
Abstract
Intrauterine adhesions (IUAs) are characterized by the injury of endometrium due to curettage and/or endometritis. The loss of functional endometrium in uterine cavity usually results in hypomenorrhea, amenorrhea, infertility, and/or recurrent pregnancy loss. Recently, stem cell transplantation has been applied to promote the endometrial regeneration. Human amnion epithelial cells (hAECs) have been shown to have stem cell characteristics. In this study, we found that PKH26-labeled hAECs were mainly distributed in the basal layer of endometrium after transplantation, and hAEC transplantation, including uterine injection and tail vein injection, could increase pregnancy rate and the number of embryos in rat model of IUAs. Moreover, hAEC transplantation was demonstrated to increase the endometrial thickness, promote the proliferation of glands and blood vessels, and decrease fibrotic areas in the endometrium. The immunohistochemical and quantitative polymerase chain reaction analysis showed the upregulated expression of growth factors, such as basic fibroblast growth factor (bFGF), vascular endothelial growth factor (VEGF), insulin-like growth factor-1 (IGF-1) after hAEC transplantation; and the downregulated expression of collagen type I alpha 1 (COL1A1), tissue inhibitor of metalloproteinase-1 (TIMP-1), and transforming growth factor-β (TGF-β), all of which are associated with the extracellular matrix (ECM) deposition after hAEC transplantation. The mRNA sequencing indicated that platelet-derived growth factor-C (PDGF-C), thrombospondin-1 (THBS1), connective tissue growth factor (CTGF), Wnt5a, and Snai2 were significantly modulated in treatment groups. These results indicate that hAEC transplantation promotes endometrial regeneration and the restoration of fertility in rat model of IUAs.
Introduction
Intrauterine adhesions (IUAs), also known as Asherman's syndrome, are a common factor contributing to infertility as a result of the damage to the basal layer of uterine endometrium [1]. In recent years, the increasing number of uterine operations such as myomectomy and curettage has lead to endometrium scar formation and the incidence of IUAs rises, followed by oligomenorrhea, amenorrhea, miscarriage, and/or infertility [2,3]. At the same time, infectious diseases are high-risk factors resulting in IUAs since the invasion of viruses and germs could induce fibrosis and inhibit the repair of endometrium itself [4,5].
Considering the injury of endometrium, various methods have been applied for treatment, such as hysteroscopic adhesiolysis, intrauterine device (IUD) placement, and estrogen administration after operation, which is recommended to prevent recurrence and assist the endometrium proliferate to restore uterine function [6 –9]. However, there are some shortcomings. For severe IUA patients, it is still a problem that they have a high recurrence rate and low pregnancy rate after operation [10].
The loss of stem cells in basal layer, which plays an important role in the regeneration of endometrium, may account for the poor recovery [11 –13]. Moreover, a new functional layer of endometrium can regenerate by the supplement of stem cells [14,15]. Stem cells have been supposed as candidate cells for biological therapy and tissue engineering for many years, and the supplement of stem cells is potential for rebuilding endometrial function. Mesenchymal stromal cells [16,17] and induced pluripotent stem cells (iPSCs) [18] have been used in the treatment of IUAs, as these cells could thicken endometrium and promote endometrial regeneration. However, there are still some difficulties in clinical practice, such as invasive collection and tumorigenicity.
Human amnion epithelial cells (hAECs) are obtained from the amnion and displayed the feature of multipotent stem cells, which could differentiate into all of the three germ layers (endoderm, ectoderm, and mesoderm) in vitro and vivo, considering as a candidate and ideal source of cell therapy [19,20]. At the same time, the procedure of cell isolation and cryopreservation has been proved to be reliable after a long-time research [21]. These cells have demonstrated a multidifferentiating ability [22,23] and considered as “immune privilege” [24 –26]. The isolation of cells from placenta poses few ethical concerns regarding the collection method and from which in excess of 100 million cells can be obtained [27]. Additionally, there had not been serious side effects reported, including tumor formation, which occurred after transplanting into volunteers' forearms for immunogenicity test [28].
Previous studies reported that hAECs had already been applied in several fields, such as lung and pulmonary fibrosis repair, wound repair, nerve system, and treatment of endothelial injury [29 –35]. However, the function of hAECs in IUAs has not been clarified.
All the above characteristics of hAECs suggest that they can be used as an excellent cell therapy. In this study, we aimed to observe whether the transplantation of hAECs promotes endometrial regeneration in rat model of IUAs and explore the underlying mechanism.
Materials and Methods
hAEC's isolation and culture
This procedure was approved by the Ethics Committee at The Second Affiliated Hospital of Chongqing Medical University and hAECs were isolated from the amnions of healthy women who provided written informed consent. Briefly, the tissue was washed in sterile Hank's Balanced Salt Solution (HBSS; Thermo Scientific, Waltham, MA) twice to discard blood cells. Then each amnion was cut into small pieces (2–3 cm), digested in 0.25% trypsin/ethylenediaminetetraacetic acid solution (Gibco, Grand Island, NY), and incubated for 30 min at 37°C in a water bath for two times. Complete culture medium (F12/Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (all from Gibco)) was added to inactivate the digestive solution. Cells were centrifuged for 10 min at 300g, and the cell pellets were suspended in complete culture medium for cell culture. Cells were seeded onto culture flasks. Postisolation cell viability was tested by the Trypan Blue exclusion assay. When the cells reached 80%–90% confluence, they were digested by 0.25% trypsin and resuspended in sterile HBSS at a concentration of 5 × 107 cells/mL for injection.
Immunofluorescence
Cells were washed with phosphate-buffered saline (PBS) twice and fixed with 4% paraformaldehyde (PFA) solution (Salarbio, Beijing, China) for 30 min on ice. Then, they were incubated in 0.2% Triton X-100 (Salarbio) for 30 min and blocked in 3% serum for 1 h at room temperature. Cells were incubated with primary antibodies against cytokeratin-18 (CK18, 1:200, ab668; Abcam, Cambridge, UK) or OCT4 (1:100, ab18976; Abcam) at 4°C for 12 h, and then with secondary antibodies (Boster, Wuhan, Hubei, China) for another 1 h at room temperature. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Boster).
Flow cytometry
Phenotypic analysis of cultured hAECs was performed by flow cytometry. The cell suspension with 5 × 105 cells was centrifuged at 1,500g for 5 min at 4°C. The cells were incubated with CK18, NANOG (Thermo Scientific), SSEA4, HLA-DQ, HLA-DR, CD324, CD146, CD34, CD45, CD29, and CD166 (BioLegend, San Diego, CA). All samples were run on a Beckman Coulter CytoFLEX-Analyzer (Beckman Coulter, Brea, CA) and data were analyzed by FlowJo software (TreeStar, Ashland, OR).
