
Abstract
Stem cell-based therapy to restore the function of abnormal trabecular meshwork (TM) and decrease intraocular pressure (IOP) provides a novel approach to treat open-angle glaucoma. However, molecular mechanism for stem cells homing and anchoring to the TM remains unclear. This study aimed to discover the function of integrins in homing and integration of exogenous TM stem cells (TMSCs) to the TM. Integrin expression in TMSCs and fibroblasts was evaluated by quantitative real-time PCR (qPCR), flow cytometry, immunofluorescent staining, and western blotting. Expression of integrin ligand fibronectin was detected in cultured TM cells and murine TM tissue by immunostaining. Cell affinity to TM cells or fibronectin matrix was examined to compare TMSCs with TMSCs functionally blocked with an α5β1 integrin antibody. TMSCs and TMSCs with α5β1 integrin-blocking were intracamerally injected into wild-type mice. Wholemounts and cryosections were analyzed to discover cell distribution and integration at 3 days and 1 month. IOP was measured to detect possible changes. We discovered that human TMSCs expressed a higher level of α5β1 integrin than fibroblasts, but similar levels of αvβ3 and αvβ5 integrin. Upregulation of fibronectin was found in both TM cells treated with dexamethasone for 14 days and murine TM tissues damaged by laser photocoagulation. TMSCs were able to attach to the TM cells and fibronectin matrix in vitro. When the surface α5β1 integrin was blocked, the attached cell numbers were significantly reduced. Both TMSCs and TMSCs incubated with an α5β1 integrin-blocking antibody could home to the mouse TM after injection. TMSCs blocked with the α5β1 integrin-blocking antibody were not retained in the TM tissue at 1 month. The injected cells did not affect mouse IOP. In conclusion, highly expressed α5β1 integrin participates in maintaining TMSCs anchored and integrated to the TM, which would be crucial for stem cell-based therapy for glaucoma.
Introduction
Glaucoma is a group of diseases that causes damage to the optic nerve and leads to irreversible vision loss [1]. Although the exact pathogenesis of glaucoma remains unclear, it is widely accepted that elevated intraocular pressure (IOP) can trigger the upregulation of downstream molecules and induce glutamate excitotoxicity and oxidative stress to deteriorate the optic nerve [2 –4]. Conventional treatments for glaucoma, whether pharmaceutical, laser, or surgical, focus on decreasing aqueous humor generation or increasing aqueous humor outflow working on the uveoscleral unconventional pathway to reduce IOP [5] but not the trabecular meshwork (TM) [6]. In fact, the TM and the adjacent Schlemm's canal endothelial lining is the main resistance site for aqueous humor outflow [7,8].
Recently discovered as novel IOP reduction drug classes, nitric oxide agonists and Rho kinase inhibitors work directly on the TM to increase aqueous humor outflow [9 –11]. It has been discovered that TM cellularity is reduced in aging and glaucomatous patients, which is associated with IOP increase [12 –14]. Our group and others have isolated and characterized multipotent stem cells from the TM [15 –20]. These TM stem cells (TMSCs), like other adult stem cells, have their own niche under Schwalbe's line [21 –24]. After transplantation into normal mouse anterior chamber, these human TMSCs were able to home to the TM region without immunorejection and inflammatory response [25]. After transplantation into the mouse anterior chamber with partial TM damaged by laser photocoagulation, these TMSCs were able to home to laser-damaged TM region and repair the tissue [26].
Other stem cell types, such as bone marrow-derived mesenchymal stem cells [27 –29] or induced pluripotent stem cells [30 –32], have also achieved success in restoration of abnormal TM. All these studies indicate that amelioration of diseased TM by introducing stem cells is a novel approach with great potential to reduce IOP and treat glaucoma [33 –35].
In our in vivo studies, we observed that intracamerally injected TMSCs preferentially distribute to murine TM in both wild-type mice and mice with laser-induced damage to TM. In contrast, corneal fibroblasts injected the same way with the same cell number disseminate in the iris, corneal endothelium, lens epithelium, and only part of the fibroblasts can reach murine TM [25,26]. The mechanism underlying the distribution difference between TMSCs and corneal fibroblasts remains elusive. Passive migration after the outflow of aqueous humor is inadequate to completely explain the distribution difference between TMSCs and fibroblasts.
Recently, we discovered that CXCR4/SDF1 chemokine axis guides homing of TMSCs to the laser-damaged TM region [26]. Nevertheless, we speculate that, resembling other types of stem cells [36], the homing process of TMSCs to the TM is precisely regulated by multiple factors. It has been confirmed that integrins had a versatile function in homing and maintaining stem cells to their niches [37]. For example, integrin-mediated adhesion has been demonstrated to participate in the recruitment of hematopoietic stem cells to bone marrow [37]. Moreover, the role of integrins in regulating stem cell proliferation and self-renewal has also been elucidated in different organisms [38]. However, the expression pattern of integrins in TMSCs and their role in the process of migration, location, and attachment of TMSCs to the TM still remains elusive.
In this study, we investigated that TMSCs express higher level of α5β1 integrin compared with corneal fibroblasts. Moreover, we discovered that fibronectin, a corresponding ligand of α5β1 integrin [39], mediates the binding of TMSCs to cultured TM cells and murine TM tissue. Functional blocking of α5β1 integrin in TMSCs attenuated attachment of TMSCs to the TM cells and fibronectin in vitro and shortened the retention time of TMSCs in murine TM tissue in vivo. Therefore, the highly expressed α5β1 integrin on TMSCs is correlated with TMSC integration in the TM but is not involved in the migration of TMSC cells to the TM.
Materials and Methods
Details of primary and secondary antibodies are listed in Supplementary Tables S1 and S2.
