
Abstract
Inflammatory response regulation is a mechanism through which human umbilical cord mesenchymal stem cells (HUCMSCs) improve myocardial ischemia reperfusion injury (IRI); however, the timing of HUCMSC delivery to achieve maximum effectiveness is controversial. To investigate the effects of HUCMSC delivery on the acute inflammatory stage of IRI, we transplanted HUCMSCs or HUCMSCs with cyclosporin A (CsA) through the coronary artery simultaneously during ischemia reperfusion in pigs. Ferumoxytol-labeled HUCMSCs (HUCMSC), HUCMSCs with cyclosporin A (HUCMSC+CsA), and PBS (control) groups were investigated to evaluate the homing of transplanted cells and changes in infarct features, cardiac activity, and inflammatory response at three time points post-transplantation. Animals were sacrificed 2 weeks later for histological analysis of the hearts. We detected Prussian blue-dyed granules distributed around T lymphocyte clusters in the infarct area in the HUCMSC group. Infarct size and collagen deposition in the infarct area were lower in the HUCMSC group than in the control and HUCMSC+CsA groups. Cardiac function was mildly impaired in both the control and HUCMSC groups, whereas added CsA had a more severe impact. The levels of proinflammatory markers were lower in the HUCMSC group than in the control group at 24-h follow-up, and the difference was more significant after adding CsA. There were more CD3+ T lymphocytes and Foxp3+ Tregs in the HUCMSC group infarct area than in the other two groups. Proliferation rate of T lymphocytes was higher in the HUCMSC group than in the other two groups. Indirect co-culture experiments in vitro showed that MSCs promoted the generation of CD4+CD25+ Foxp3+Tregs through a paracrine mechanism. These results indicate that immediate intracoronary delivery of HUCMSCs after ischemia reperfusion can reduce acute myocardial IRI and promote myocardial repair, mainly through T lymphocyte interactions to regulate the intense inflammatory response during the acute inflammatory stage.
Introduction
Recently, morbidity of acute myocardial infarction (AMI) has been increasing worldwide. Although timely revascularization is the most effective approach for salvaging the myocardium from death, reperfusion of the ischemic myocardium can induce further and varying degrees of myocardial injury, known as ischemia reperfusion injury (IRI) [1].
Studies have shown that the inflammatory immune response plays a vital role in IRI and is divided into three stages: acute inflammatory injury stage, proliferative repair stage, and chronic inflammatory stage [2]. At the beginning of reperfusion, proinflammatory cells and cytokines are rapidly recruited to the myocardial infarction site to clear dead cells and extracellular matrix, providing space for subsequent proliferation and repair. However, excessive inflammatory responses tend to aggravate myocardial injury, inducing inadequate myocardial repair and myocardial remodeling [3]. Cyclosporin A (CsA) was used to treat myocardial IRI as an immunosuppressor [4]. Preclinical studies have shown that CsA can inhibit T lymphocyte activation by binding to cyclophilin to block the inflammatory response [5] and reduce cell apoptosis in IRI [6]. However, clinical trials showed disappointing results [7 –9], indicating that regulating rather than simply blocking inflammatory responses is more important for attenuating myocardial IRI.
Mesenchymal stem cells (MSCs) have been shown to have extraordinary immunomodulatory properties, in addition to cardiovascular regeneration abilities [9,10]. Among these cells, human umbilical cord MSCs (HUCMSCs) exhibit less immunogenicity and better immunomodulatory properties than other MSCs from different sites [10]. Correspondingly, MSCs were shown to reduce intense inflammatory responses to inflammatory stimuli, but enhance these reactions during the inflammation remission period [11,12]. In the acute phase of inflammation, MSCs strongly inhibit inflammation by suppressing the activation of effector T lymphocytes [13]. Hence, the early phase of myocardial IRI may be an ideal time to administer MSCs to relieve violent inflammatory reactions. Although previous studies suggested that 1 week after reperfusion is the ideal administration time, as it is more suitable for transplanted cells survival [14], the administration time remains controversial.
Therefore, a porcine model of acute myocardial ischemia and reperfusion was developed and HUCMSCs were transplanted by intracoronary infusion immediately at the time of reperfusion to investigate the feasibility of HUCMSC transplantation in the acute inflammatory injury stage. Furthermore, CsA was added before cell therapy to block T lymphocyte function, to evaluate the influence of the inflammatory environment on HUCMSC homing and explore the immune regulation effects of HUCMSCs.
Materials and Methods
Experimental overview
This study involved three steps. In the first step, a myocardial IRI model was established. After the distal side of the left anterior descending artery (LAD) of pigs was blocked with an over-the-wire balloon for 90 min, blood flow was restored. In the second step, the experimental pigs were divided into three groups: (1) HUCMSC group: immediately after the over-the-wire balloon was deflated, 5 × 107 HUCMSCs were injected through the LAD; (2) HUCMSC+CsA group: CsA (Sandimmune®; NOVARTIS, Basel, Switzerland) was infused intravenously at a dose of 5 mg/kg body weight 30 min before blood flow restoration and, immediately after reperfusion, 5 × 107 HUCMSCs were injected through the LAD; and (3) control group: PBS was injected through the LAD immediately after myocardial infarction and reperfusion. In the third step, cardiac magnetic resonance (CMR) imaging was performed on a 1.5-T scanner (Philips Achieva, Cleveland, OH) at 24 h and 1 and 2 weeks after the second step to evaluate changes in cardiac function, infarct size, and regional myocardial perfusion. Serum inflammatory markers, including interleukin (IL)-10, IL-6, and C-reactive protein (CRP), were analyzed at the three time points. Animals were sacrificed 2 weeks later and the infarct areas, infarct border areas, and normal areas were embedded and fixed to make paraffin sections. Three slices of 5–8 μm thickness were sectioned from each paraffin-embedded block using microtome for histopathology (Fig. 1).