Establishment of IUA model and hAEC tracking
The experimental protocols and animal handling procedures were reviewed and approved by the Ethics Committee at The Second Affiliated Hospital of Chongqing Medical University. Female Sprague Dawley rats (8–10 weeks old, 220–260 g) were purchased from the Experimental Animal Center of Chongqing Medical University and housed at the temperature of 20–26°C, the humidity of 50%–60%, and a 12-h light/12-h dark cycle. At 8:00–10:00 am, vaginal smears were obtained daily for the check of estrus. Female rats were randomly assigned into four groups after two consecutive 4-day estrous cycles: (1) control group (n = 12), the uterine horns were only exposed in air for 30 min and then sutured; (2) model group (n = 12), received only mechanical injury without any treatment; (3) hAEC intrauterine injection group (n = 12), 1 week after mechanical injury, the uterine horns were exposed, inside each one of which, hAEC suspensions (5 × 106 cells in 100 μL PBS) were injected respectively, then sutured immediately; (4) hAECs tail vein injection group (n = 12), 1 week after mechanical injury, received hAEC suspensions (5 × 106 cells in 100 μL PBS) through the tail vein injection.
IUA models were established by mechanical injury. Under anesthesia with 5% pentobarbital sodium (5 mg/kg) injected intraperitoneally, the abdomen of rats was incised and the uterine horns were exposed. Two 0.1 × 0.1 cm incisions were clipped separately in the bottom of uteruses, and a 5 mm endometrial curette was used to scrape the upper two-third of the uteruses, respectively, until uterine walls became pale and rough. Both uterine horns of each rat were treated equally. After the exposure of uterine horns for 30 min, uterine and peritoneal cavities were flushed, the wounds were stitched, and the disinfection with iodophor solutions was performed.
To track the distribution of hAECs after transplantation, cells were stained with the PKH26 Red Fluorescent Cell Linker Kit (Sigma-Aldrich, St. Louis, MO) followed by the manufacturer's instructions [36]. One week after the establishment of IUA models, PKH26-labeled hAECs were injected into uterine horns or the tail vein. Two weeks and 4 weeks after hAEC transplantation, the uterine horns, spleen, lung, ovary, brain, liver, heart, and kidney were excised, fixed with optimal cutting temperature compound, and sliced into 10-μm sections. Subsequently, they were fixed in precooled acetone for 10 min and incubated with DAPI (Boster) for 10 min at room temperature. The sections were photographed under a fluorescent microscope (Nikon, Tokyo, Japan).
Cell counting kit-8 assay
The cell viability of PKH26-labeled hAECs was detected by the Cell Counting Kit-8 (CCK-8) (Beyotime, Haimen, Jiangsu, China) according to the manufacturer's instructions. Cells were seeded at a density of 5 × 103 cells/100 μL in 96-well plate and cultured in an incubator. The absorbance value at 450 nm was measured at 0, 24, 48, 72, and 96 h after culture, using a Microplate Reader (Nikon).
Histology examination
Two weeks after transplantation, uterine horns were fixed in 4% paraformaldehyde for 48 h. Tissues were dehydrated and embedded in paraffin, and 5 μm serial paraffin sections were stained with HE in a Tissue-Tek DRS 2000 (Sakura Finetek, Tokyo, Japan) according to the manufacturer's instructions. After Hematoxylin and Eosin (H&E) staining, the blood vessels and glands were counted under microscope (Nikon) by taking the mean value of four random high-power fields (HPFs). Endometrial thickness was measured by the vertical distance between serosa and luminal surface. Masson staining was performed according to a previously described method [37]. The fibrosis was evaluated by the fibrosis rate of four random HPFs. The fibrosis rate was calculated by the ratio of fibrotic areas to total endometrial areas (excluding the uterine cavity). Data were analyzed by Image-Pro Plus software (version 6.0).
Immunohistochemistry
The antibodies, including basic fibroblast growth factor (bFGF) (1:100; no. 36769; SAB, College Park, MD), vascular endothelial growth factor (VEGF) (1:100; no. 48707; SAB), tissue inhibitor of metalloproteinase-1 (TIMP-1) (1:200; bs-0415R; Bioss, Beijing, China), insulin-like growth factor-1 (IGF-1) (1:200; bs-4985R; Bioss), collagen type I alpha 1 (COL1A1) (1:200; bs-10423R; Bioss), and transforming growth factor-β (TGF-β) (1:100; no. 48569; SAB), were used for immunohistological staining according to a previously described method [37]. The percent of positive staining areas of random five HPFs were analyzed by Image-Pro Plus software (version 6.0).
RNA isolation and quantitative polymerase chain reaction
Total RNA was extracted using TRIzol Reagent (Invitrogen, Carlsbad, CA). The cDNA was synthesized using All-in-One First-Strand cDNA Synthesis Kit (GeneCopoeia, Guangzhou, Guangdong, China). All of the primers (Table 1) were synthesized by GeneCopoeia. The Real-Time PCR Kit was purchased from GeneCopoeia. Relative quantification of mRNA was performed using the comparative threshold cycle (CT) method. This value was used to plot the gene expression using the formula 2−ΔΔCT.
Primers of Specific Genes Used in Quantitative Polymerase Chain Reaction Analyses
bFGF, basic fibroblast growth factor; COL1A1, collagen type I alpha 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IGF-1, insulin-like growth factor-1; TGF-β, transforming growth factor-β; TIMP-1, tissue inhibitor of metalloproteinase-1; VEGF, vascular endothelial growth factor.
RNA sequencing
After concentration testing for total RNA of the uterus in control, model, and hAEC treatment groups, the mRNAs were abstracted from each sample using Oli-go(dT)-magnetic beads. Then they were subjected to terminal repair and addition of sequencing adapters, followed by agarose gel electrophoresis to recover the target fragment for PCR amplification. The complete library was sequenced with Illumina HiSeq X Ten PE150 sequencing strategy.
The sequence reads from transcriptome sequence was aligned to the Rattus norvegicus genome by HISAT2 with default parameters. The differences of gene expression in different samples were compared by calculated gene expression value. To further understand the biological functions, differentially expressed genes in each group were identified statistically to overrepresent by gene ontology (GO) terms and biological pathways. Kyoto Encyclopedia of Genes and Genomes Orthology Based Annotation System (KOBAS) was applied to identify significantly enriched pathways by using hypergeometric test and Benjamini–Hochberg false discovery rate (FDR) correction method with corrected P < 0.05.
Fertility evaluation
Two weeks after hAEC treatment, the female rats mated with healthy fertile male Sprague-Dawley rats at a ratio of 2:1. The day when a vaginal plug appeared was designated as day 0 of gestation. At day 15 of gestation, the female rats were euthanized. The embryos in uterine cavities were counted and photographed. The pregnancy rate was calculated by the ratio of pregnant rats to total rats.
Statistical analysis
All data were expressed as the mean and standard deviation and analyzed by using SPSS version 17.0 (SPSS, Chicago, IL). Student's t test or one-way analysis of variance (ANOVA) was used to analyze the data. A two-tailed P value of <0.05 was considered statistically significant.
