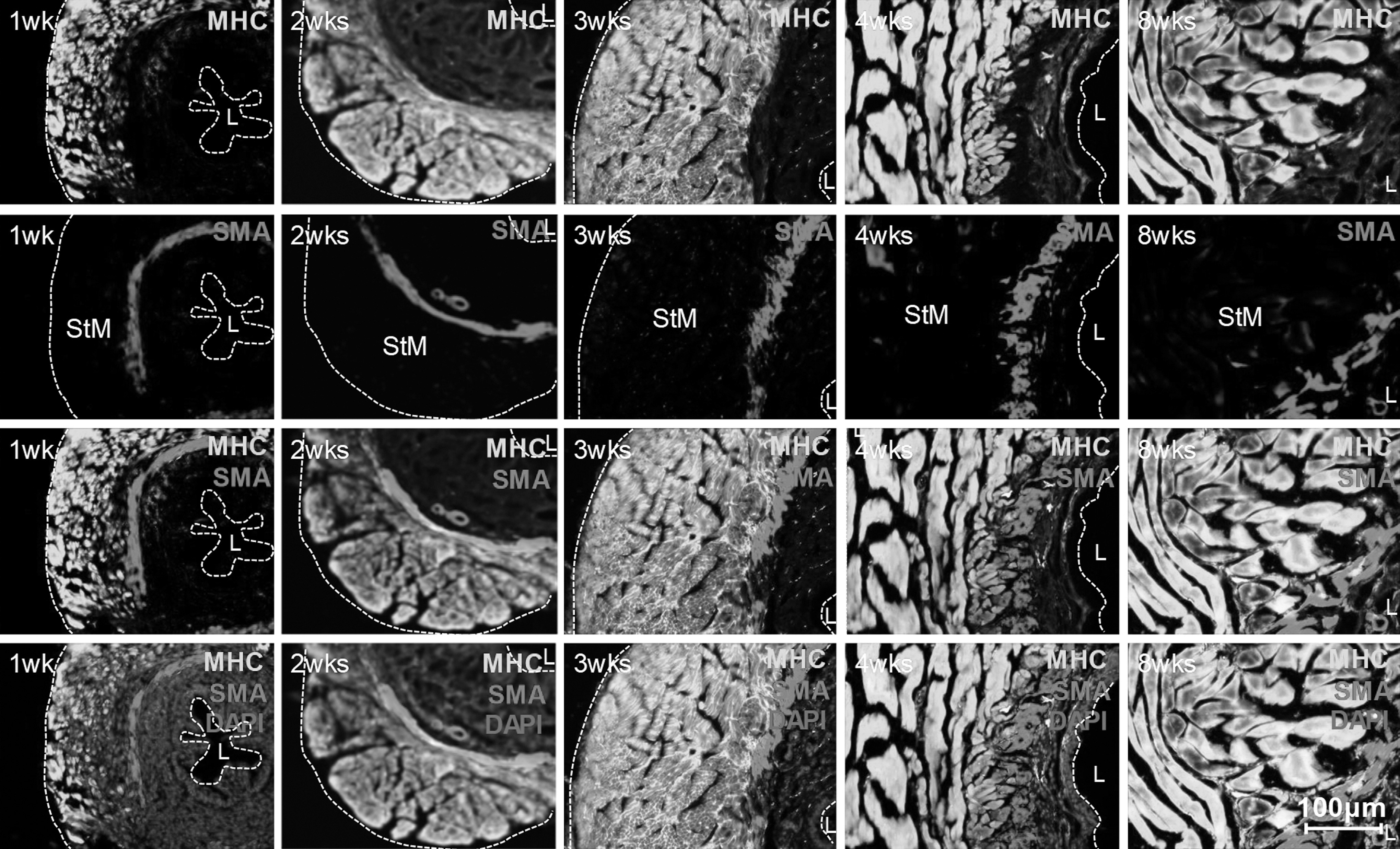
Detailed time-course development of STM. The STM layer remains underdeveloped at 1, 2, and 3 weeks. At 4 weeks, the structure becomes clear, and at 8 weeks it appears mature (original at × 200).