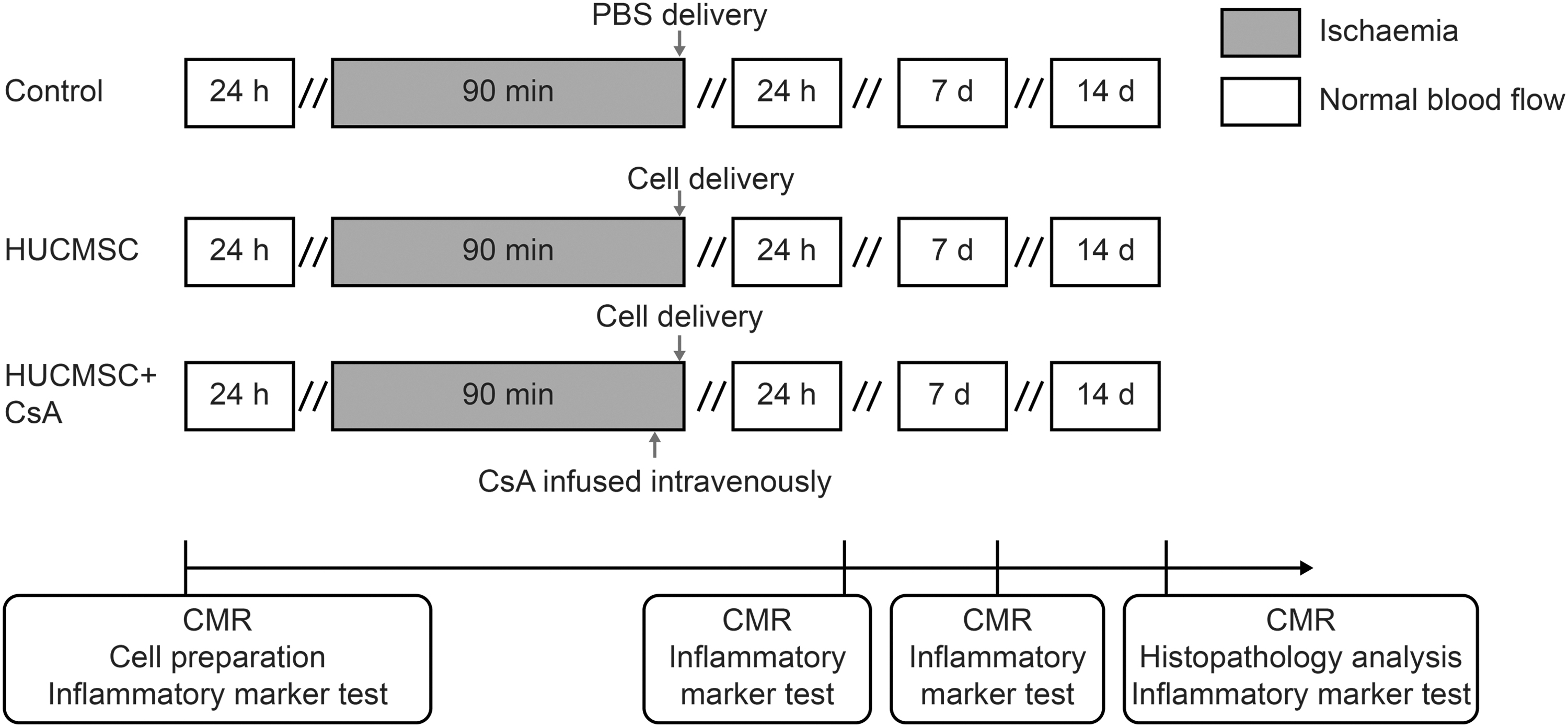
Experimental protocol.
Experimental animals
Animal studies were approved by the Animal Ethics Committee of Nanjing University and complied with the Chinese National Regulations on the Use of Experimental Animals. Nineteen female Chinese experimental minipigs (20 ± 5 kg) were obtained from Jiangsu Academy of Agricultural Sciences (Nanjing, China).
Cell culture and labeling
Passage 2 HUCMSCs were obtained from Nanjing Drum Tower Hospital Clinical Stem Cell Center (Nanjing, China). The cells were incubated in DMEM (low glucose; Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (Invitrogen) and 1% antibiotic-penicillin and streptomycin solution (Invitrogen). Approximately 5 days later, these cells had reached confluence, and thus subculturing was performed after detaching the cells with 0.25% trypsin solution containing 0.01% EDTA (Invitrogen). Passage 6 cells were used for transplantation, and cell characteristics, including CD44, CD73, CD90, CD105, CD19, CD11b, CD34, CD45, HLA-DR, and HLA-DQ, were tested with the corresponding anti-human antibodies (eBioscience, San Diego, CA) and flow cytometry. To trace cells in vivo and in vitro, we labeled MSCs with 50 μg/mL ferumoxytol, a new ultra-small superparamagnetic iron oxide, in the presence of poly-L-lysine for 24 h. This resulted in ∼100% labeling efficiency in previous studies [15,16]. The labeling efficiency was determined using a Prussian blue staining kit (Abcam, Cambridge, United Kingdom).
Myocardial ischemia reperfusion procedure
Animals were administered propofol for general anesthesia. A guiding catheter was advanced to the left coronary artery through a femoral artery puncture. After a coronary angiogram, an over-the-wire balloon catheter was placed in the distal LAD near the second diagonal branch through the guiding catheter and inflated for 90 min of blood flow occlusion, followed by balloon deflation. A myocardial ischemia reperfusion model was successfully established when dynamic evolution of ST-segment elevation and retreat was observed through electrocardiography monitoring (Philips, Amsterdam, Netherlands).
Cell and CsA delivery
A total of 5 × 107 HUCMSCs suspended in 10 mL of PBS, or 10 mL of PBS alone was infused through the over-the-wire balloon catheter immediately after balloon deflation. For the HUCMSC+CsA group, CsA was intravenously injected 30 min before cell infusion.
Cell tracing
CMR imaging of T2*-weighted gradient-echo scans was performed to trace iron-labeled MSCs in vivo as in our previous study [15]. After the animals were sacrificed, paraffin-embedded heart sections were stained with Prussian blue to detect the iron particles and calculated with Image-Pro Plus 6.0 software (Media Cybernetics, Rockville, MD).
Infarct size and collagen deposition analysis
Late gadolinium-enhanced (LGE) CMR imaging was performed to confirm the infarct size. The infarct size was calculated automatically as the ratio of the area of myocardium with delayed enhancement in each slice to the area of left ventricular (LV) myocardium from the short-axis delayed-enhancement images [17].
Local myocardial collagen deposition in sections was detected with a Masson's Trichrome Staining kit (Abcam), and the percentage of the positive staining area in a unit region was calculated and analyzed by Image-Pro Plus 6.0 software.
Regional myocardial perfusion analysis
Myocardial perfusion was evaluated with first-pass perfusion imaging of CMR [15]. Short-axis sections were positioned covering the infarct area as identified by gadolinium-enhanced imaging, as well as the basal and LV blood pool. Time-intensity curves were generated, and the area under the curves was computed to reflect blood supply in a certain region [18].
Microvascular obstruction (MVO) was detected by CMR as a persisting area of low signal, surrounded by enhanced myocardial tissue on LGE images [19].
Paraffin sections were stained with an anti-von Willebrand factor (vWF) antibody (Abcam) to measure microvessel density. The numbers of vWF-positive cells were determined by counting at least 10 random fields in each section in a blinded manner at 100 × magnification. Four sections of each heart were measured and averaged.
Cardiac function analysis
Cine MRI was performed using a balanced fast field echo sequence in the long- and short-axis orientations. Analyses of left ventricular ejection fraction (LVEF), left ventricular end-diastole volume (LVEDV), and wall motion were performed with the Cardiac Explorer software package [15].
Serum inflammatory marker test
Venous blood samples were collected and centrifuged. Serum IL-6, IL-10, and CRP levels were analyzed by porcine quantikine enzyme-linked immunosorbent assay (ELISA) (R&D Systems, Minneapolis, MN) following the manufacturer's protocols.
Analysis of T lymphocyte infiltration and proliferation
Paraffin sections from the infarct zone were immunostained with rabbit anti-CD3 antibody (Abcam) to identify T lymphocytes. The results were calculated and analyzed with Image-Pro Plus 6.0 software.
A double-labeling immunofluorescence staining protocol was used to detect cell proliferation in CD3 cells with anti-Ki67 antibody (Abcam) and anti-CD3 antibody (Abcam). The proportion of CD3+- and Ki67+- positive cells among CD3+ cells indicated the proliferation rate of T lymphocytes.
Paraffin sections of the infarct zone were immunostained with an anti-mouse/anti-rat Foxp3 antibody (eBioscience) to identify Foxp3+ cells. The results were calculated and analyzed with Image-Pro Plus 6.0 software.
Isolation of human peripheral blood mononuclear cells
Peripheral blood samples were obtained from three healthy male volunteers (donor age 25–33 years). All volunteers provided written informed consent, and the study was approved by the local ethics committee on human research (2020-190-05). Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation and cultured in RPMI-1640 medium (Gibco, Grand Island, NY) containing 10% fetal bovine serum (Gibco) and 1% penicillin/streptomycin (Gibco) in a 5% CO2 atmosphere at 37°C. The cell count and viability were determined using the trypan blue exclusion method.