Results
Characterization of hAECs
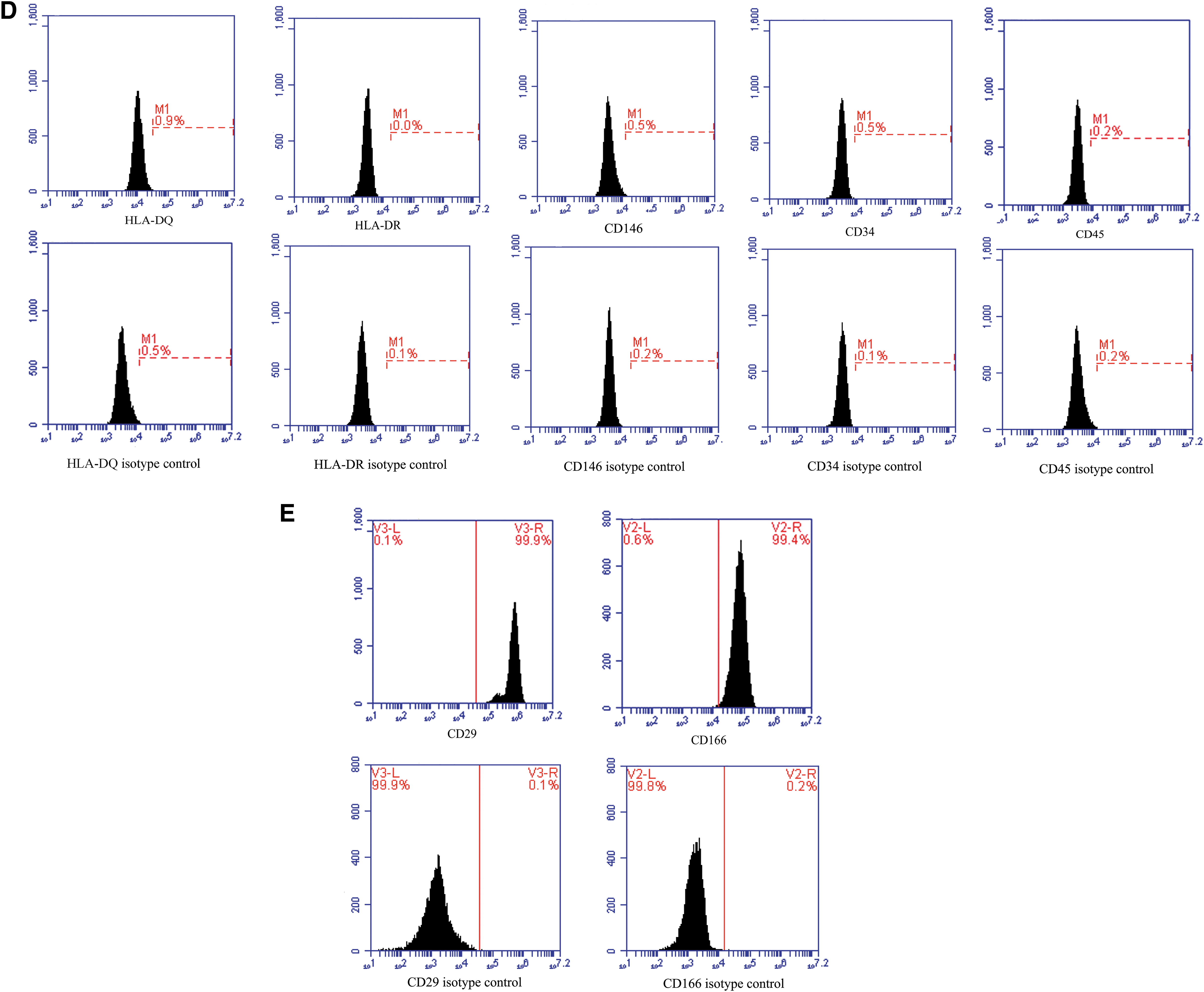
In our study, an average of ∼1.38 ± 0.25 × 108 hAECs could be yielded from each amnion. The hAECs in this study showed the cobblestone-like epithelial cell shape (Fig. 1A). The cells suggested positive expression of CK18 and CD324, which are epithelial markers (Fig. 1B). In addition, different abundance levels of pluripotent markers OCT4, SSEA4, and NANOG were detected in hAECs (Fig. 1C). The hAECs were negative for HLA-DR or HLA-DQ (Fig. 1D), revealing weak immunogenicity. The hAECs expressed CD29 and CD166, but did not express CD146, CD34, and CD45 (Fig. 1D, E).
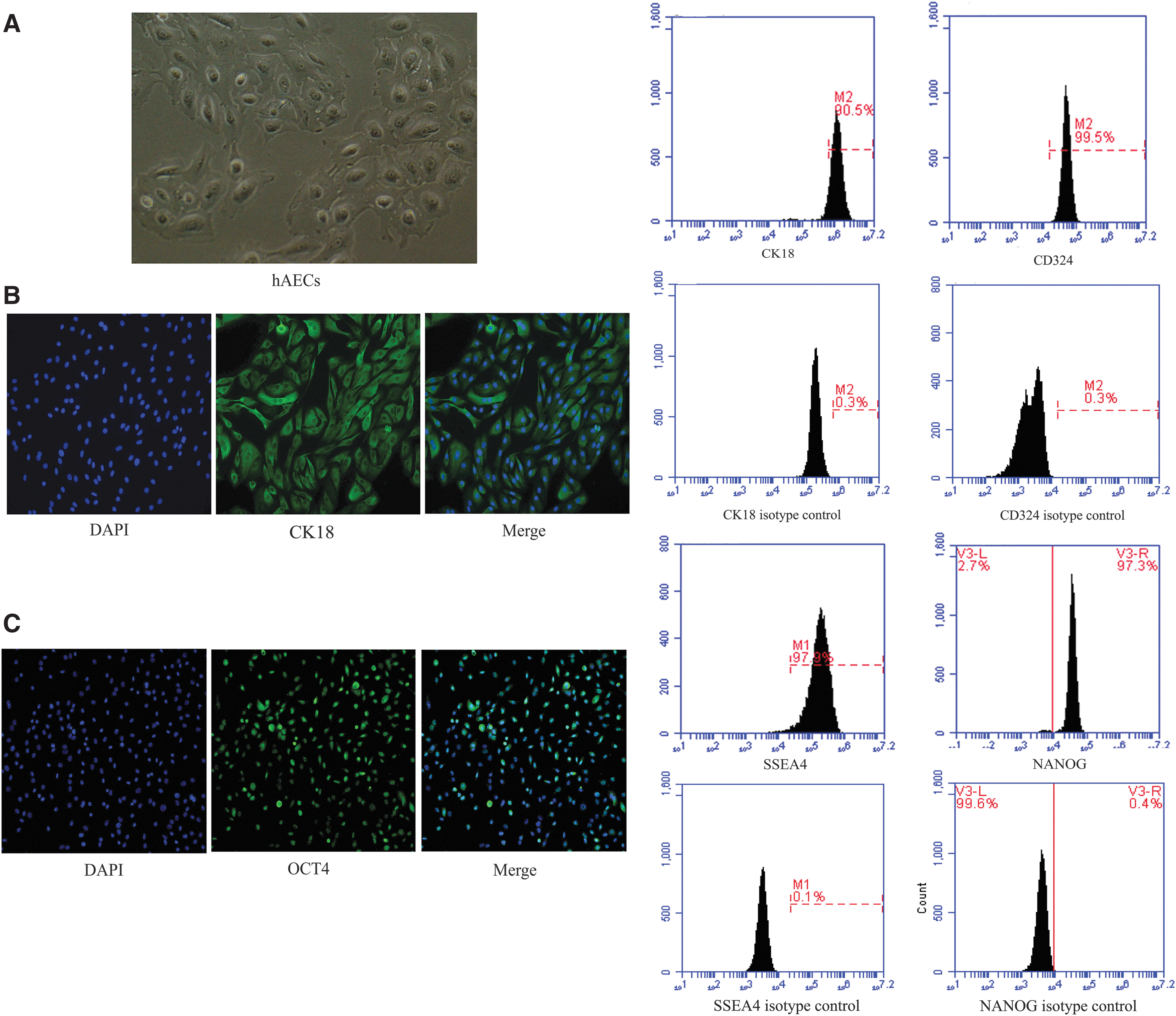
The biological characteristics of hAECs.