The ratio of the EUS length/total urethra in the rat was higher than reported in the human, but the rat had a similar ratio of bulbocavernosus muscle (Fig. 3A). The EUS was anatomically located between the bladder/prostate and bulbocavernosus muscle (Fig. 3B). After maturity, the smooth muscle layer crossed and merged with the inner longitudinal STM layer (Fig. 3C). The inner longitudinal STM layer and the outer circular STM layer differed from each other significantly by phalloidin and laminin IF staining (Fig. 3D).
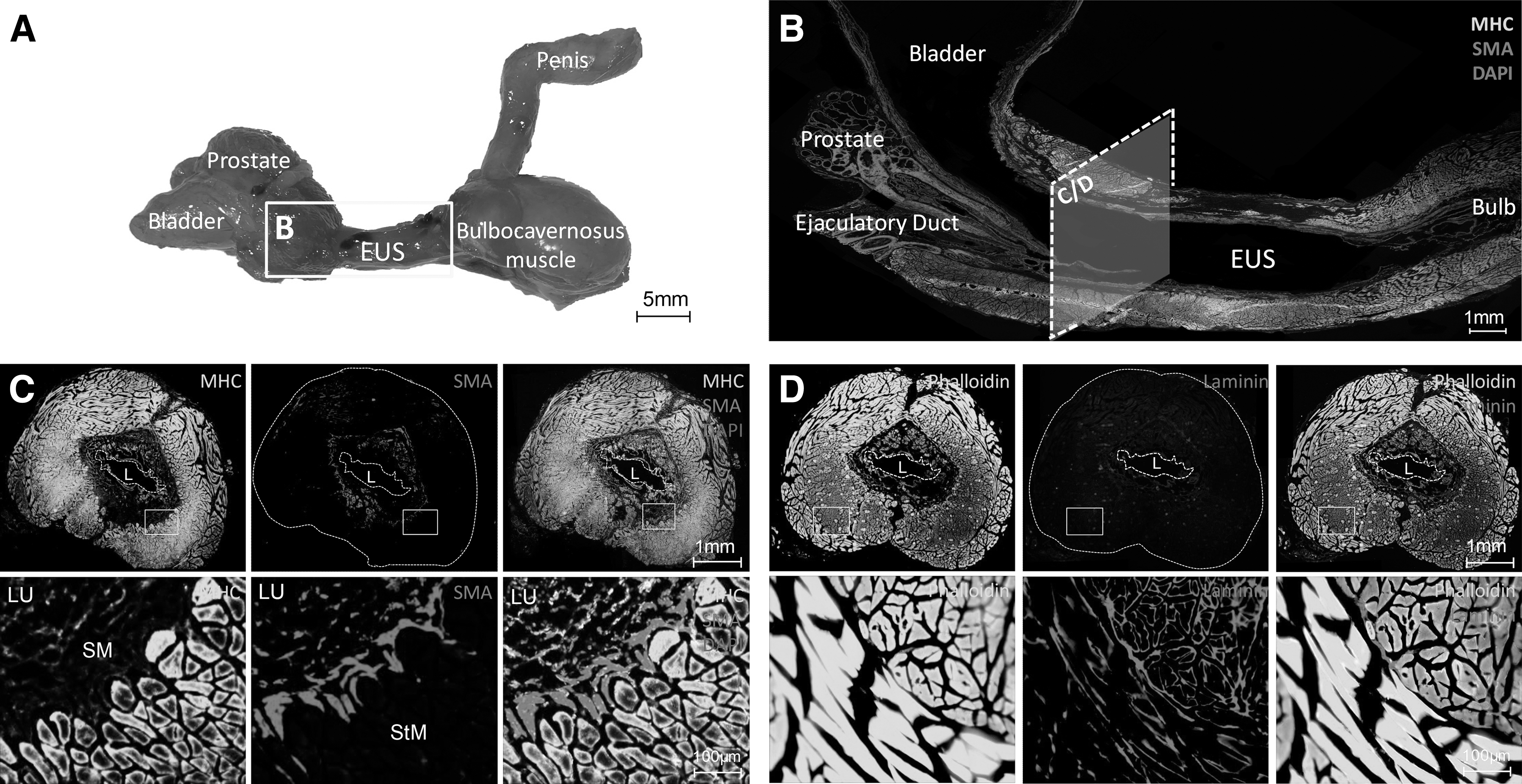
Anatomic structure of the male rat EUS.