Primary cell cultures
Human corneas, including the limbus obtained from the Center for Organ Recovery and Education (Pittsburgh, PA), were used for isolation and cultivation of human TMSCs, TM cells, and corneal stromal fibroblasts. Cells were cultured and passaged as previously reported [15,20,40]. Human TMSCs were cultured in a medium containing Opti-MEM (Invitrogen, Carlsbad, CA) supplemented with 5% fetal bovine serum (FBS; ThermoFisher, Pittsburgh, PA), 10 ng/mL epidermal growth factor (EGF), 20 μg/mL ascorbic acid, 200 μg/mL calcium chloride, 0.08% chondroitin sulfate (Sigma-Aldrich, St. Louis, MO), 100 μg/mL bovine pituitary extract, 100 IU/mL penicillin, 100 μg/mL streptomycin, and 50 μg/mL gentamicin (ThermoFisher). Human TM cells and human corneal stromal fibroblasts were cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 with 10% FBS and the antibiotics mentioned previously. TMSCs and TM cells at passages 3–5 and fibroblasts at passages 5–7 were used for all the experiments in this study. All the experiments were repeated with cells from at least three different donors. TMSCs, TM cells, and fibroblasts were not necessarily derived from the same donors.
Cell affinity assay
The procedures we followed were published previously [26]. TMSCs stained with Vybrant DiI (ThermoFisher) were seeded onto three different conditions. Fibronectin (FN) coating group: 0.5 mL of fibronectin and collagen (FNC)-coating solution (Athena ES, Baltimore, MD) containing 10 μg/mL FN was coated at room temperature for 3 h on 8-chamber slides. FN+TM group: TM cells diluted at 1 × 106 cells/mL were labeled with Vybrant DiO (ThermoFisher) at 50 μg/mL for 20 min at 37°C. Ten thousand DiO-labeled TM cells were seeded on the FN-coated slides and cultured for 24 h before DiI-labeled TMSCs seeding. None group: TMSCs were seeded directly to nontreated chamber slides.
α5β1 integrin blocking
TMSCs were incubated with the blocking antibody against α5β1 integrin at a concentration of 10 μg/mL (MABT820) or 10 μg/mL control immunoglobulin G (IgG) (553987) for 24 h. TMSCs or TMSCs treated with the antibody or IgG were stained with DiI and seeded into the chambers with FN coating, FN+TM, or None for 60 min. The slides were washed with PBS twice to remove any unattached cells and fixed in 4% paraformaldehyde (PFA) for 15 min. The cells were stained with anti-fibronectin antibody at 4°C overnight, followed by secondary antibody and 4′,6-diamidino-2-phenylindole (DAPI; ThermoFisher) staining for 2 h. Images were taken under a confocal laser scanning microscope (Olympus FluoView FV1000 confocal microscope; Olympus, Center Valley, PA). Cells were counted and compared. At least five fields per well were photographed and four replicate wells were used for each condition.
Flow cytometry
To assess integrin expression in the cells, TMSCs and corneal stromal fibroblasts were cultured in 75 cm2 flasks and collected after reaching 70%–80% confluence. Cells were incubated with antibodies anti-α5β1 integrin (MAB1999), anti-αvβ3 integrin (MAB1976), or anti-αvβ5 integrin (MAB1961) for 1 h on ice. After washing and blocking with 1% bovine serum albumin (BSA), cells were labeled with a donkey-anti-mouse-488 secondary antibody for 30 min on ice. No staining and secondary antibody only were used as controls. Cell samples were run on a flow cytometer (BD Biosciences, San Jose, CA). The analysis was performed using FlowJo_V10 software (FlowJo, Ashland, OR).
For antibody endocytosis analysis, suspended or attached TMSCs were incubated with anti-α5β1 integrin antibody (MABT820) at 10 μg/mL. The incubation time for suspended TMSCs was 30 min on ice, whereas attached TMSCs were incubated for 30 min or 24 h. The cells were collected and washed with PBS and blocked with 1% BSA, incubated with donkey-anti-rat-488 for 30 min before running in a flow cytometer. This experiment was repeated three times for each group.
Animals
All experimental procedures were reviewed and approved by the University of Pittsburgh Institutional Animal Care and Use Committee and handled according to guidelines provided by the Association for Research in Vision and Ophthalmology Resolution on the Use of Animals in Ophthalmic and Vision Research. Passage-3 TMSCs were preincubated with the antibody against α5β1 integrin (10 μg/mL) or IgG (10 μg/mL) for 24 h. TMSCs were labeled with DiO and were resuspended in DMEM/F12 at a concentration of 1.67 × 107/mL for injection; 5 × 104 cells in 3 μL medium were injected into the anterior chamber of 9-week-old (male and female) C57BL/6 mice (Jackson Laboratory, Bar Harbor, ME) following procedures described previously [25,26].
The experimental groups were TMSCs (n = 12), TMSCs-α5β1 integrin (n = 12) or TMSCs-IgG (n = 12). DMEM/F12 medium (3 μL) was injected as sham controls (n = 12). IOP was measured using a rebound tonometer for rodents (TonoLab; Colonial Medical Supply, Franconia, NH) before and after cell injection at 1 month. All IOP measurements were performed around 1 pm on mice that had been anesthetized by intraperitoneal injection of ketamine (50 mg/kg) and xylazine (0.5 mg/kg) (IVX Animal Health, St. Joseph, MO), as described previously [25,26,41]. Mice were killed at 3 days or 1 month after transplantation. Mouse eyes were enucleated for wholemount and cryosectioning for histological analysis. Nine-week-old C57BL/6 mice (n = 6) received laser photocoagulation to damage the TM. The parameter for 532 nm diode laser composed of 80-mW power, 150-ms duration, and 50-μm spot size. Age-matched mice (n = 6) without laser treatment were used as normal controls.