Establishment of models and location of hAECs after transplantation
H&E staining showed the discontinuous endometrium and decreased number of glands after uterine scratching (Fig. 2A, B; P < 0.001). Masson staining revealed significantly increased fibrotic areas in the endometrium of IUA models (Fig. 2C, D; P < 0.001). PKH26 was applied to label hAECs before transplantation (Fig. 3A). As assessed by flow cytometry, the rate of PKH26-labeling cells was 99.52% ± 0.35% (Fig. 3B), which did not reduce after cell passaging (99.19% ± 0.26%; Fig. 3C). There was no significant difference in cell proliferation between PKH26-labeled and unlabeled hAECs (Fig. 3D). These findings suggested that PKH26 labeling is stable and efficient, without affecting the viability of hAECs. Two weeks and 4 weeks after cell transplantation, PKH26-labeled hAECs (the red fluorescent signal) were mainly located in the basal layer of endometrium in both uterine injection group (2 weeks, 38.7%; 4 weeks, 18.3%) and tail vein injection group (2 weeks, 33.4%; 4 weeks, 16.6%) (Fig. 3E). Comparatively, a little fluorescent signal was also seen in spleen (uterine injection group, 4.35%; tail vein injection group, 3.86%), lung (uterine injection group, 3.52%; tail vein injection group, 3.48%), and ovary (uterine injection group, 3.60%; tail vein injection group, 3.27%), whereas the signal was rarely observed in other organs, including brain (0%), liver (0%), heart (0%), and kidney (0%) (Supplementary Figs. S1 and S2).
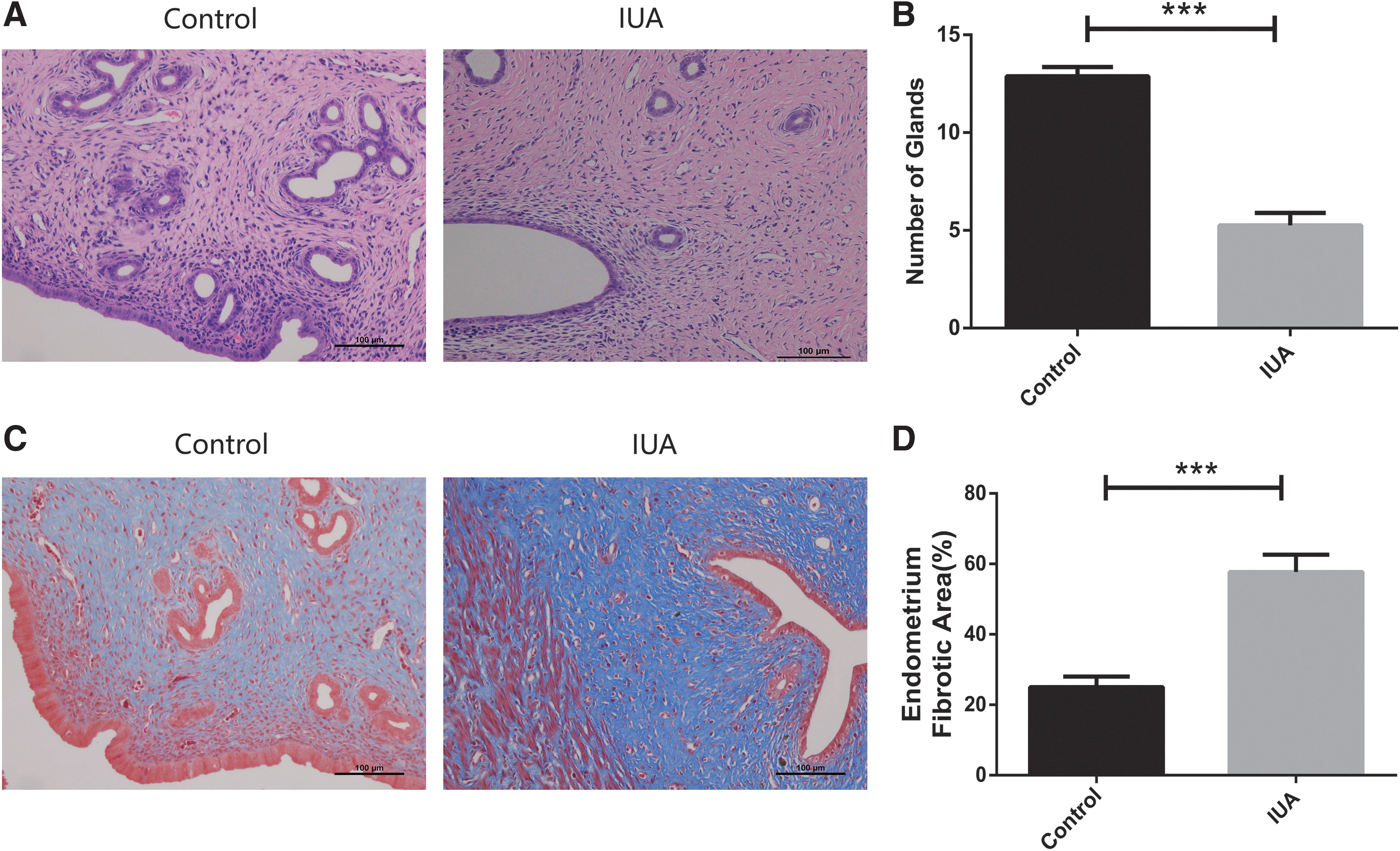
Establishment of IUA rat models.