Indirect co-culture experiments
To evaluate the paracrine mechanism of HUCMSCs on T lymphocytes, indirect co-culture experiments were performed. HUCMSCs were implanted in a 12-well plate (106 cells/well). After 12 h, wells were separated with semipermeable membrane (pore size 0.4 μm; Millicell, Millipore, Billerica, MA). PBMCs were stimulated with 10 mg/mL mitogen phytohemagglutinin (PHA) and cultured in the upper chambers, whereas HUCMSCs were cultured in the lower chambers (1:1) for 3 days. PBMCs and medium supernatants were collected on days 1 and 3 for flow cytometry analysis and ELISA analysis. Flow cytometry was performed using FACS Aria flow cytometer (BD Biosciences, Franklin Lakes, NJ), and data were analyzed with FlowJo software (TreeStar, Ashland, OR). PBMCs were incubated with antibodies for human CD3 APC, CD4 FITC, CD8 PE, CD25 PE, and FoxP3 Alexa Fluor (BD Biosciences) according to the manufacturer's instructions. The protein levels of IL10 and TGF-β1 were detected using the corresponding ELISA kit according to the manufacturer's instructions (Biolegend, San Diego, CA).
Statistics analysis
Continuous variables are presented as the mean ± SD as indicated. Differences between two groups were tested using the independent samples t-test, whereas differences between three groups were compared by one-way analysis of variance followed by the least significant difference test. Changes from baseline within groups were compared by paired Student's t-test. The chi-square test was used to compare the rates between groups. All hypothesis tests were 2 sided, and a P value <0.05 was considered statistically significant. All statistical analyses were performed with SPSS 17.0 software (SPSS, Inc., Chicago, IL).
Results
HUCMSC preparation and labeling with ferumoxytol
HUCMSCs were expanded to passage 6 before transplantation, and cell characteristics were tested by flow cytometry. Flow cytometry confirmed that passage 6 HUCMSCs had characteristics consistent with those of MSCs, with ≥95% of cells expressing CD44, CD73, CD90, and CD105, whereas the expression of CD19, CD11b, CD34, CD45, HLA-DR, and HLA-DQ was 1% or less (Fig. 2A, B). Prussian blue staining showed that almost all ferumoxytol-labeled cells were stained (displaying blue-stained particles) (Fig. 2C), whereas unlabeled cells were not stained (Fig. 2D).

HUCMSC preparation. (
AMI reperfusion model establishment
A total of 19 pigs were used to prepare the myocardial IRI model by 90 min of balloon inflation of LAD followed by balloon deflation (Fig. 3A). Successful acute myocardial ischemia was evidenced by substantial ST-segment elevation in electrocardiography monitoring (Fig. 3B), LGE MRI scanning (Fig. 3C), and anatomical images (Fig. 3D). Ventricular fibrillation occurred during blood flow occlusion in seven pigs, six pigs were rescued successfully and one pig died. The remaining 18 pigs were randomly assigned to three groups of six pigs each: the control group, HUCMSC group, and HUCMSC+CsA group. Two pigs in the control group and one pig in the HUCMSC+CsA group were excluded because they died of unknown causes within 1 week after model establishment.
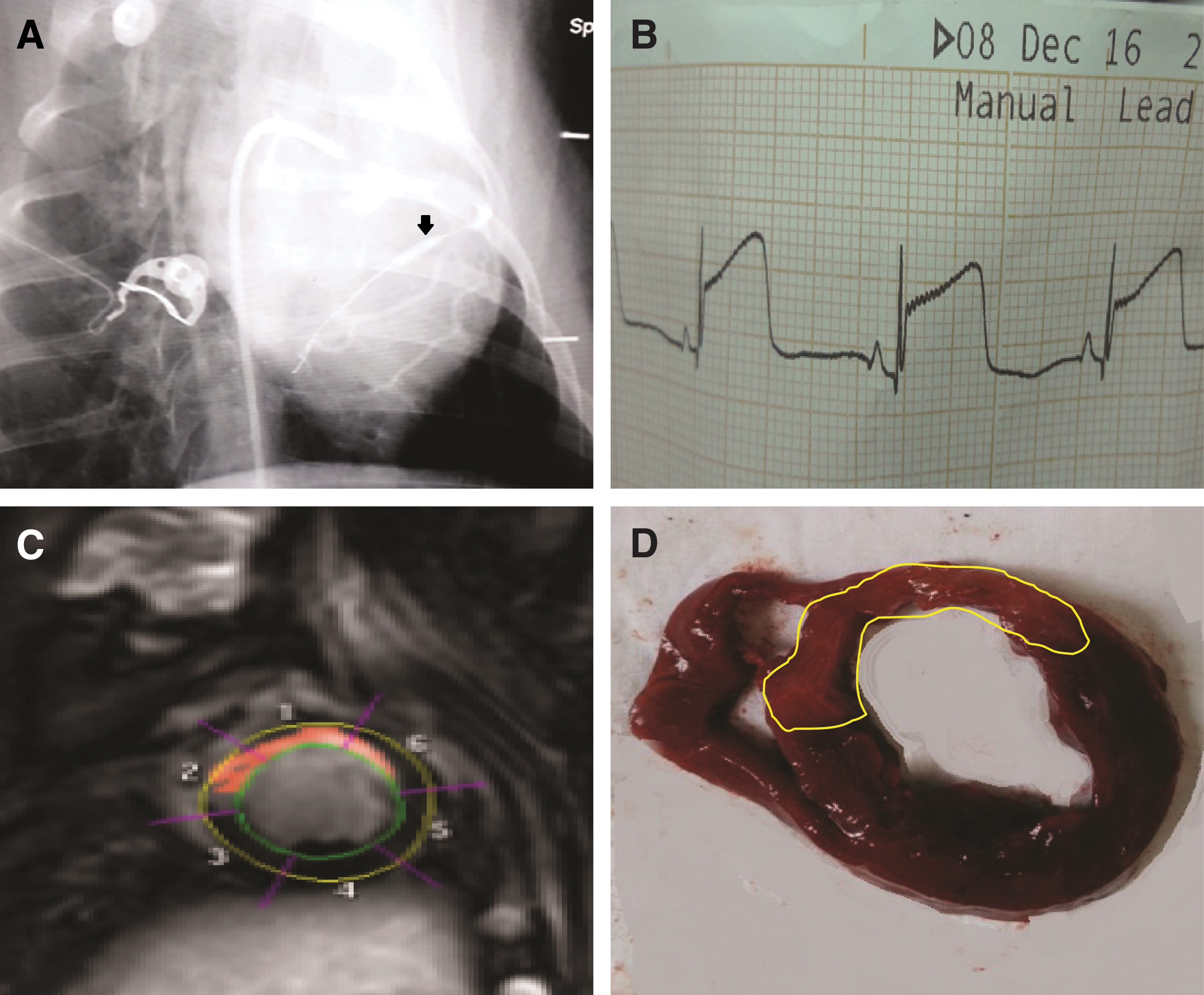
AMI reperfusion procedure.
Cell tracing
T2*-weighed images showed no hypointense signal in any of the three groups at any time point (Fig. 4A). Prussian blue staining revealed that the number of blue-dyed granules in the infarct area was significantly higher in the HUCMSC group than in the HUCMSC +CsA group (71.3 ± 6.9 vs. 20.6 ± 3.7, P < 0.01), whereas no blue-stained particles were found in the control group (Fig. 4B, C). Particularly, blue-stained particles in the HUCMSC group were distributed around some cell clusters (Fig. 4B).