Distribution of EdU-LRCs during development
The location and distribution of EdU LRCs were studied at different time points to describe the development of the EUS. To localize the resident stem/progenitor cells, cellular markers, including SMA, MHC, and laminin, were used to detect stem/progenitor cells at different time points. One-week-old rats had a higher rate of stem cell labeling (Fig. 4A), and the EdU-LRCs were distributed in every layer of the EUS. The number of EdU-LRCs decreased by time: 203 ± 24, 159 ± 17, 131 ± 21, 75 ± 12, and 46 ± 8 at 1, 2, 3, 4, and 8 weeks, with statistical significance compared with the previous time point (**P < 0.01) (Fig. 4B). At 8 weeks, long-term EdU retaining cells were present, and we defined these cells as EUS resident stem/progenitor cells [21]. The proportion of EdU-LRCs in the muscle layer, including the STM layer and smooth muscle layer, increased from 57.1% at 1 week to 89.1% at 8 weeks compared with the submucosal layer (from 42.9% at 1 week to 10.9% at 8 weeks) (Fig. 4C).

Distribution of EdU+ resident stem/progenitor cells during EUS development.
The EdU-LRCs in the STM and smooth muscle layers decreased at a slower rate than those in the submucosal layer. This indicates that EdU-labeled cells in connective tissue decrease faster than EdU-LRCs in muscle tissue (Fig. 5A). The EdU-LRCs were located immediately under the basal membrane of laminin and were adherent to the edge of the muscle fibers as shown in the schematic diagram (Fig. 5B, C).