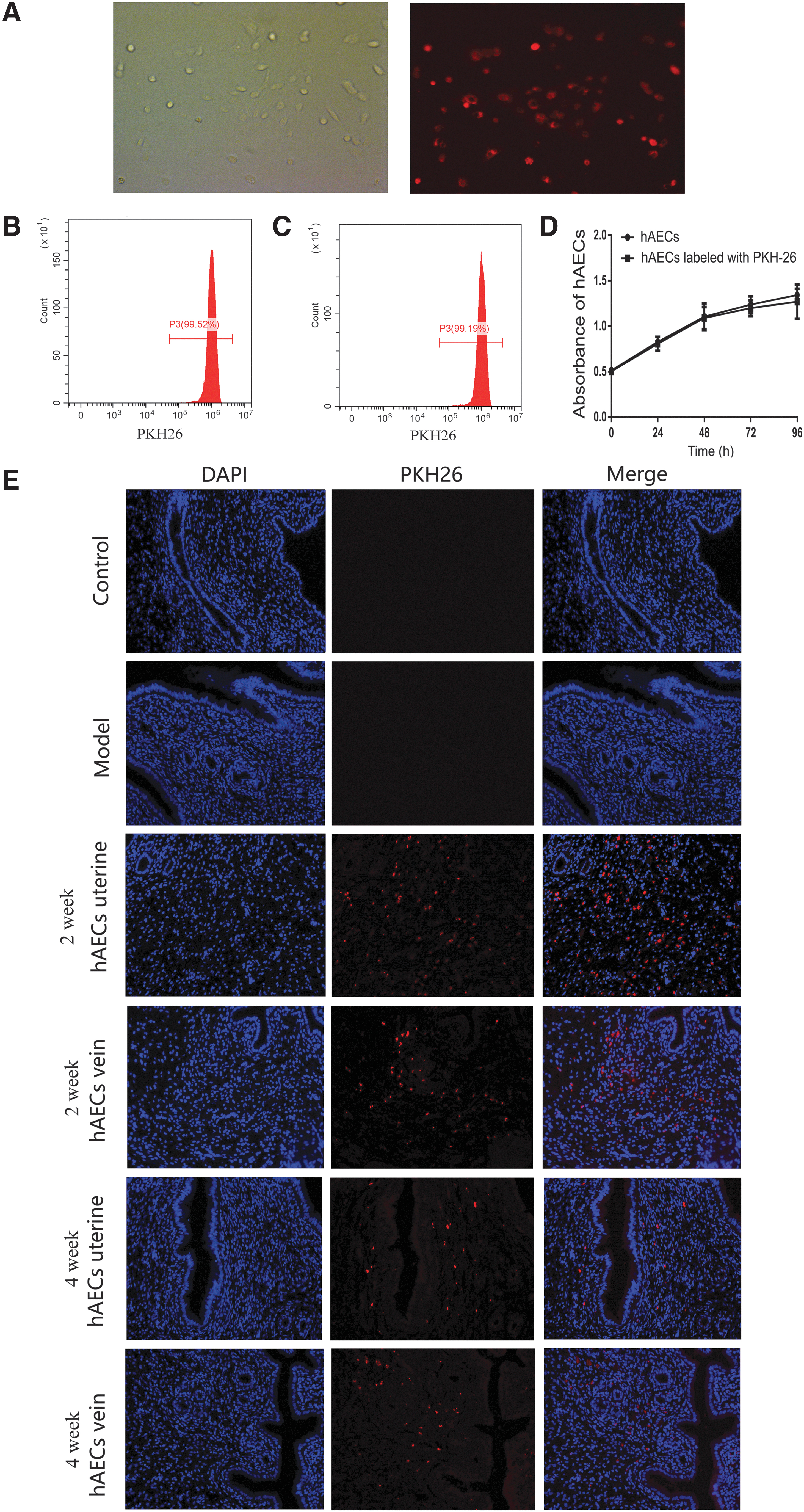
The hAECs were located in the basal layer of endometrium after transplantation.
Pregnancy outcomes after hAEC transplantation
The average number of embryos in the uterine injection group (9.5), tail vein injection group (9), and control group (12.5) was significantly higher compared with the model group (2.8) (Fig. 4A, B; P < 0.000 1). Besides, the pregnant rate in both the uterine injection (83.33%) and tail vein injection groups (83.33%) was significantly higher compared with the model group (50%), whereas all the rats in the control group were pregnant (Table 2).
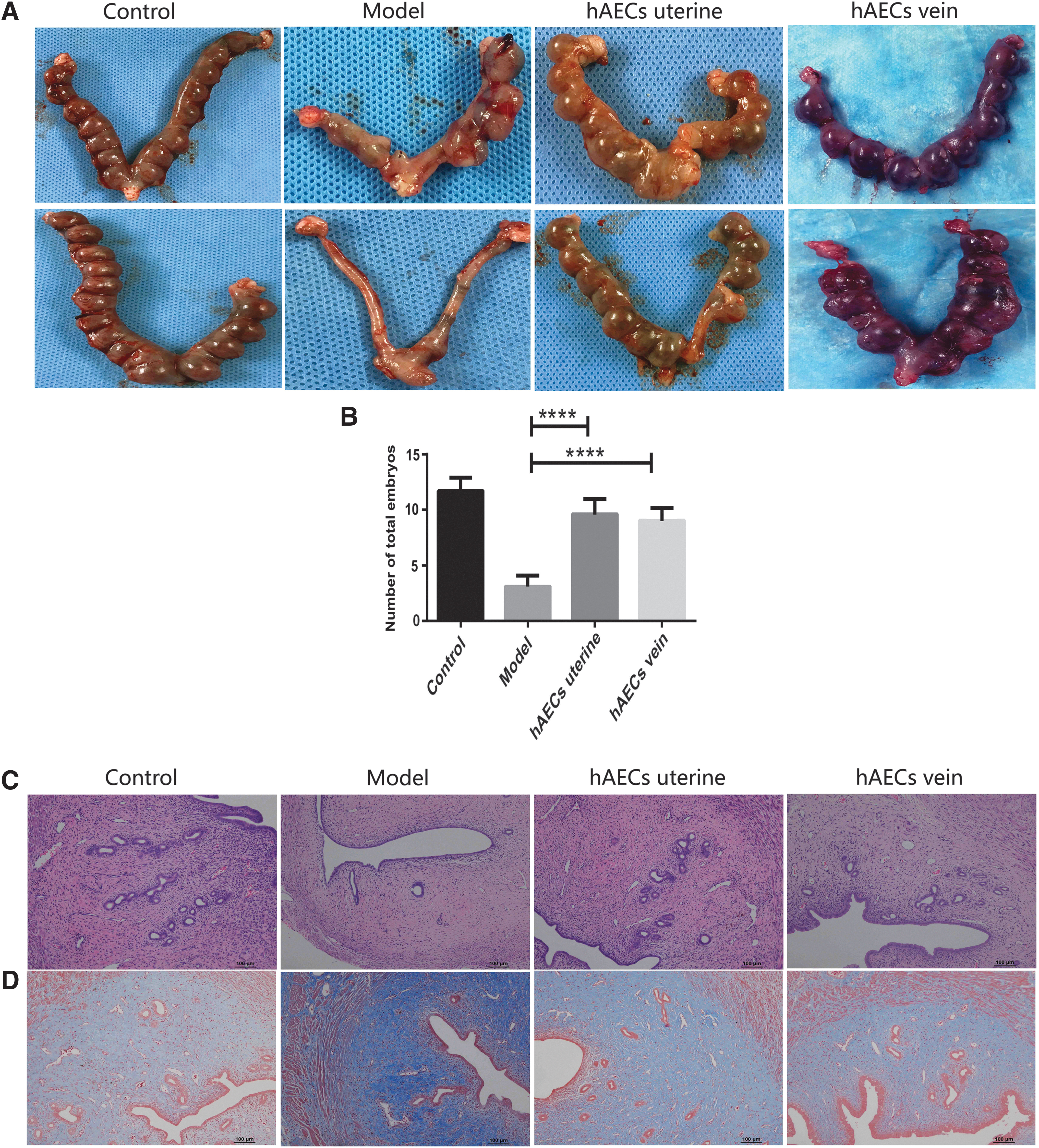
Pregnancy outcome and histological changes of endometrium after hAEC transplantation.