Wholemount analysis
Enucleated mouse eyes (n = 3 for each group) were fixed with 1% PFA overnight at 4°C. The posterior part of eyes was cut off 1 mm of the limbus. The corneas including limbus were cut into quarters after removing the lens, iris, and ciliary body. The tissues were incubated with DAPI for 30 min, washed five times, and mounted to capture images to detect the DiO-labeled TMSC distribution on a confocal microscope.
Histological evaluation of TMSCs distribution
After fixation in 1% PFA overnight, the murine eyes (n = 3 for each group) were frozen at −20°C in optimal cutting temperature embedding compound (Tissue-Tek OCT; Electron Microscopy Sciences, Hatfield, PA) and cut into 10-μm thick cryosections on a cryostat (Leica, Buffalo Grove, IL). After dehydration in PBS, sections were incubated with DAPI to stain nuclei and photographed to detect the injected green cells using the confocal microscope.
Immunofluorescent staining
Cells were cultured on glass coverslips and fixed with 4% PFA. Fixed cells or mouse tissue cryosections were incubated with 0.1%Triton X-100 for permeabilization and 1% BSA to block any nonspecific binding. Samples were stained with appropriate primary antibodies overnight at 4°C followed by fluorescent secondary antibodies and DAPI staining. Pictures were taken with the same settings under the confocal microscope. Fluorescent intensity of images was analyzed using ImageJ software (National Institute of Health, Bethesda, MD). Mean fluorescent intensity of each group was obtained from at least three images.
For detection of antibody endocytosis, cultured TMSCs were incubated with 10 μg/mL anti-α5β1 integrin (MABT820) for 30 min or 24 h. The cells were fixed with 4% PFA and permeabilized with 0.1% Triton X-100. After blocking with 1% BSA, Donkey-anti-rat 488 was used to identify α5β1 integrin. All the experiments were repeated three times.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
TMSCs were seeded in 96-well plates with 1 × 104 cells per well. After incubation with 10 μg/mL anti-α5β1 integrin (MABT820) or 10 μg/mL IgG for 24 h, MTT solution [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] (5 mg/mL; Millipore) was added to the wells for 4 h at 37°C. Subsequently, 100 μL of pure dimethyl sulfoxide was added to each well after removing the culture medium to lyse the cells. The absorbance of each well was measured at 570 nm taking 600 nm as reference wavelength with the multimode Microplate Reader Infinite M Nano+ (Tecan, Switzerland). TMSCs without treatment were used as a control. The equation (OD sample − OD medium)/(OD control − OD medium) × 100%, was used to calculate the percentage of cell viability.
Quantitative real-time PCR
Total RNAs from cultured cells were isolated using an RNA purification kit (RNeasy Mini Kit; Qiagen, Valencia, CA). cDNAs were transcribed using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Quantitative real-time PCR (qPCR) was performed by direct dye binding (SYBR Green; Applied Biosystems). Primers were designed using Primer3 and blasted on the NIH website to confirm the specificity. The sequences of the primers are given in Supplementary Table S3. Relative mRNA level was calculated as the amplification Ct of a specific cDNA minus the average Ct of 18S as a power of 2 (2−ΔΔCt). Three repeats were averaged as mean ± standard deviation (SD).
Western blotting
Cultured cells were lysed with RIPA buffer containing protease inhibitors (Santa Cruz Biotechnology, Dallas, TX). Proteins were quantified using the BCA Protein Assay Kit (Pierce biotechnology, Waltham, MA). Total protein (30 μg per lane) was resolved on 4%–12% sodium dodecyl sulfate–polyacrylamide gel (ThermoFisher) and transferred to a PVDF membrane, blocked, and incubated with primary antibodies overnight. The membranes were incubated with secondary antibodies (IRDye 680LT, IRDye 800CW) and fluorescent signals were captured on an infrared imager (Odyssey Infrared Imager; LI-COR Biosciences, Lincoln, NE).
Statistical analysis
The results were expressed as mean ± standard error of the mean or mean ± SD for qPCR. The statistical differences were determined by Student's t-test or two-way analysis of variance followed by Bonferroni's multiple comparisons test. P < 0.05 was considered statistically significant.
Results
Cultured TMSCs express higher level of α5β1 integrin than fibroblasts
Previously, we demonstrated that the distribution patterns were different between TMSCs and human corneal stromal fibroblasts after anterior chamber injection [25,26]. Immunofluorescent staining (Fig. 1A, B) and western blotting (Fig. 1C, D) of primary human TMSCs and fibroblasts from three different donors showed that Nestin and OCT3/4 were highly expressed in TMSCs, whereas these stem cell markers could not be detected in fibroblasts. Alpha smooth muscle actin (α-SMA) could be examined in both TMSCs and fibroblasts with much lower intensity in TMSCs than in fibroblasts (Fig. 1A). As given in Fig. 1B, the expression levels of Nestin, OCT3/4, and α-SMA between TMSCs and fibroblasts were statistically significant. The quantification of mean fluorescent intensity was averaged from at least five fields of the staining. Figure 1D shows that expression levels of Nestin and OCT3/4 were statistically different, whereas the expression of α-SMA was not significantly different by western blotting. Quantification of the western blot data was averaged from three different strains of each cell type.

Different marker expression profile in TMSCs and corneal fibroblasts.
To determine if integrins played a role in the homing of stem cells to the TM, we first examined if TMSCs and fibroblasts have different integrin expression profiles. The expression of α5β1, αvβ3, and αvβ5 integrins, which play an important role in stem cells homing [37], was analyzed. As given in Fig. 2A, the mRNA levels of α5 and β3 integrins were significantly higher in TMSCs than in fibroblasts, whereas αv and β1 integrins were not significantly different between TMSCs and fibroblasts by qPCR. FACS analysis demonstrated that TMSCs expressed higher levels of α5β1 integrin than fibroblasts (P < 0.001), whereas both TMSCs and fibroblasts expressed similar levels of αvβ3 and αvβ5 integrins (P > 0.999) (Fig. 2B, C). Figure 2B shows representative figures of isotype controls and antibody staining for the aforementioned integrins. Figure 2C shows statistical analysis with three different cell strains of both TMSCs and fibroblasts.