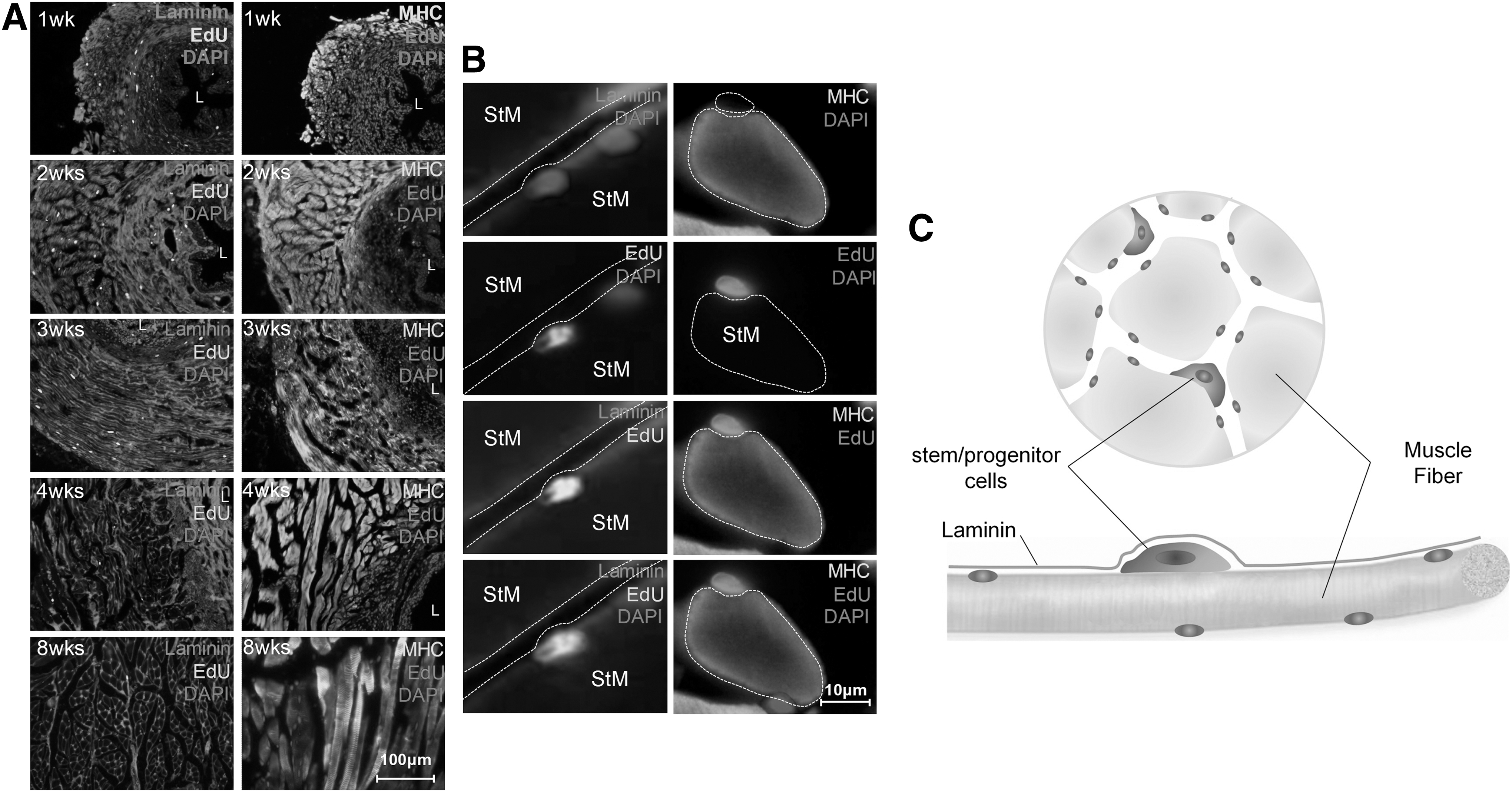
Location and proportion of stem/progenitor cells in different EUS layers.
Characterization of EUS resident stem/progenitor cells
To determine the ratio of proliferating cells during EUS development, expression of Ki67 and PCNA was assessed. These are markers of cellular proliferation and are expressed only in cycling cells (Fig. 6A and 7A). IF staining revealed that Ki67+ cells and PCNA+ cells decreased over time with statistical significance (**P < 0.01) (Fig. 6B and 7B). This decrease was similar to the decrease of EdU-LRCs (Fig. 6C and 7C), but the rate of decline in EdU-LRCs was faster. The ratio of EdU-LRCs to Ki67+ cells decreased over time, most notably at the time points of 4 weeks (**P < 0.01) and 8 weeks (*P < 0.05) (Fig. 6D). There were only a few colocalized EdU+Ki67+ cells. The ratio of EdU+Ki67+ cells to Ki67+ cells was 3.19%, 2.84%, 2.25%, 1.69%, and 1.15%, respectively, with statistical significance (**P < 0.01) (Fig. 6E).
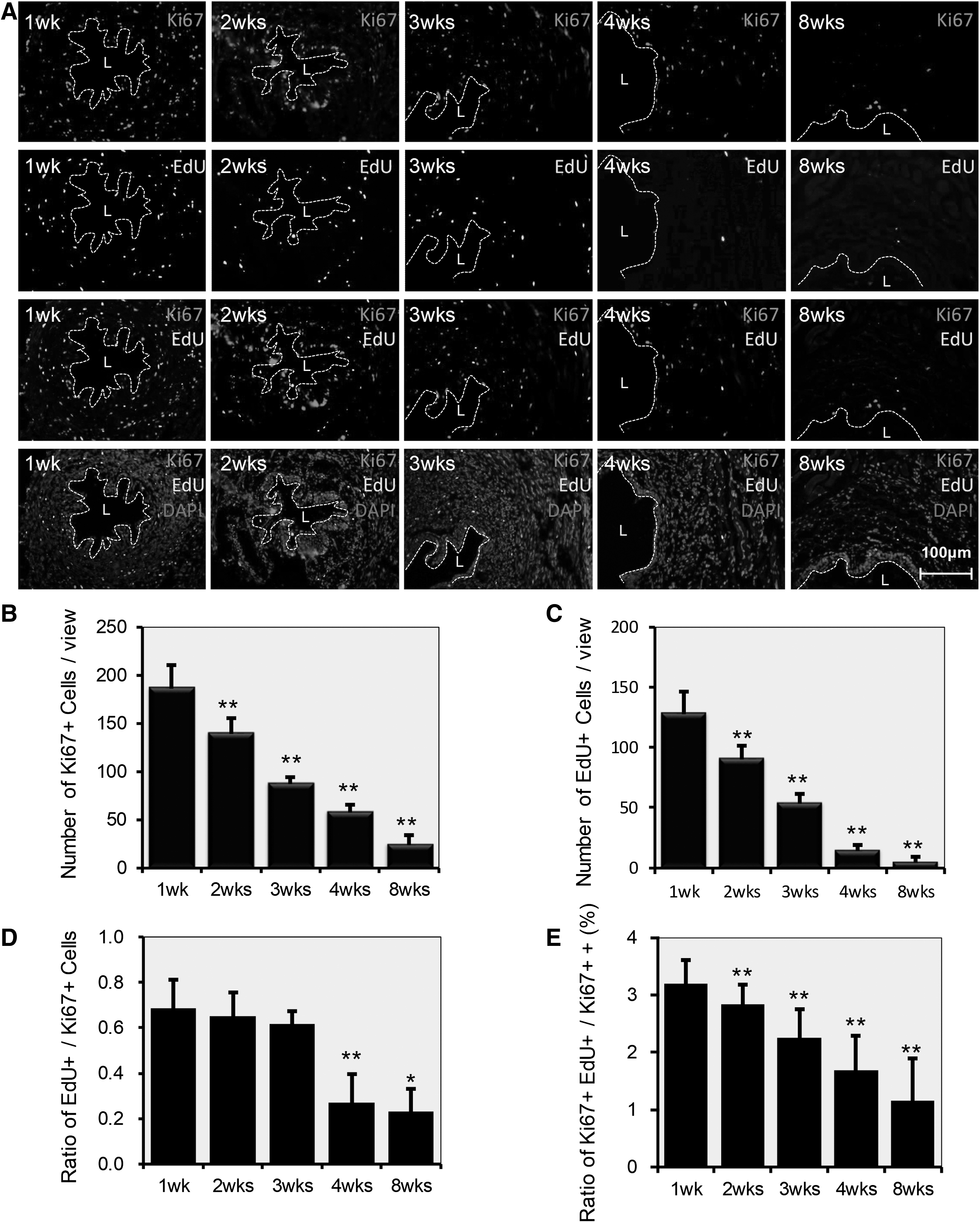
Expression of Ki67 as a characterization of EUS resident stem/progenitor cells.
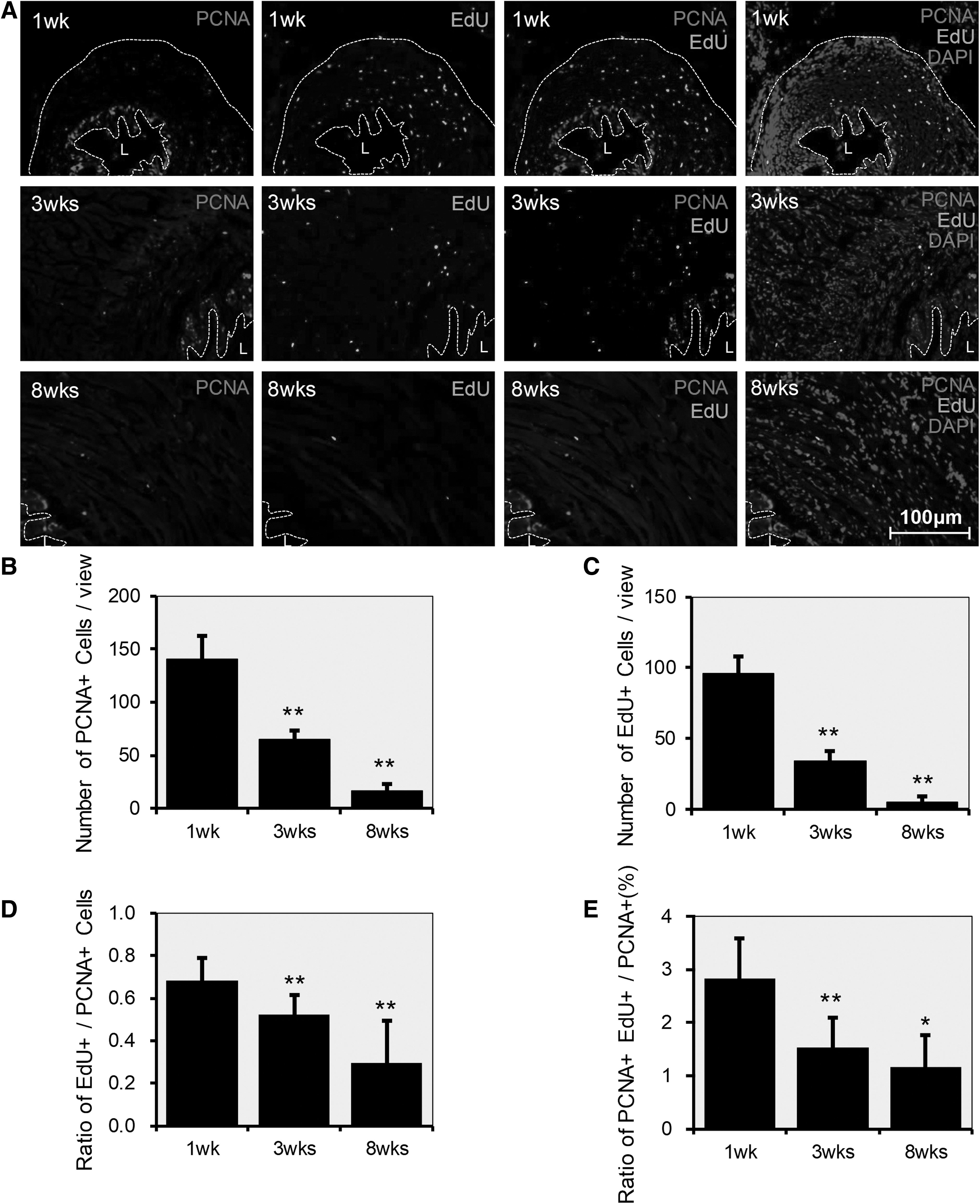
Expression of PCNA in the time-course development of the EUS.