Pregnancy Outcomes Among Different Groups
Treatment with hAECs promote endometrial regeneration
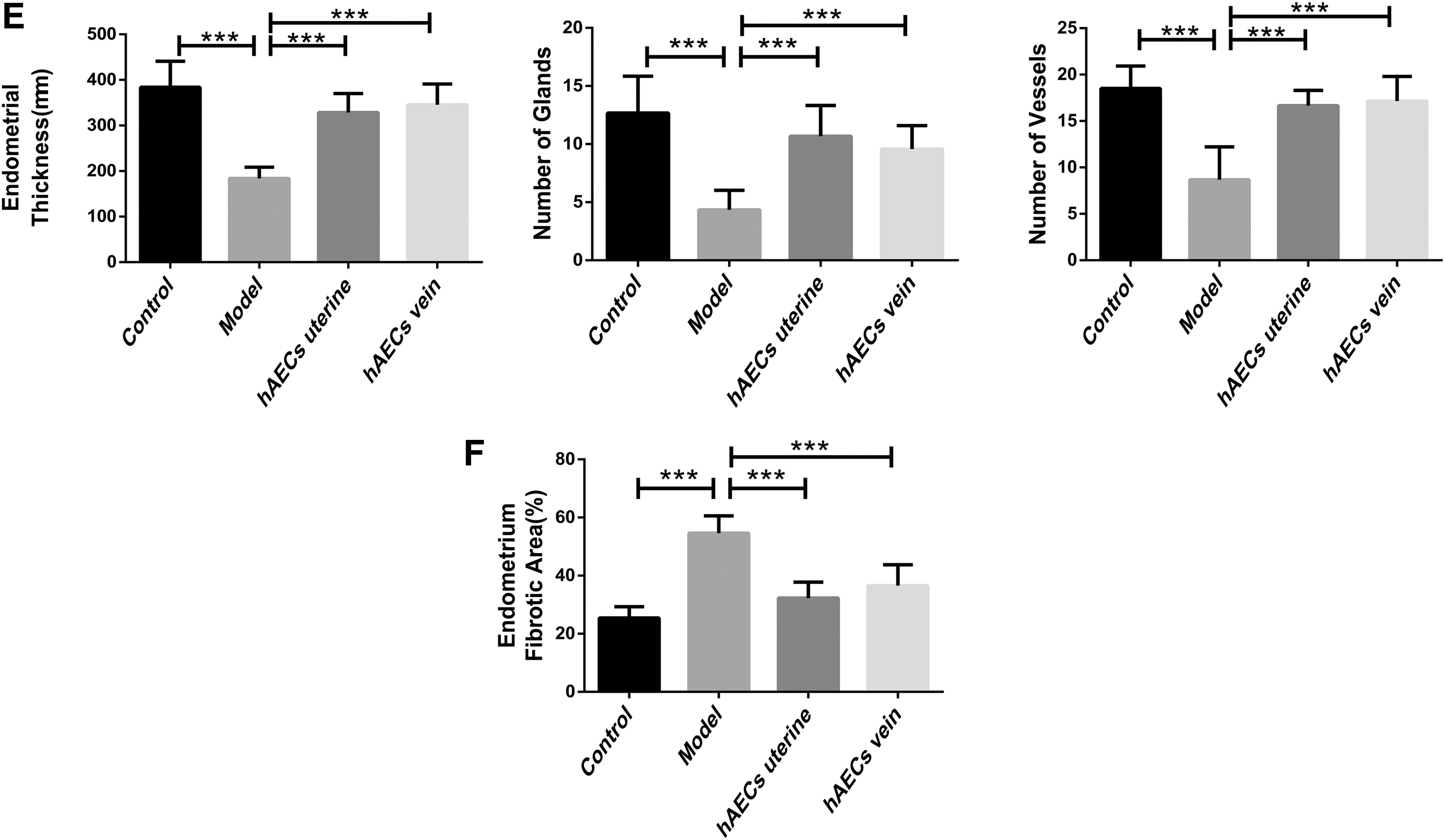
H&E staining revealed decreased endometrial thickness, blood vessel number, and gland number in the model group compared with the control group. However, the endometrial thickness, blood vessel number, and gland number were significantly higher in both uterine injection group and tail vein injection group, when compared with that in model group after 2 weeks of transplantation (Fig. 4C, E; P < 0.001). Moreover, there was no significant difference in these parameters between uterine injection group and tail vein injection group (Fig. 4C, E; P = 0.444). Masson staining results validated that fibrotic areas in the model group were significantly increased compared with that in control group. The fibrotic areas were remarkably decreased in both hAEC injection groups compared with that in the model group (Fig. 4D, F; P < 0.001). However, there was no significant difference in fibrotic areas between uterine injection group and tail vein injection group (Fig. 4D, F; P = 0.276).
The immunohistochemistry (IHC) and quantitative polymerase chain reaction (qPCR) analysis showed that the expression of bFGF, VEGF, and IGF-1 significantly increased in both uterine injection group and tail vein injection group compared with that in the model group and control group (Figs. 5A and 6A; P < 0.05). The IHC analysis revealed that the expression of bFGF, VEGF, and IGF-1 was lower in model group compared with the control group (Fig. 5A, B; P < 0.05). However, the qPCR analysis showed that the expression of IGF-1 was lower in model group compared with the control group (Fig. 6A; P < 0.05), and there was no significant difference in the expression of bFGF and VEGF between model group and control group (Fig. 6A; P = 0.092 and P = 0.119, respectively). As for extracellular matrix (ECM) deposition-related cytokines, COL1A1, TIMP-1, and TGF-β, the IHC and qPCR analysis showed that the expression level of these cytokines increased apparently in the model group when compared with the control group (Figs. 5C and 6B; P < 0.05), whereas transplanted hAECs significantly downregulated the expression of these cytokines compared with model group (Figs. 5C and 6B; P < 0.05). The IHC analysis revealed that the expression of COL1A1, TIMP-1, and TGF-β in hAEC treatment groups was higher compared with the control group (Fig. 5C, D; P < 0.05), whereas the qPCR analysis showed that there was no significant difference in the expression of COL1A1, TIMP-1, and TGF-β between hAEC treatment group and control group (Fig. 6B; uterine injection group vs. control group P = 0.075, P = 0.093 and P = 0.139, respectively; tail vein injection group vs. control P = 0.082, P = 0.107 and P = 0.146, respectively;). Moreover, there was no significant difference in the expression level of these cytokines between uterine injection group and tail vein injection group (Fig. 6A, B; P = 0.941, P = 0.077, P = 0.428, P = 0.488, P = 0.875 and P = 0.386, respectively).

The immunohistochemical assay of cytokines. The expression levels of growth factors (bFGF, VEGF, and IGF-1) and ECM deposition-related cytokines (COL1A1, TIMP-1, and TGF-β) were measured by IHC after hAEC transplantation.