Expression of integrins in TMSCs and corneal fibroblasts.
Western blotting further confirmed that the protein levels of α5 integrin (Fig. 3A, B) were significantly higher in TMSCs than in fibroblasts. Western blotting results on β1 integrin (Fig. 3C, D) also indicated that expression of β1 integrin was similar in both cell types. Immunofluorescent staining (Fig. 3E, F) of TMSCs further confirmed that expression of α5 integrin was higher on TMSCs than fibroblasts. Figure 3F shows the mean fluorescent intensity of the immunofluorescent staining.
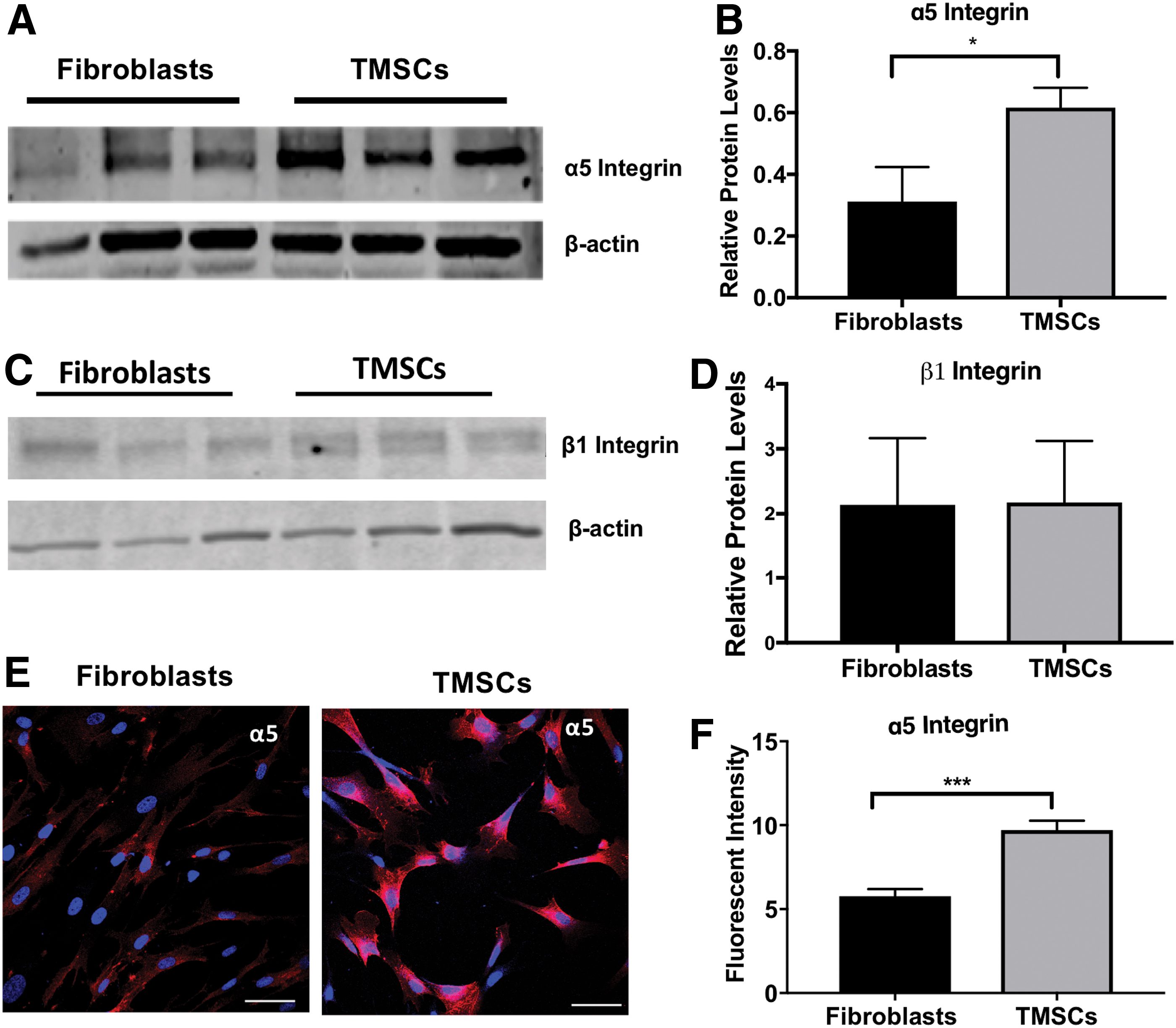
Expression of α5β1 integrin in TMSCs and corneal fibroblasts. Western blotting bands showing expression of α5 integrin
α5β1 integrin is involved in attachment of TMSCs to TM cells in vitro
Next, we evaluated the function of α5β1 integrin on TMSCs in cultures of normal TM cells using a cell affinity assay. Normal TM cell cultures were validated by analyzing their response to dexamethasone (Dex) [42]. As given in Supplementary Fig. S1A–D, TM cells responded to Dex with increased expression of myocilin (Myoc). The cultured Dex-treated TM cells also formed cross-linked actin networks (CLANs; Supplementary Fig. S1E) as previously reported [43 –45] and showed deposition of fibronectin after 5- and 14-day culture with and without Dex treatment (Supplementary Fig. S1F). The fibronectin deposition was dramatically increased after a 14-day Dex treatment (Supplementary Fig. S1G).