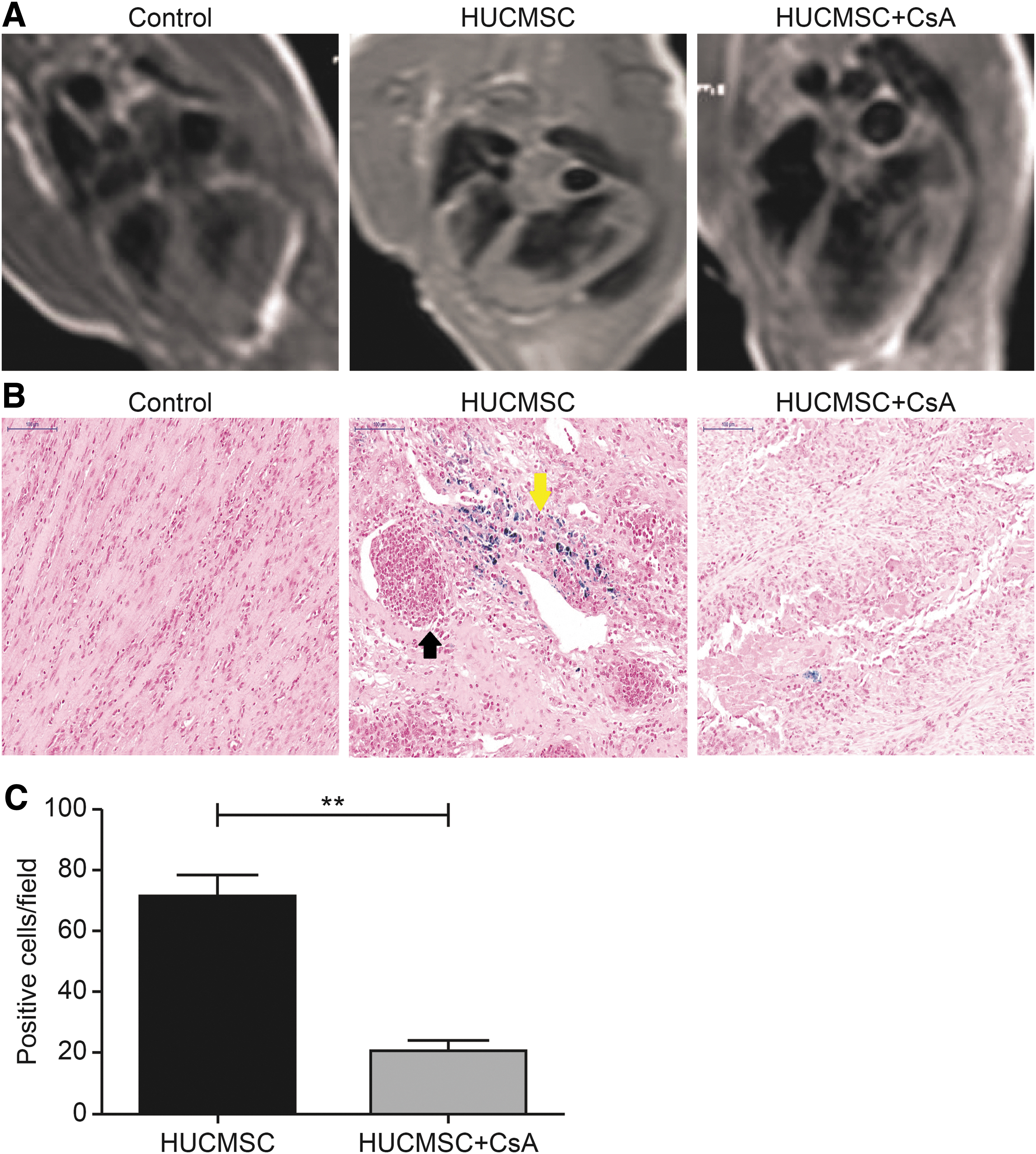
Cell tracing.
Infarct size and collagen deposition
The percentage of infarcted LV in the control group was 22.2% ± 1.5% and improved to 12.1% ± 4.4% in the HUCMSC group as observed by gadolinium-enhanced CMR scans at 24 h after transplantation (P < 0.05), whereas CsA addition before HUCMSC perfusion increased the infarct size to 23.8% ± 3.1% (P < 0.01), which was similar to that in the control group. At two follow-up time points, the infarct area of each group changed moderately, and the HUCMSC group was consistently lower than the other two groups (Fig. 5A–C). Two cases of serious MVO in the HUCMSC+CsA group were detected (Fig. 5A). Collagen deposition in the infarct area of the HUCMSC group was significantly lower than that in the control group (72.0% ± 5.3% vs. 87.7 ± 2.5, P < 0.001) and the HUCMSC+CsA group (72.0% ± 5.3% vs. 86.6 ± 3.5, P < 0.001). There was no significant difference in the infarct border zone between the three groups (Fig. 5D, E).
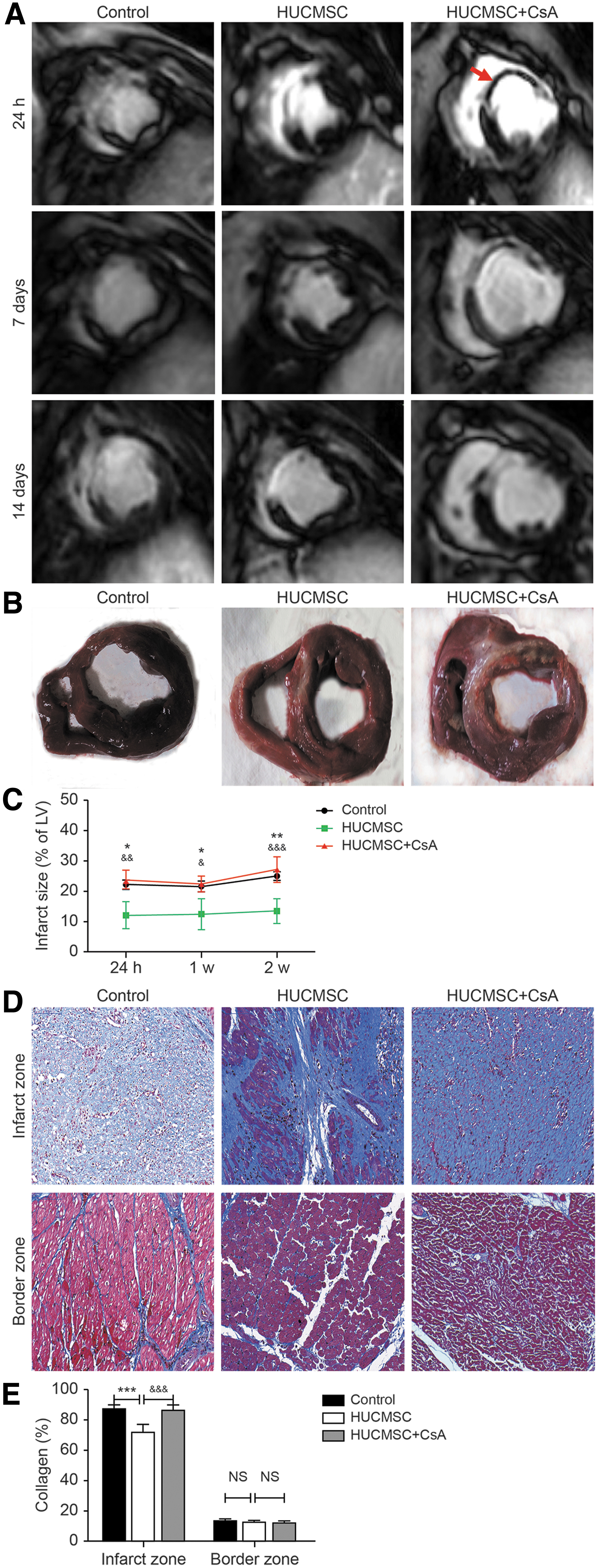
Infarct size and collagen deposition.