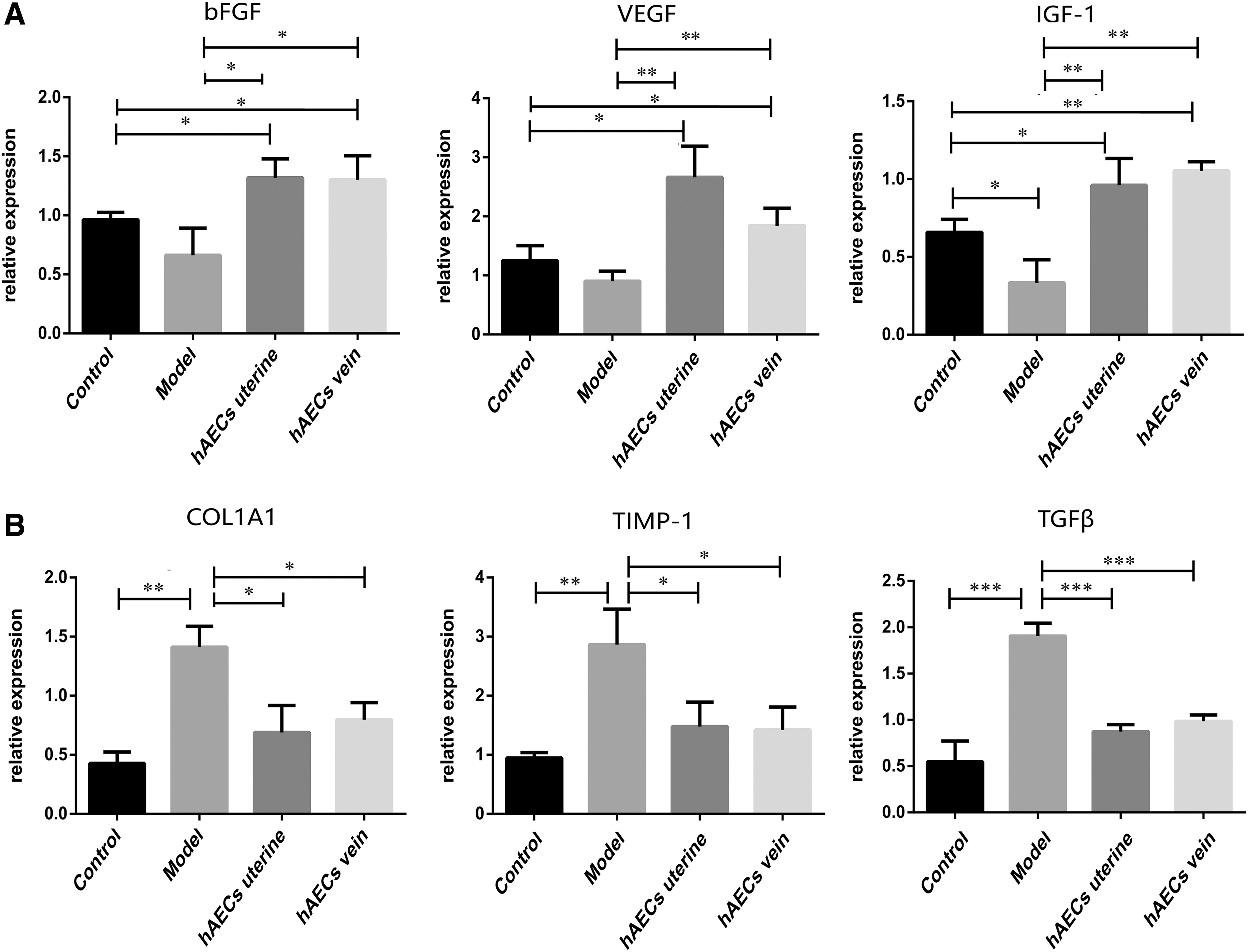
The qPCR assay of cytokines. The mRNA levels of growth factors (bFGF, VEGF, and IGF-1) and ECM deposition-related cytokines (COL1A1, TIMP-1, and TGF-β) were measured by qPCR after hAEC transplantation.
Differentially expressed mRNA screening and qPCR validation
Totally, the expression of 1, 035 mRNAs was found to be different between hAEC treatment group and model group, including 600 genes that were upregulated and 435 genes downregulated. Moreover, the expression of 53 mRNAs was found to be different between control group and model group, including 38 genes that were upregulated and 15 genes downregulated (Fig. 7A). The differentially expressed genes between hAEC treatment group and model group were mainly enriched for PI3K-Akt, Wnt, Chemokine, cAMP, protein processing in endoplasmic reticulum, and cytokine-mediated signaling pathways. Five differentially expressed genes were selected to be validated by qPCR, including platelet-derived growth factor-C (PDGF-C), thrombospondin-1 (THBS1), and connective tissue growth factor (CTGF) of PI3K-Akt signaling pathway, and Wnt5a and Snai2 of Wnt signaling pathways. Consistent with the result of mRNA sequencing, hAEC transplantation significantly downregulated the expression of PDGF-C, THBS1, and CTGF, whereas significantly upregulated the expression of Wnt5a and Snai2 (Fig. 7B; P < 0.05).
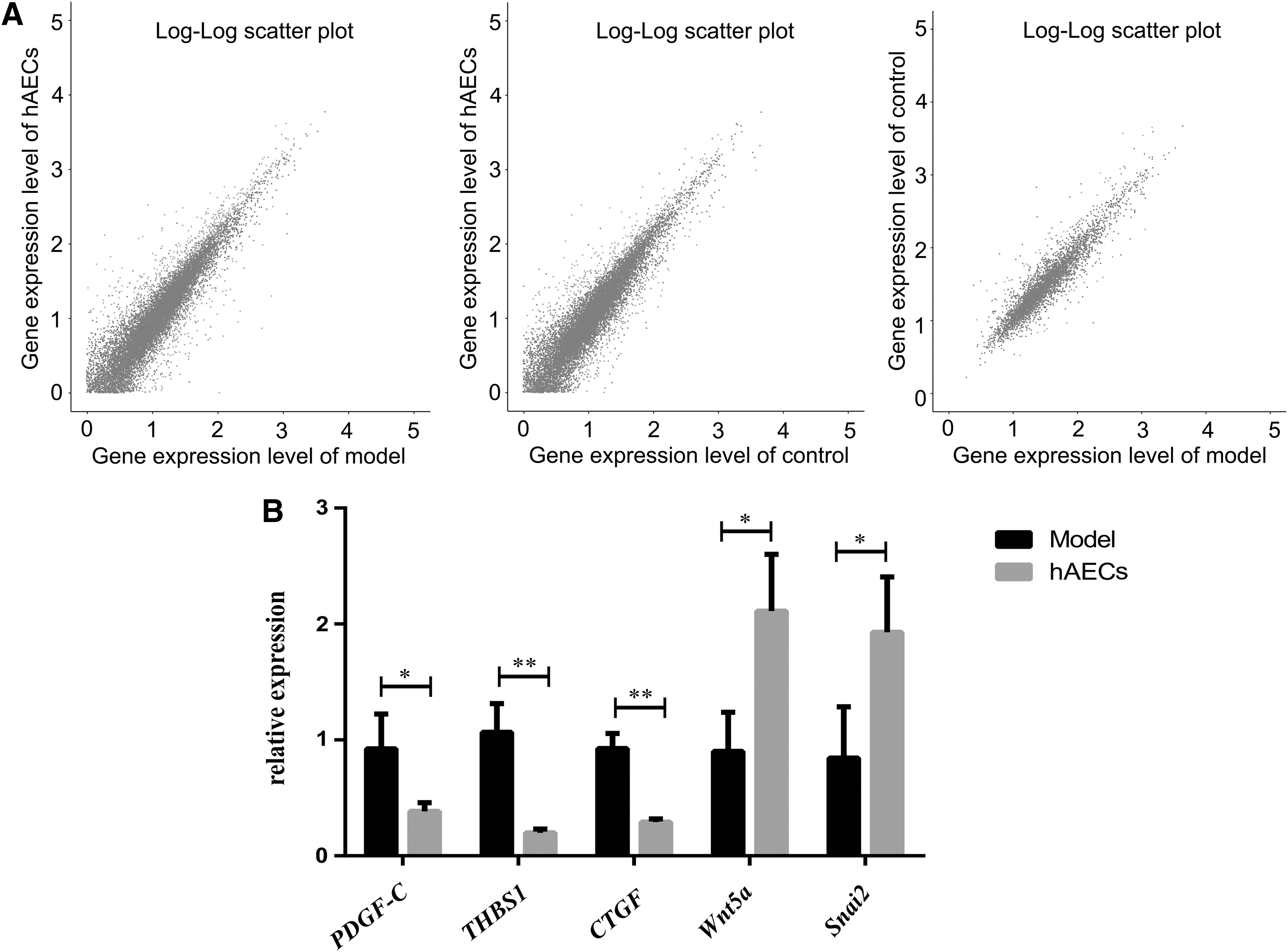
The mRNA sequencing and qPCR validation.