The ratio of EdU-LRCs to PCNA+ cells also decreased by time, with statistical significance (**P < 0.01) (Fig. 7D). The colocalized EdU+ and PCNA+ cells decreased over time. The ratio of EdU+Ki67+ cells to Ki67+ cells was 2.84%, 1.54%, and 1.17% at 1, 3, and 8 weeks, with statistical significance (**P < 0.01, *P < 0.05) (Fig. 7E).
Isolation and myogenesis potential of male EUS resident stem/progenitor cells in vitro
We isolated EUS resident stem/progenitor cells from the 2-week-old rats using the MACS® MicroBeads system (CD34+ and integrin 7α+). The first passage pretended clonal growth, then the cells were induced to form myotubes in DMEM supplemented with 2% HS. Differentiated cells expressed MHC, which indicates that they could differentiate into myotubes (Fig. 8), whereas CD34-/integrin 7α- cells could not be induced into myotubes.

Isolation and myogenesis property of male urethral sphincter resident stem/progenitor cells in vitro.
Discussion
This study had several complementary objectives. First, we aimed to develop insight into the anatomic temporal patterns of development of the male rat EUS. Second, we sought to identify, localize, and describe the EdU+ resident stem/progenitor cells in the male rat EUS. Finally, we endeavored to sort out the relationship between EdU+ resident stem/progenitor cells and EUS development. No similar studies exist in the current literature. In 2006, Lehtoranta et al. reported the distribution of dense connective tissue and STM of the EUS along with the urodynamic recordings, electromyography, and longitudinal contraction force of the urethra in adult rats [22]. In 2001, researchers reported that the human EUS is recognizable early in fetal development as condensed mesenchyme and later as STM fibers [23]. Our study furthers this body of research by describing the sequential development of the male EUS and the location of the stem/progenitor cells.
Our results indicated that the STM layer of the EUS developed much faster than the smooth muscle layer. In addition, the different EUS layers were clearly differentiated by 4 weeks of age. These layers include the mucosal layer, submucosal layer, smooth muscle layer, inner longitudinal STM layer, and outer circular STM layer. With the detailed gross and IF images, this study provides a more complete description of the male rat EUS and further describes how its structure relates to its function for micturition and ejaculation. This information is particularly pertinent as most recent studies of the EUS investigate female SUI and very few studies examining the male EUS have been published [24,25].
Our findings suggest that the male EUS continues to develop and mature after birth and that the layers become more clearly defined with age. These findings may help to clarify the etiology of one very common urinary concern: pediatric enuresis. As children grow, the EUS matures and enuresis episodes gradually decrease as the EUS STM develops to control urinary continence.