Fibronectin (FN), an α5β1 integrin ligand, or DiO-labeled TM cells and fibronectin (TM+FN) were seeded into 8-chamber culture slides to mimic the cellular and extracellular surroundings in the TM tissue. Subsequently, human TMSCs, TMSCs treated with α5β1 integrin blocking antibody (TMSC-α5β1-blocking), or TMSCs treated with IgG (TMSC+IgG) were seeded onto FN, TM+FN, or uncoated (None) chambers. One hour later, only a few TMSCs attached to the plates without any FN coating (Fig. 4A, None, 5.00 ± 1.00 cells/field). The number of TMSCs adhered to the fibronectin-coated plates (Fig. 4A; FN) increased to 28.66 ± 1.76 cells/field, and the cells adhered to plates with both TM cells and fibronectin (Fig. 4A; TM+FN) increased to 45.00 ± 5.50 cells/field. The attached TMSC numbers significantly decreased to 13.00 ± 2.52 cells/field on FN and 24.00 ± 5.29 cells/field on TM+FN after anti-α5β1 integrin antibody blocking (Fig. 4A, B). There was no difference between TMSCs and TMSC+IgG in comparison with the attached cell numbers with the same conditions. The attached TMSCs to FN and to TM+FN were similar with no statistical significance (Fig. 4B).
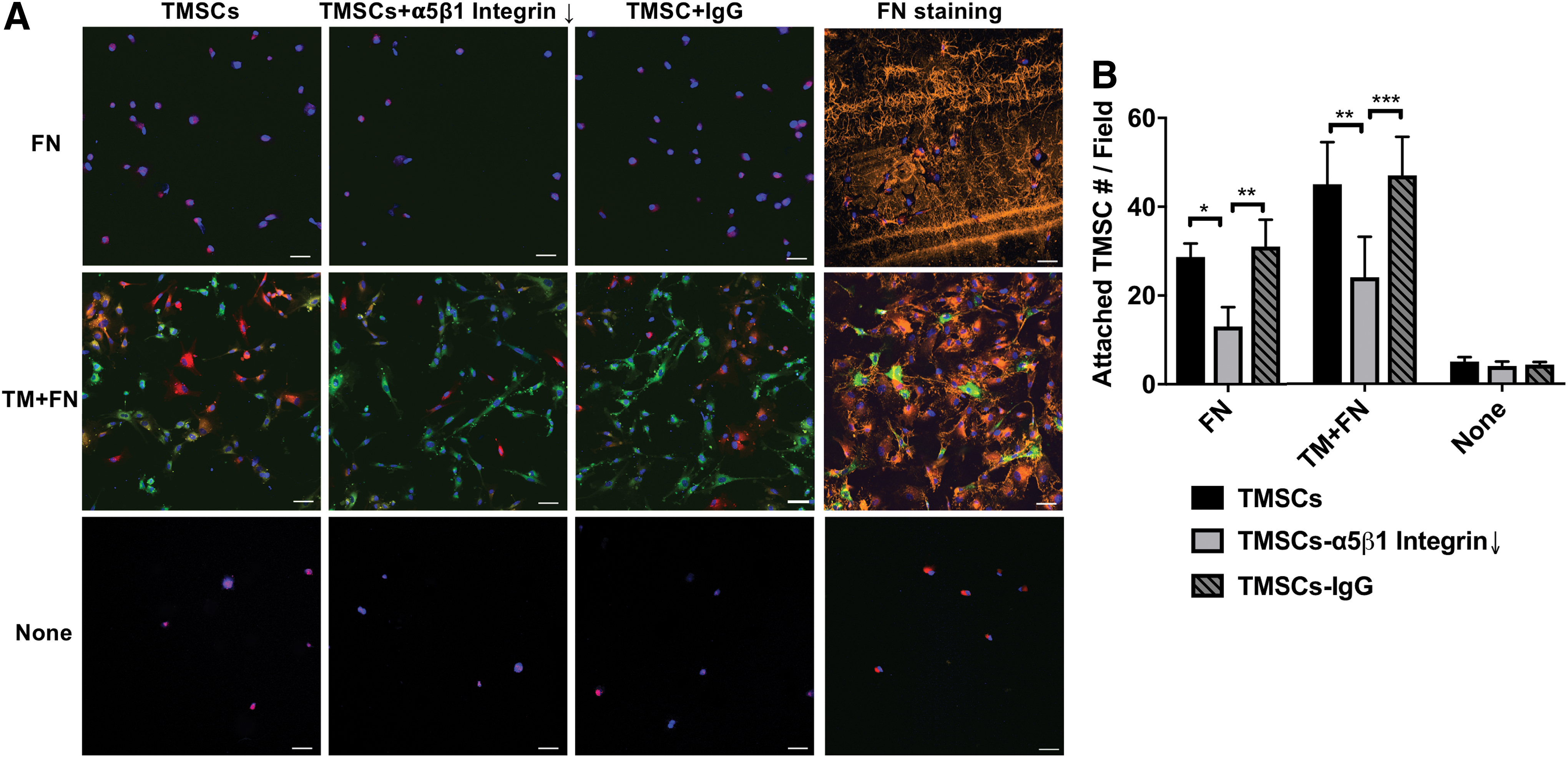
Cell affinity between TMSCs-TM cells and between TMSCs-matrix.
To determine whether the α5β1 integrin blocking antibody had been endocytosed during the incubation period for cell affinity assay, we did flow cytometry and immunofluorescent staining. These studies verified that the antibody was still on the cell surface and had not been endocytosed after the 24-h incubation (Supplementary Fig. S2).
Fibronectin is expressed at the site of TMSCs homing
Because we previously reported that TMSCs preferred to home to the laser-damaged TM region [26], we then examined if TM cells in vivo expressed fibronectin, which is the only ECM ligand that would bind the α5β1 integrin [46,47] on the TMSCs. As given in Fig. 5A, fibronectin was detectable at the TM in normal mice. The fibronectin staining in the laser damaged TM at 2 weeks was much stronger (Fig. 5B). Quantification of fluorescent intensity verified that positive signal of fibronectin in laser-injured TM area was greater than that in normal TM (P < 0.05, Fig. 5C). CD31 staining of Schlemm's canal endothelium and vessel endothelium confirmed that this region was the TM.