Discussion
In the present study, we aimed to evaluate whether the transplantation of hAECs improves the reproductive performance of IUA rat model. The hAEC transplantation showed positive effects on endometrial regeneration, no matter through uterine or tail vein injection. The endometrial thickness, blood vessels, and glands increased, and the fibrotic areas decreased after hAEC treatment. Moreover, we demonstrated that hAEC transplantation can facilitate the embryo implantation in rat model of IUAs. In addition, we revealed that hAEC transplantation could upregulate the expression of growth factors (bFGF, VEGF, and IGF-1) and downregulate the expression of ECM deposition-related cytokines (COL1A1, TIMP-1, and TGF-β) in uterine tissue.
Endometrium is a highly vascularized, dynamic, and cyclically regenerating tissue. The histopathological feature of IUAs is usually characterized by avascular or thin-walled telangiectatic vessels. It has been suggested that angiogenesis is crucial for the endometrial regeneration [38]. The growth factors, including bFGF and VEGF, play an important role in angiogenesis, epithelial and stromal development, and maintaining the normal function of blood vessels in endometrium [39 –41]. Additionally, a collagen-binding bFGF delivery system had been demonstrated to improve the regenerating ability of uterine endometrium and vascularization in rat model with severe endometrial damage [42]. VEGF is critical for vascular regeneration and re-epithelialization of endometrium [43 –47]. In our study, we observed the increased vascularization after hAEC transplantation in the endometrium of IUD rats by H&E staining. Consistent with the results of previous studies, we demonstrated that the expression of bFGF and VEGF was upregulated by IHC and qPCR assay. Therefore, we speculated that hAECs could promote the recovery of injured endometrium by promoting the expression of bFGF and VEGF and then the angiogenic effect. IGF-1 is a key mediator of tissue oxygen, wound healing, growth-promoting action, and has important effects on cell proliferation, differentiation, and metabolism [48 –51]. The proliferative and differentiative effects of IGFs on human uterine endometrium are thought to support the growth, regression, and regeneration of uterine tissue during the menstrual cycle [52,53]. Therefore, the increased expression of IGF-1 after hAEC treatment might partly account for the proliferation of glands and tissue repair in the endometrium.
Endometrial fibrosis is the main pathological feature of IUAs [54]. Massive ECM produced by stromal cells accumulating in the injured endometrium leads to the failure of functional endometrium to regenerate [55]. A substantial change in ECM deposition is the excessive production of collagens, for example, type I collagen, which is coded by two genes, COL1A1 and COL1A2 [56,57]. TIMPs are central molecules in tissue fibrosis, playing a pivotal role in progressive fibrosis and fibrolysis [58,59]. The latest evidence suggests that the collagen turnover and remodeling of ECM is mainly regulated by matrix metalloproteinases and their inhibitors, TIMPs (TIMP-1 and TIMP-2) which could inhibit ECM degradation [60 –63]. TGF-β is a central mediator of profibrosis and regulator of the ECM, playing a critical role in the development of fibrogenesis and organ dysfunction in a number of diseases, such as renal diseases and pulmonary fibrotic diseases [64 –67]. Experimental and clinical data have shown that TGF-β plays a leading role in the repair process after gynecological operations and is involved in the pathogenesis of postsurgical adhesions [68 –70]. In our study, we demonstrated that the fibrosis increased in the endometrium of rat model and hAEC transplantation inhibited the ECM deposition probably by downregulating the expression of COL1A1, TIMP-1, and TGF-β.
After hAEC transplantation, five differentially expressed genes were identified through mRNA sequencing. PDGF-C, a member of the PDGF ligand family, plays a critical role in the regulation of fibrogenesis [71,72]. The latest studies suggest that the overexpression of PDGF-C results in tissue fibrosis and ECM deposition by inducing a variety of profibrotic genes and related pathways, indicating that PDGF-C may act as an initiator of fibrosis [73 –75]. THBS1, a homeotrimeric ECM glycoprotein, has been reported to be a main endogenous activator of TGF-β in vivo and identified as a profibrogenic cytokine that regulates the expression of fibrillar collagen [76 –79]. In addition, THBS1 also has been found to inhibit cell proliferation and angiogenesis in the human endometrium [80 –83]. CTGF is a multifunctional matricellular protein, which is induced by TGF-β and considered as a downstream mediator of the profibrogenic effect of TGF-β [84]. CTGF is overexpressed in endometrial tissue with IUAs and is considered as an effective therapeutic indicator against IUAs [85]. In our study, hAEC transplantation led to decreased ECM deposition and the decreased expression of PDGF-C, THBS1, and CTGF. It can be speculated that hAECs decrease deposition collagen probably by downregulating the expression of PDGF-C, THBS1, and CTGF. However, the specific mechanism still requires further exploration. Wnt5a, a member of Wnt cytokine family, plays an important role in regulating many key developmental steps, such as embryo development, cell growth, and tissue regeneration [86,87]. Wnt5a is also necessary for epithelial differentiation and development of endometrial glands [88]. The proper level of Wnt5a is vital for early pregnancy events, which contribute to crypt formation for blastocyst attachment [89]. Snai2, a transcription factor of Snai family, regulates the epithelial/mesenchymal transition-like process during embryogenesis and wound healing, and accelerates reepithelialization in vitro [90 –92]. Besides, Snai2 has been found to express highly in glandular epithelium of endometrium around the implantation window in mice, so it may play a key role in embryo implantation [93,94]. In this study, hAEC transplantation led to the increased pregnancy rate and the increased expression of Wnt5a and Snai2. It can be speculated that hAECs promote embryo implantation by secreting Wnt5a and Snai2, both of which are critical to this important physiological process of pregnancy; however, the specific mechanism remains to be studied.
Conclusions
In conclusion, this study found that hAEC transplantation increased pregnancy rate in rat model of IUAs, which was probably attributed to a thicker endometrium, increased number of glands and blood vessels, and decreased fibrotic areas in endometrium, which might be the results of the upregulated expression of growth factors and the downregulated expression of ECM deposition-related factors. Further clinical studies are needed to examine the efficacy of hAEC transplantation in patients with IUAs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the grants from the Health Commission of Chongqing (nos. 2017MSXM037 and ZY201702077).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.