Regional myocardial perfusion change
Time-intensity curves of the infarct area and remote myocardial and LV blood pool curves were generated, and the area under the curves was computed as a parameter reflecting blood supply in certain regions (Fig. 6A–C). The ratio of the area under the curve of the infarct zone to that of the remote zone was calculated to represent the reduced perfusion efficiency in myocardial infarction. The results showed that perfusion of the infarct zone was impaired at 24-h follow-up and gradually recovered by later follow-up time points. The perfusion efficiency of the HUCMSC group changed from 54.5% ± 5.2% at 24-h follow-up to 83.5% ± 8.3% at 2-week follow-up, which was similar to the corresponding change in the control group (56.3% ± 8.2% at 24-h follow-up and 75.2% ± 12.3% at 2-week follow-up), whereas CsA treatment before HUCMSC perfusion led to a more severe decrease in the perfusion rate (35.3% ± 7.3% at 24-h follow-up and 46.2% ± 13.6% at 2-week follow-up, P < 0.001 and P < 0.01 vs. the HUCMSC group, respectively) (Fig. 6D). The number of new blood vessels in both the infarct and border areas was higher in the HUCMSC group than in the other two groups (Fig. 6E, F).

Change in regional myocardial perfusion.
Global and regional LV systolic function
There was no difference in LVEDV, LVEF, or systolic wall thickening between the groups at baseline. LVEDV expanded mildly in both the control and HUCMSC groups, whereas addition of CsA had a more severe impact, particularly at 2-week follow-up, compared with that in the HUCMSC group (47.6 ± 9.1 cm3 vs. 25.3 ± 5.4 cm3, P < 0.001) (Fig. 7A). After AMI reperfusion, LVEF decreased in all groups. In the HUCMSC group, LVEF mildly decreased from 64% ± 1.8% before AMI reperfusion to 53.1% ± 4.6% at 1-week follow-up and recovered to 58.6% ± 9.0% at 2-week follow-up, which was similar to the corresponding changes in the control group. In contrast, LVEF decreased sharply from 60.1% ± 1.2% before AMI reperfusion to 29.7% ± 2.5% in the HUCMSC+CsA group at 24-h follow-up and recovered slowly to 38.7% ± 3.3% at 2-week follow-up, which was significantly lower than that in the HUCMSC group at three time points (P < 0.001) (Fig. 7B). Systolic wall thickening of the infarct area was impaired after AMI reperfusion in each group. The reduction was most pronounced in the HUCMSC+CsA group, namely, from 60.3% ± 2.9% before AMI reperfusion to 33.6% ± 4.3% at 2-week follow-up (Fig. 7C, D).
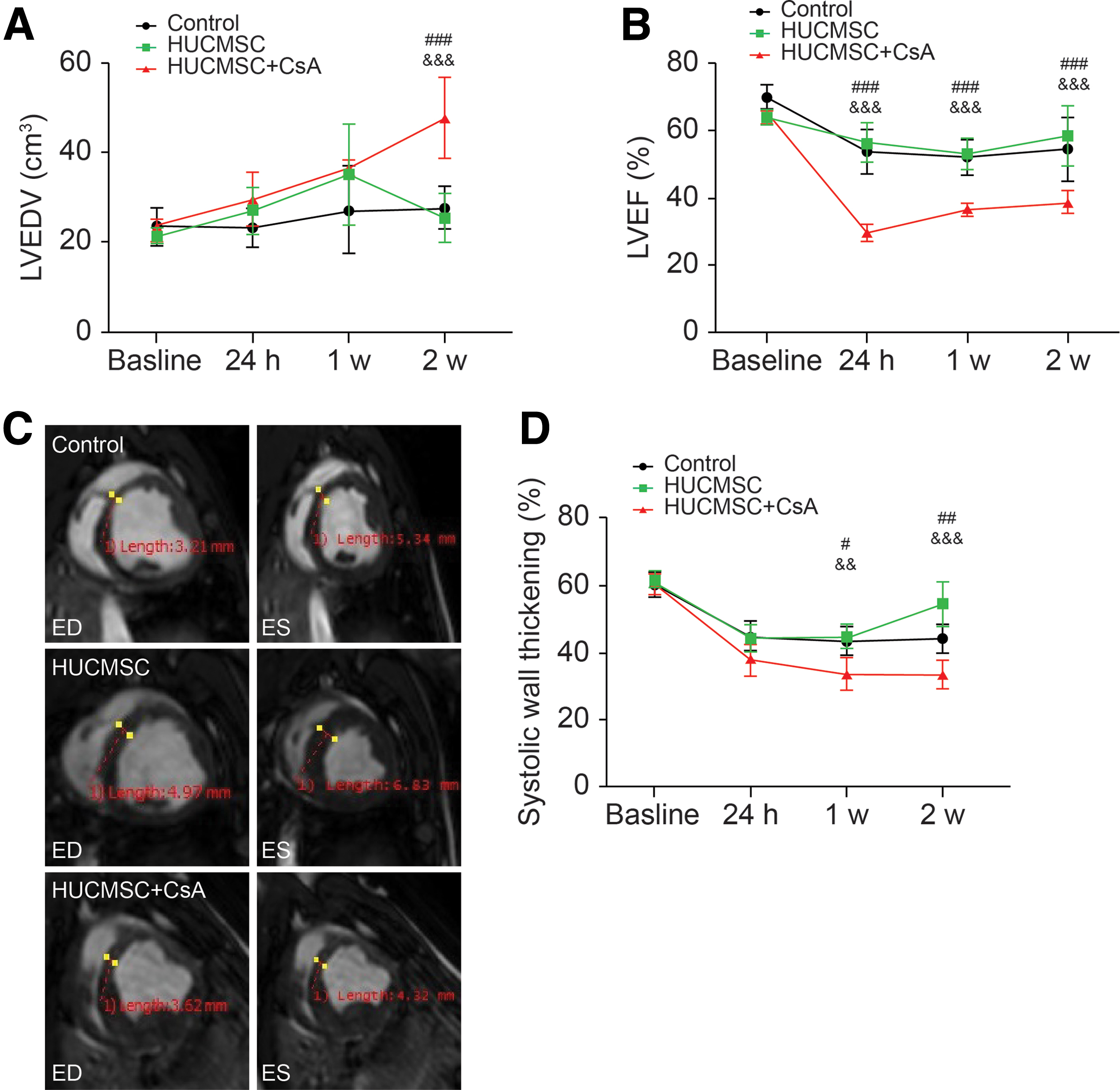
Global and regional LV systolic function.
Changes in serum inflammatory marker levels
There was no difference in inflammatory marker levels between groups before AMI reperfusion. In the control group, IL-6 level increased from 112.6 ± 11.2 pg/mL before AMI reperfusion to 463.4 ± 32.6 pg/mL at 24-h follow-up, which was higher than that in the HUCMSC group (463.4 ± 32.6 vs. 326.7 ± 22.8 pg/mL, P < 0.001) and HUCMSC+CsA group (463.4 ± 32.6 vs. 256.3 ± 16.7 pg/mL, P < 0.001). The levels of IL-6 declined gradually at the other two follow-up times and there were no significant differences among groups (Fig. 8A). Changes in IL-6 were similar to those in CRP levels for each group. CRP was lower in the HUCMSC+CsA group than in the HUCMSC group at 24-h follow-up (12.5 ± 4.2 vs. 18.3 ± 3.9 μg/mL, P < 0.01) (Fig. 8B). The levels of IL-10 increased and reached a maximum at 1-week follow-up in each group. In the HUCMSC group, the level of IL-10 was higher than that in the control group (76.5 ± 10.4 vs. 43.5 ± 8.6 pg/mL, P < 0.001) and HUCMSC+CsA group (76.5 ± 10.4 vs. 32.5 ± 6.9 pg/mL, P < 0.001) at 24-h follow-up. The level of IL-10 was lower in the HUCMSC+CsA group than in the HUCMSC group (74.2 ± 10.2 vs. 112.4 ± 12.3 pg/mL, P < 0.001) and the control group (74.2 ± 10.2 vs. 96.4 ± 10.2 pg/mL, P < 0.01) at 1-week follow-up (Fig. 8C).