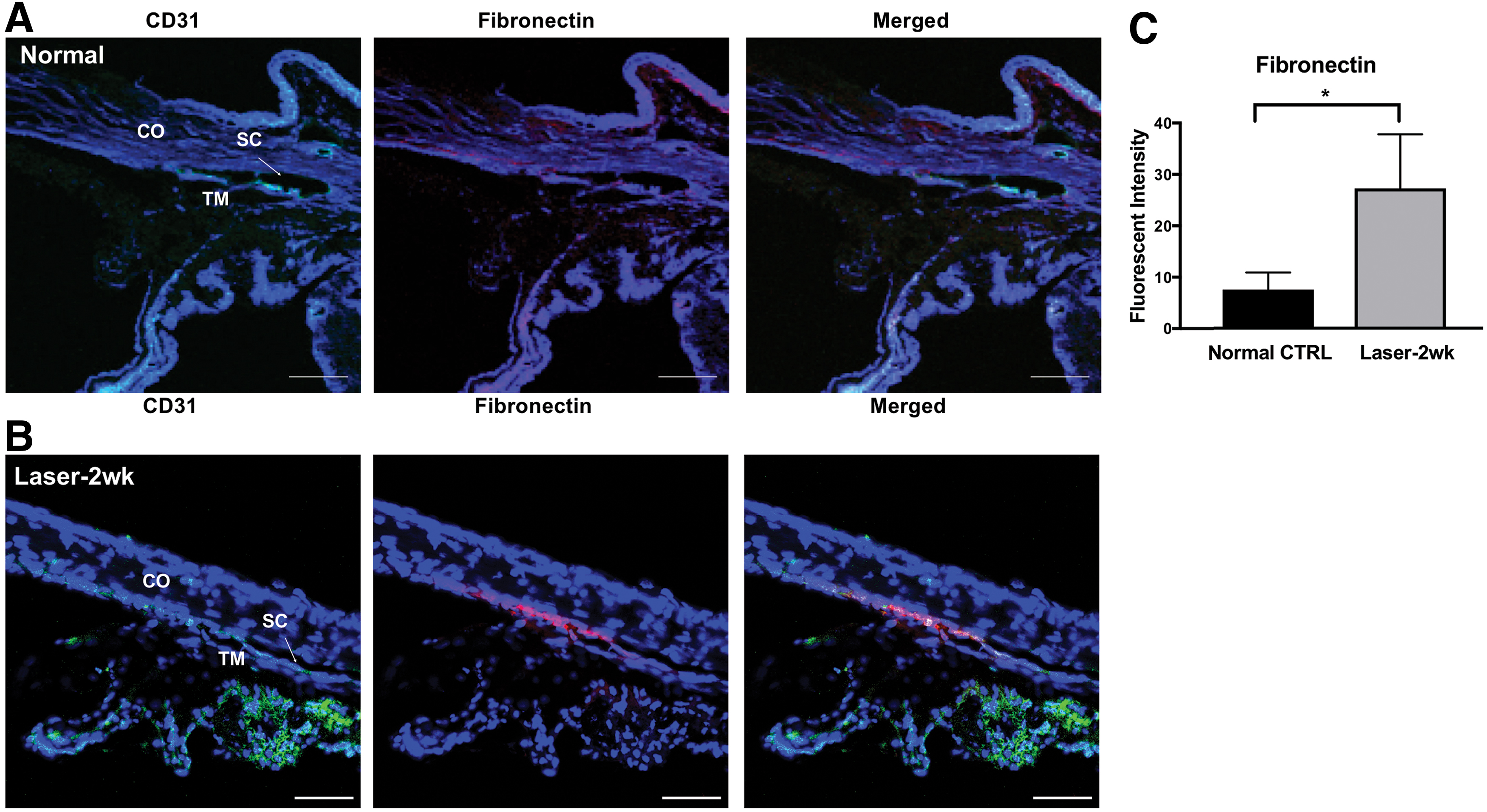
Expression of fibronectin in murine TM.
Blocking α5β1 integrin interrupts TMSCs retention in the TM tissue
To explore the function of α5β1 integrin in the process of TMSCs homing and anchoring to the TM tissue in vivo, we injected 5 × 104 DiO-labeled TMSCs into the anterior chamber of C57BL/6 mice. Three days after injection, the DiO fluorescent cells were mainly detected at the TM region and the injection site of the corneal endothelium, indicating TMSCs distributed mainly to the TM (Fig. 6A–F). No significant difference was observed among TMSCs (Fig. 6A, B), TMSCs with functional α5β1 integrin blocking (Fig. 6C, D), or TMSCs incubated with IgG (Fig. 6E, F). Intriguingly, 1 month after cell injection, the DiO-labeled TMSCs with α5β1 integrin blocking on the TM dramatically decreased (Fig. 6I, J) compared with TMSCs (Fig. 6G, H) and TMSC-IgG (Fig. 6J, K).