Tissue-resident stem/progenitor cells have been extensively studied as a target of potential therapeutic intervention for regenerative medicine. Lin et al. recently reported the location of potential stem/progenitor cells in the penis [21]. Sallustio et al. reported that toll-like receptor 2 plays a role in the activation of human tissues-resident renal stem/progenitor cells and that it promotes adult renal progenitor/stem cells to differentiate into renal epithelial cells [26]. Najimi et al. reported that human liver mesenchymal stem/progenitor cells inhibit hepatic stellate cell activation thus preventing liver fibrosis [27]. With all these studies in mind, the activation of resident stem/progenitor cells presents a promising potential therapy for the cure of various chronic diseases.
In our study, the resident stem/progenitor cells were located in all layers of the male rat EUS. These cells decreased over time, but there were still remaining stem/progenitor cells even at 8 weeks. The proportion of resident stem/progenitor cells in the muscle layer increased from 57.1% at 1 week to 89.1% at 8 weeks compared with the submucosal layer (from 42.9% at 1 week to 10.9% at 8 weeks). Furthermore, the resident stem/progenitor cells in the mucosal layer and submucosal layer decreased faster than in the muscle layer. The total number of EUS resident stem/progenitor cells at the 8 weeks was ∼25% of the total at 1 week (46 ± 8/203 ± 24).
We corroborated the evidence of cell proliferation by assessing for expression of Ki67 and PCNA, which are markers of cell proliferation expressed only in cycling cells [28]. The exact location of EUS resident stem/progenitor cells was under the basal membrane of laminin, clinging to the edge of muscle fibers. We believe that these cells are colocated with muscle fibers as they can be activated by some specific signal or external stimulus for the repair of adjacent muscle fibers after injury. The number of Ki67+ and PCNA+ cells decreased over time, similar to the decrease in EdU+ resident stem/progenitor cells. At 4 weeks, the ratio of EdU+ cells to Ki67+ cells decreased sharply, and the structure of the different EUS layers clearly differentiated. This explains the development of the EUS, with increasing thickness of STM as the number of EdU+ resident stem/progenitor cells decreases. Even after maturity, tissue-resident stem/progenitor cells remain in the EUS, and these cells may provide a therapeutic target for those suffering from sphincteric dysfunction. Only 1%–3% of cells colocalized with EdU and Ki67/PCNA. This means that the EUS resident stem/progenitor cells occurred randomly, independent of the expression of Ki67 or PCNA.
Most animal studies of SUI have focused on female SUI with only a few studies investigating male SUI. Animal models of SUI include nerve crush, EUS injury by electrocauterization, transabdominal urethrolysis, pubourethral ligament transection, and female vaginal distention [29 –32]. Although this study focused on the anatomy of the male EUS and its stem cells in situ, we plan to utilize the techniques employed in this study to perform additional studies of male SUI in animal models.
Our study is unique in that it examined the postnatal development of the male EUS and defined the location and distribution of stem/progenitor cells in the male EUS. One limitation of our study is that we did not perform the study of the EUS development in the embryonic period. Another limitation is that we did not perform the research using a male rat incontinence model. A final limitation is that we did not study the feasibility of activating resident stem/progenitor cells either in vivo or in vitro. We are planning additional studies with these limitations in mind. We hope to be able to develop treatment modalities that can activate the resident stem/progenitor cells with potential clinical applications for postoperative SUI.
Conclusions
This study provides a detailed description of the anatomy of the male rat EUS and delineates the location and distribution of the resident stem/progenitor cells in the different layers of the EUS as it matures. This information provides a critical foundation for future studies of male micturition.
Footnotes
Disclaimer
Opinions, interpretations, conclusions, and recommendations are those of the authors and are not endorsed by the Department of Defense and do not represent the official views of the National Institutes of Health.
Author Disclosure Statement
T.F.L. is a consultant to Acoustic Wave Cell Therapy, Inc. All others have no conflict of interest.
Funding Information
Research reported in this publication was supported by NIDDK of the National Institutes of Health under award number R56DK105097 and 1R01DK105097-01A1. It was also supported by Army, Navy, NIH, Air Force, VA and Health Affairs to support the AFIRM II effort under award number W81XWH-13-2-0052. It was also supported by Jiangsu Government Scholarship for Overseas Studies (grant no. JS-2018-177).