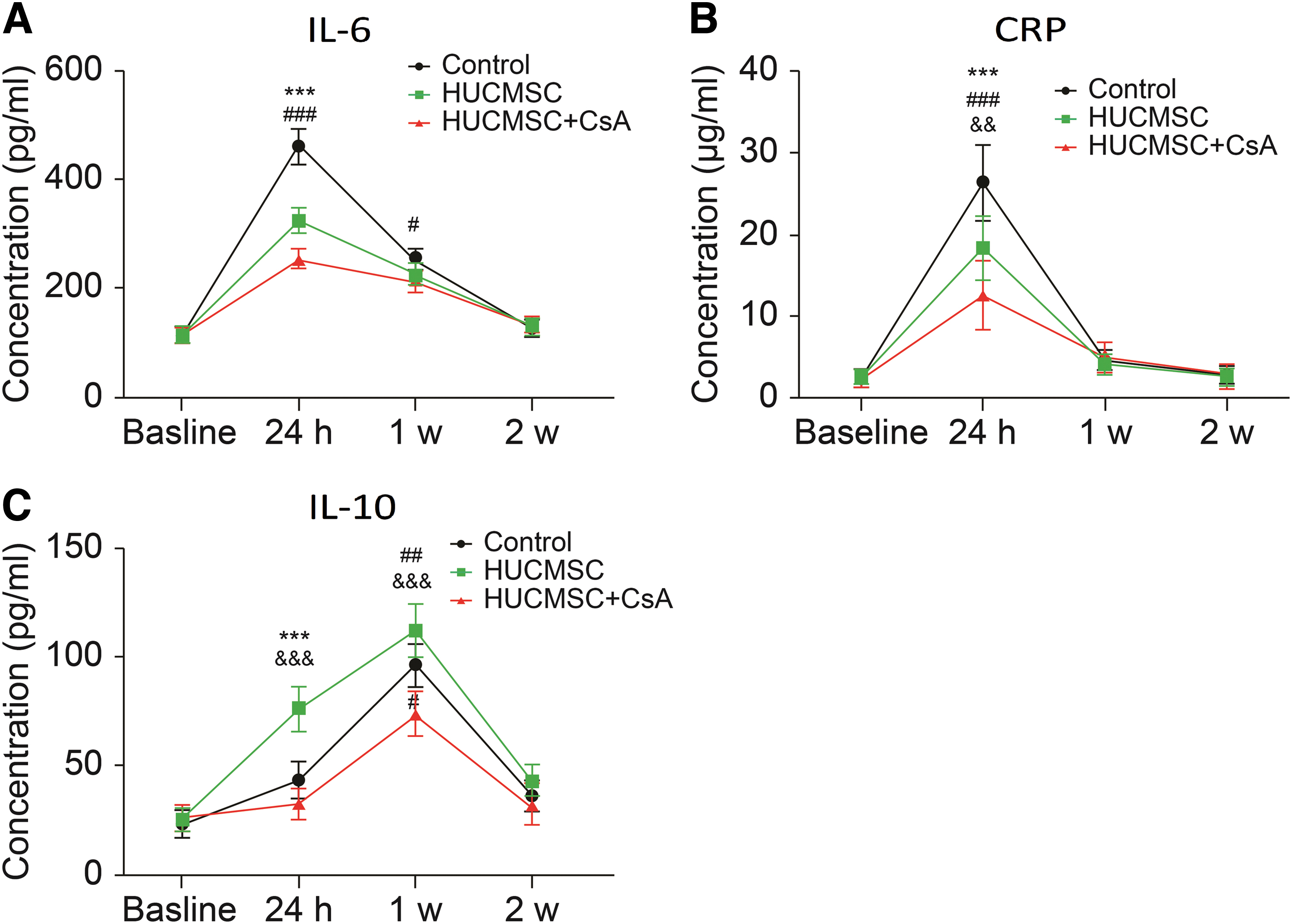
Serum inflammatory marker levels.
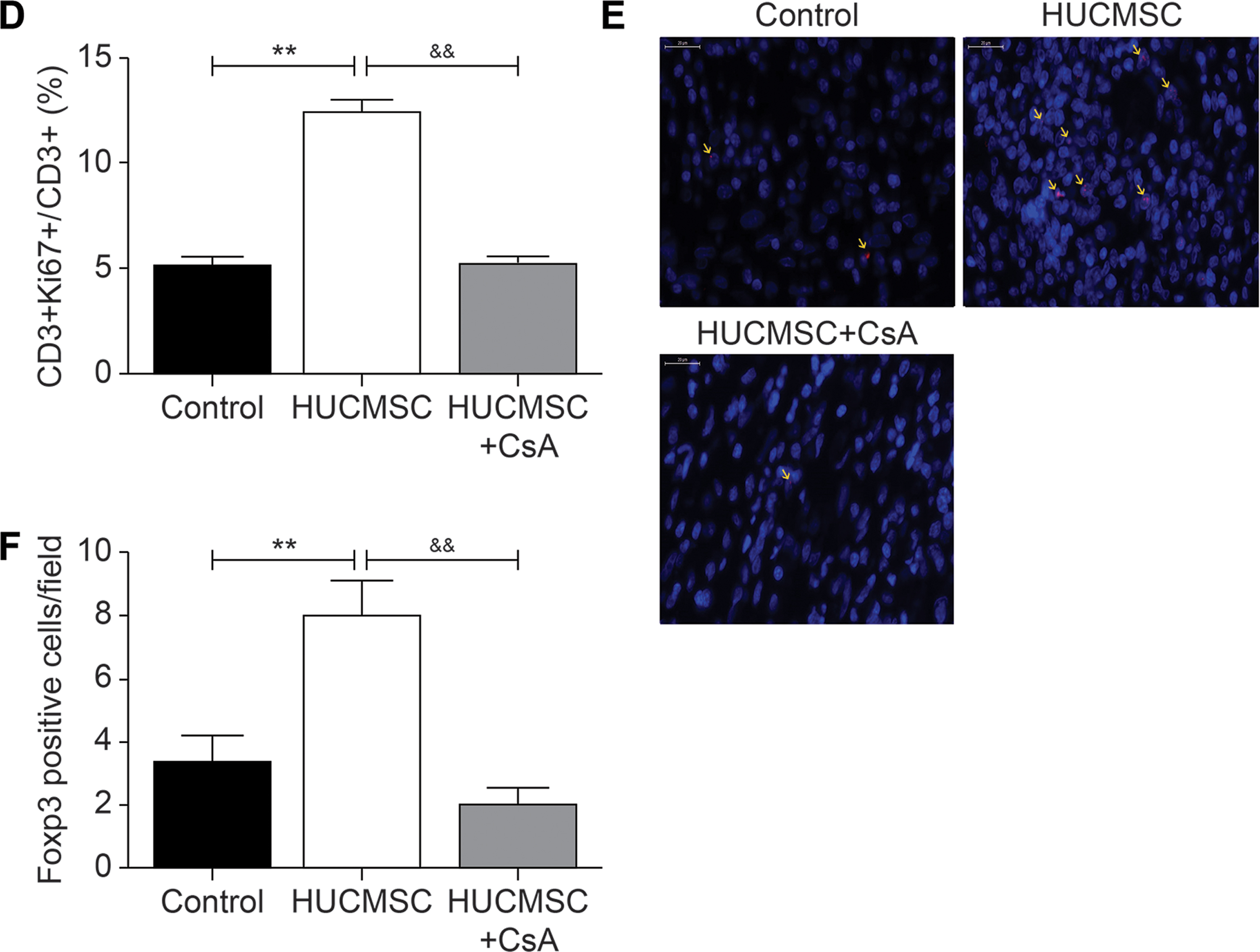
Infiltration and proliferation of T lymphocytes in myocardial infarction and border areas
Immunohistochemistry analysis revealed more presence of CD3+ cells in the infarct area of the HUCMSC group than in the control group (70.4 ± 7.5 vs. 38.2 ± 6.2, P < 0.001), whereas addition of CsA reduced the local infiltration of T lymphocytes (35.2 ± 4.8 vs. 70.4 ± 7.5, P < 0.001). There was no significant difference in the border zone among all groups (Fig. 9A, B). The proportion of CD3+- and Ki67+- positive cells among CD3+ cells was detected by double-labeling immunofluorescence to represent the proliferation rate of T lymphocytes. The results also showed a greater presence of CD3+ cells in the infarct area of the HUCMSC group. The results showed that the proliferation rate of T lymphocytes was higher in the HUCMSC group than in the control group (12.4% ± 1.1% vs. 5.1% ± 0.6%, P < 0.01) and HUCMSC+CsA group (12.4% ± 1.1% vs. 5.2% ± 0.6%, P < 0.01) (Fig. 9C, D). The number of Foxp3+ cells in the infarct area of the HUCMSC group was higher compare with the control group (8% ± 1.2% vs. 3.3% ± 1.5%, P < 0.01), whereas addition of CsA reduced the number to 2% ± 0.5% (P < 0.01) (Fig. 9E, F).