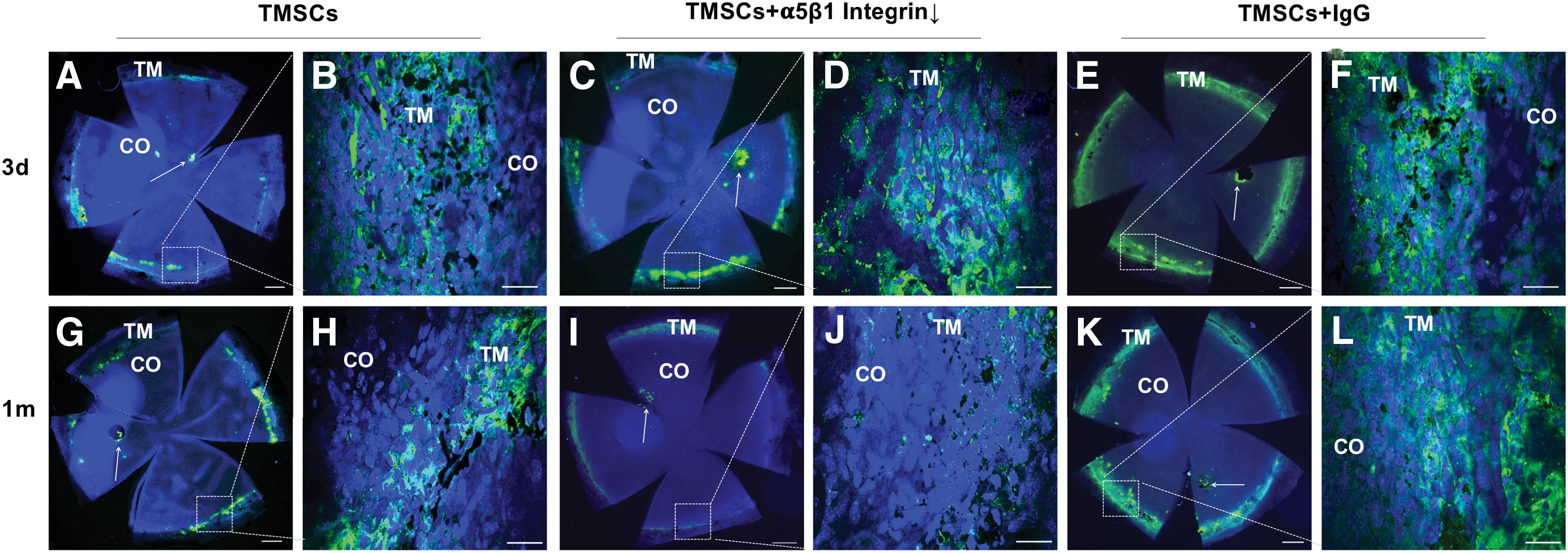
Wholemount analysis of TMSC homing and anchoring after intracameral injection. DiO-labeled green TMSCs, TMSCs treated with anti-α5β1 integrin antibody (TMSC+α5β1Integrin↓) or TMSCs treated with IgG (TMSC+IgG) were injected into the anterior chamber of wild-type C57BL/6 mice.
Histological analysis of cryosections from the mouse eyes (Fig. 7) showed similar results observed in the wholemounts as given in Fig. 6. Figure 7A shows that DiO-labeled TMSCs, TMSCs with the α5β1 integrin blocking, and TMSCs with a control IgG all homed to the TM region day 3 after the injection. Figure 7B shows that DiO-labeled TMSCs and TMSCs+IgG were still anchored in the TM 1 month after injection, whereas most of the homed TMSCs with the α5β1 integrin blocking disappeared from the TM region. Using a TUNEL assay, we did not detect any obvious apoptosis in the remaining TMSCs at both 3 days and 1 month shown by TUNEL staining (Supplementary Fig. S3). IOP between preinjection and 1 month postinjection among all groups remained stable (P = 0.989 to 0.999; Fig. 7C).
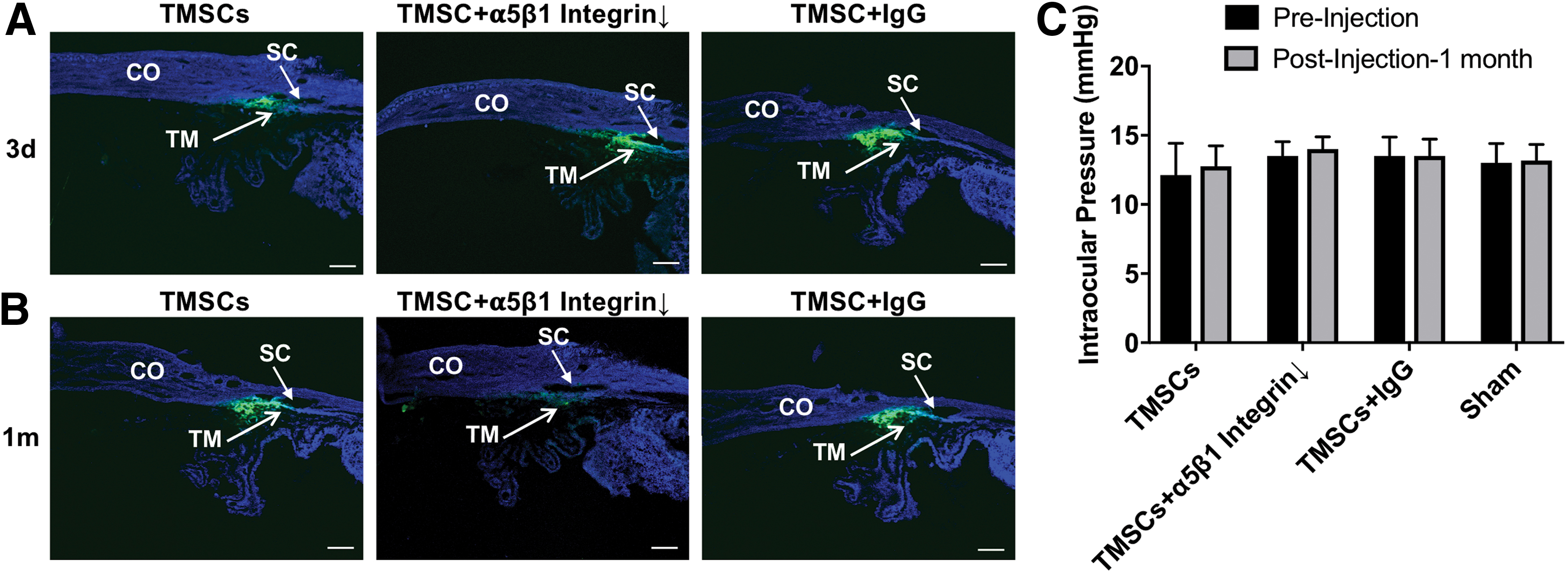
Cryosections show TMSC homing and integrating with cell injection have no effects on mouse IOP.