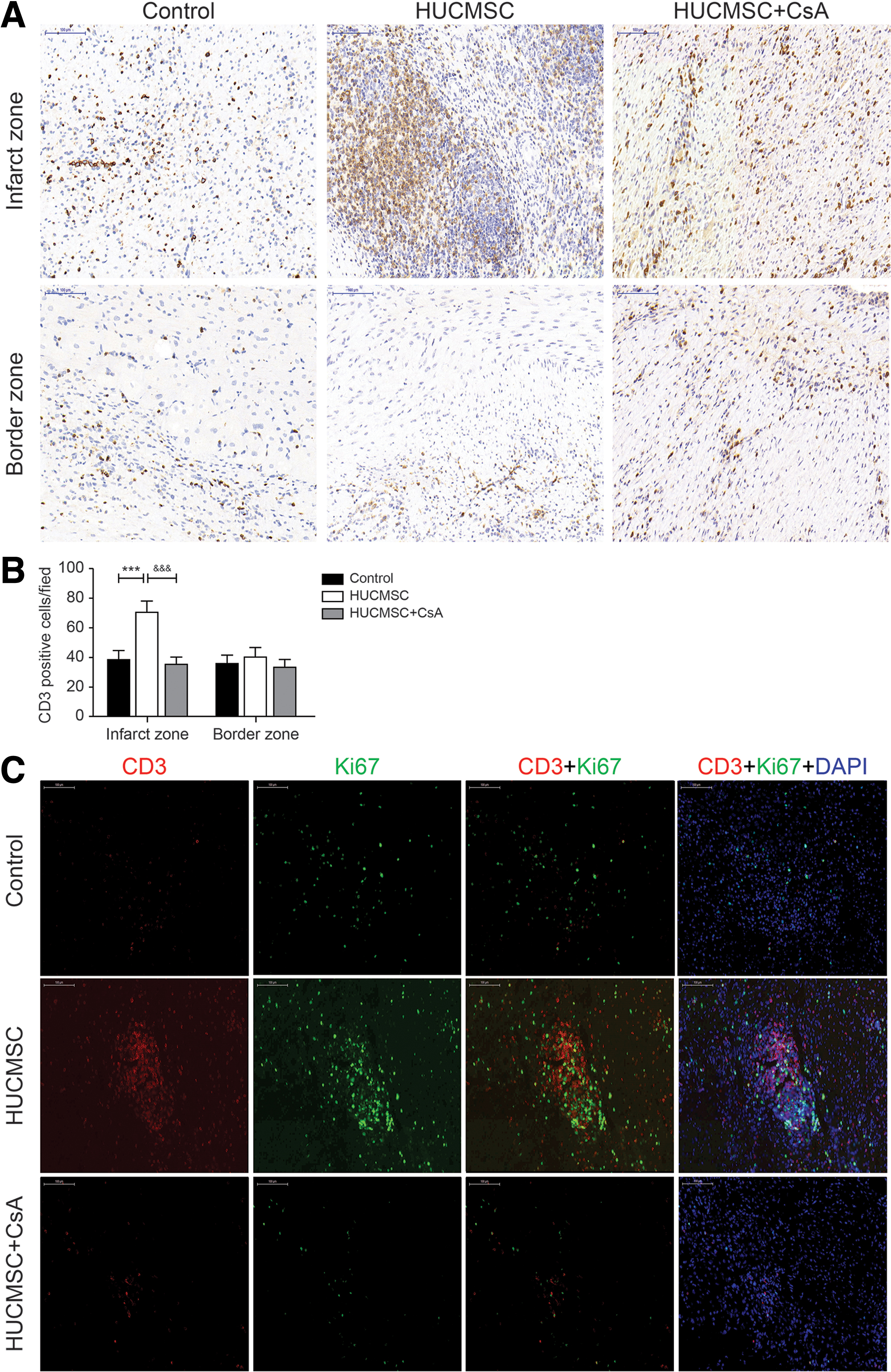
Infiltration and proliferation of T lymphocytes in the myocardial infarction and border areas.
Paracrine effects of HUCMSCs on T lymphocytes in vitro
The levels of IL-10 and TGF-β1in the supernatants were determined by ELISA. There were no significant differences between two groups on day 1. However, on the third day, both IL-10 and TGF-β1 were obviously increased by HUCMSCs in the Transwell experiments (Fig. 10A). PBMCs were stained with CD3, CD4, and CD8 to evaluate the effects of HUCMSCs on T cell subsets. Although there were no significant differences between groups with regard to the percentage of CD4+/CD8+T cells, a higher frequency of CD4+T cells was found in the HUCMSC co-culture group (Fig. 10B). The expression level of Treg markers was also determined by flow cytometry. On the first day of co-culture, the frequency of CD4+CD25+ T cells was significantly increased in the co-culture group, which was maintained by prolonging the stimulation time of HUCMSCs for 3 days (Fig. 10C).

Paracrine effects of HUCMSCs on T lymphocytes in vitro.
Discussion
In this study, we investigated the effect of HUCMSC transplantation on reducing myocardial IRI when transplantation is performed immediately following reperfusion. The mechanism of myocardial IRI is considered to be related to an increase in free radical production, local myocardial calcium overload, and the inflammatory process [20]. Measures for preventing IRI include ischemic preconditioning, ischemic postconditioning, regulation of the production of oxygen free radicals, and anti-inflammatory drugs [21,22]. MSCs show bidirectional regulation of inflammation and have a strong immunosuppressive function when the inflammatory response is intense [12,23]. HUCMSCs have some advantages over other types of MSCs, including a greater ability to proliferate and differentiate, lower immunogenicity, and more convenient sampling [10,24]. Therefore, HUCMSCs were used in this study.