Discussion
In this study, we showed that although the α5β1 integrin was not needed for the migration of TMSCs to the TM and subsequent attachment to the TM, the α5β1 integrin was essential for homed TMSCs to integrate into the TM tissue. This suggests that the α5β1 integrin may play an important role in the regenerative function of TMSCs in situ. Whether different levels of α5β1 integrin expression could elucidate why stem cell remaining periods vary between studies remains to be determined. We have reported that human TMSCs could be detected in mouse TM tissue up to 4 months after injection [25]. Zhu et al. reported that iPSC-induced TM cells were detectable in mouse TM at 12 weeks after injection [31]. Another study reported that mesenchymal stem cells stayed in rat TM region for at least 48 h but could not be detected at 96 h after injection [27]. Clearly there is/are special mechanism(s) for anchoring and integrating of injected stem cells to the TM in vivo.
The expression and function of integrins vary significantly among cells. Some integrins are critical to the homing process for one type of stem cells to its niche, although they have no effects on other types of stem cells [46]. It has been reported that excessive expression and activation of α5β1 and αvβ3 integrins were correlated to mesenchymal stem cell homing and engraftment to the site of tissue injury [48]. Eleven integrins have been reported to be expressed in the TM tissue [39], but there is no report yet about integrin expression in TMSCs.
In this study, we report that TMSCs express at least three integrins. They were the α5β1, αvβ3, and αvβ5 integrins. Of those, the α5β1 integrin was highly expressed on the cell surface of TMSCs compared with corneal fibroblasts. We speculate that the higher level of expression of the α5β1 integrin in TMSCs compared with the corneal fibroblasts, despite the β1 integrin expression levels being similar in TMSCs and fibroblasts, was because of differences in expression of the another yet to be determined α-subunit. The β1 integrin subunit can form a heterodimer with α1–11 and αv integrin subunits [49].
This study showed that TMSCs can use α5β1 integrin to bind to fibronectin-expressing TM cells and fibronectin-coated substrates in vitro. Using a function blocking α5β1 integrin antibody dramatically reduced the numbers of TMSCs adhered to both TM cell cultures and fibronectin-coated substrates in vitro. This finding indicates that interactions between the α5β1 integrin and fibronectin could be involved in TMSCs binding to the TM tissue in vivo. The enhanced expression of fibronectin in the TM of mice with laser photocoagulation compared with that of normal mice, also suggests that an interaction between upregulated fibronectin in the murine TM and excessively expressed α5β1 integrin on TMSCs is likely to contribute to the integration of TMSCs in the laser damaged TM.
Of interest, blocking α5β1 integrin in TMSCs did not alter their distribution pattern after the intracameral injection. Three days after cell injection, TMSCs could still home to the TM region regardless of whether α5β1 integrin was inhibited or not. This suggests that other adhesion molecules, such as cadherins [50] and other integrins might be involved in directing the migration of TMSCs to the TM.
Despite not playing a role in migration, our study showed that the highly expressed α5β1 integrin can assist homed TMSCs to lodge and integrate into the TM. Functional blocking of α5β1 integrin dramatically reduced the number of TMSCs attached to TM cells in vitro and shortened the time of TMSCs staying in the murine TM, further emphasizing the importance of α5β1 integrin in this step. One month after injection, most of the TMSCs treated with α5β1 integrin antibody disappeared, whereas untreated TMSCs and TMSCs treated with IgG stayed in the TM region. It is still unknown why the TMSCs treated with α5β1 integrin antibody disappeared. Our study indicated that it was not because of the antibody or a disruption in α5β1 integrin–fibronectin binding inducing TMSC cell death, which may lead to reduced cell number staying at the TM tissue after intracameral injection.
This study confirms our previous study that TMSCs could remain in the TM for up to 4 months after the intracameral injection [25]. This result highlights the importance of α5β1 integrin–fibronectin interactions in the long-term maintenance and integration of TMSCs in the TM and suggests that the loss of α5β1 integrin-mediated adhesion can lead to failure of TMSC integration in the TM. This suggests that the mesenchymal stem cells were not detected at 96 h [27] might be at least partially related to the integrin activity.
Our previous study confirmed that TMSCs actively home to the TM mainly enforced by chemotactic factors CXCR4/SDF1 [26]. Meanwhile, passive drifting through the outflow of aqueous humor is speculated to play a minor role in this process, which drives a small portion of injected fibroblasts to the TM region [25,26]. After homing, the next step is anchoring and integrating of TMSCs to the TM. In this study, we demonstrated that highly expressed α5β1 integrin can assist homed TMSCs to lodge and integrate in the TM by binding to fibronectin.
In summary, our study indicates that α5β1 integrin expression on the surface of TMSCs interacts with fibronectin in the TM cells and TM matrix to promote the anchoring and integration of TMSCs for regeneration. A better understanding of molecular mechanisms underlying TMSCs homing and anchoring to the TM will support the development of novel strategies for improving the efficiency of applying TMSCs for glaucoma treatment in the future.
Footnotes
Acknowledgments
The authors thank Nancy Zurowski (Department of Ophthalmology) for assisting with Flow Cytometry and Kira Lathrop for helping with confocal microscopy.
Author Disclosure Statement
University of Pittsburgh owns a patent “trabecular meshwork stem cells” with Y.D. as one of the inventors. All the other authors declare that they have no conflicts of interest.
Funding Information
The study was supported by NIH grants EY025643 (Y.D.), P30-EY008098, EY017006 (D.M.P.), EY026009 (D.M.P.), Research to Prevent Blindness; and Eye and Ear Foundation (Pittsburgh, PA).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.