In terms of the timing of transplantation, some studies suggested that 2 weeks after reperfusion is the ideal time because the local inflammatory response has been alleviated by this point and the local microenvironment is more suitable for the survival of transplanted cells [25,26]. The initiation of reperfusion injury appeared at the time of reperfusion, and the inflammatory response was most intense at this time. After 1 week, the inflammatory response in the reperfusion injury area gradually subsided. However, as mentioned above, MSCs have a bidirectional effect on inflammation regulation. In the repair period, MSCs could not exert their immunosuppressive function. Therefore, from an inflammatory perspective, early cell transplantation may be appropriate. A clinical trial showed that treatment with autologous bone marrow-derived mononuclear cells either 5–7 days or 3–4 weeks after AMI did not improve LV function at 12 months [27]. To confirm this, we performed HUCMSC transplantation through a coronary artery at the time of immediate reperfusion after myocardial infarction when the acute inflammatory response is intense to evaluate its protective effect on myocardial IRI. In addition, from the perspective of clinical practice, cardiac intervention 1 week after emergency percutaneous coronary intervention increases the risk of related complications and the economic and mental burden on patients. In terms of clinical application, improving rapid cell resuscitation technology provides feasibility for clinical application.
In addition, we used pigs as experimental animal models because they are highly consistent with humans with respect to the anatomy and physiology of the cardiovascular system, and HUCMSC xenotransplantation experiments have also been reported previously [28]. The special immunological privileged status of MSCs offers the possibility of xenotransplantation.
In this study, there was no case of postoperative death in the HUCMSC group, but one case in the HUCMSC+CsA group and two cases in the control group. Although the sample size of the animal experiments was not as large as that of clinical trials, it suggested the feasibility of HUCMSC transplantation. The cause of death of these pigs was considered to be associated with malignant arrhythmia or heart failure because no cardiac rupture was found. These two complications are closely related to the infarct size.
We found that HUCMSC transplantation reduced the myocardial infarct size compared with that of the control group, whereas adding CsA before HUCMSC transplantation inhibited the protective effect, which may explain the low mortality in the HUCMSC group. In the Masson staining images, we found that collagen fibers in the infarct core area were longitudinal and transverse after HUCMSC transplantation, and the normal shape of myocardial cells was observed next to the collagen fibers, whereas collagen deposition in the control group was distributed in mesh sheets. Moreover, we found that the collagenous area in the infarct core area of the HUCMSC group was lower than that in the control group.
HUCMSC transplantation had no significant effect on cardiac structure and function compared with those in the control group, which may be correlated with the insufficient myocardial infarction area and short observation time. The myocardial infarct size in our model was moderate because we found, in the preliminary experiment, that the intraoperative mortality rate would be too high if the infarct size was too large. However, adding CsA before HUCMSC transplantation caused significant ventricular remodeling and impaired cardiac function. This may be related to the impact of local perfusion disorder after adding CsA, particularly the occurrence of MVO.
Some studies have shown that homing, proliferation, and differentiation into cardiomyocytes are mechanisms for reducing the infarct size in MSC therapy [29]. Therefore, to dynamically understand the distribution of HUCMSCs in myocardium, CMR was used to trace ferumoxytol labeling of HUCMSCs in vivo, and Prussian blue staining was performed on anatomical specimens. The results revealed no obvious low-signal shadow after stem cell transplantation at the T2* sequence, whereas Prussian blue staining results showed that multiple cell clusters appeared in the infarct core area of the HUCMSC group with surrounding blue-dye particle agglomeration. We considered that homing HUCMSCs did not reach the minimum number of cells that could be detected by MR because, in a previous study, we found that they could be traced under MR only when there were more than 5 × 104 clustered cells [15]. We observed significantly fewer Prussian blue-stained dye particles after adding CsA than those in the HUCMSC group, indicating that addition of CsA in the acute phase of myocardial ischemia reperfusion inhibits the homing of stem cells. These results suggest that the acute inflammatory environment is more favorable for HUCMSC homing. This result is consistent with those of some studies showing that stem cell homing requires the involvement of inflammatory mediators [30,31].
Through the CMR first-pass perfusion curve examination, we found that the local perfusion was partly obstructed after 24 h of postoperative cell transplantation, whereas 1 week after surgery, perfusion in the cell group was significantly recovered compared with that in the control group. However, local perfusion of the myocardial infarction was not recovered in the HUCMSC+CsA group, and there were even two cases of MVO. This is likely related to the adverse effect of complete suppression of inflammation on the passage of HUCMSCs over the microcirculation to the infarcted myocardium and continuous obstruction of the microcirculation. Because MSCs are much larger than the diameter of capillaries, the microvasculature may be a physical barrier that blocks the trafficking of exogenous MSCs. A previous study showed that 92% of intra-arterially infused expanded MSCs were arrested at the precapillary level during the first pass, with only 14% of the stem cells surviving and extravasating through the microvascular wall at 3 days following injection [32]. The homing efficiency of MSCs is closely related to various chemokines in the inflammatory environment. In our study, combined with the previous Prussian blue staining results, this finding further suggests that the weak inflammatory response induced by CsA is not conducive to the homing of HUCMSCs.
In addition, the amount of angiogenesis in the infarct and the border zone in the HUCMSC group was greater than those in the other groups. Some studies suggested that HUCMSCs promote neovascularization by secreting paracrine factors and by differentiating to endothelial cells [33]. Therefore, in our study, the increased amount of neovascularization may be correlated with the secretion of vascular growth-promoting factors by homing HUCMSCs or their direct differentiation into vascular endothelial cells, although this requires further analysis.
A significantly larger number of CD3+ cells clustered in the infarct zones of the HUCMSC group than in the control group. CD3 is the main cell surface marker of T lymphocytes. Recruitment of immune cells after AMI is a dynamic process. T lymphocytes in the myocardial infarction region after AMI gradually increased, reaching a peak at ∼7 days. After AMI reperfusion, T lymphocytes began to appear in the infarction region for several hours, reaching a peak at ∼3 days after perfusion [34]. After AMI reperfusion, MSCs can reduce the intensity of the inflammatory response by regulating the function of T lymphocytes, thereby reducing injury after ischemia reperfusion.
In this study, the number of clustered CD3+ cells in the HUCMSC+CsA group was lower than that in the HUCMSC group, and cardiac protection by HUCMSCs was prevented when CsA was added to inhibit T lymphocytes. This indicates that T lymphocytes play a crucial role by reducing IRI during the acute stage of HUCMSC therapy. HUCMSCs can recruit T lymphocytes to “inhibit” their inflammatory damage and regulate the inflammatory response, whereas T lymphocytes can increase the homing rate of stem cells. Addition of CsA inhibited the activity of T lymphocytes and then blocked the interaction between HUCMSCs and T lymphocytes, finally affecting the regulation of the inflammatory response by HUCMSCs and protective effect on myocardial IRI.
Previous studies have also found that MSCs can express IDO, iNOS, and some chemokines in the acute inflammatory stage, recruit T lymphocytes through chemokine receptors such as CXCR3 and CCR5, and exert immunosuppression through intercellular interactions [11]. After T lymphocyte recruitment, MSCs can inhibit the release of IFN-γ by Th1, promote the release of IL-10 by Th2, and reduce the inflammatory response [35]. In this study, the level of IL-10 was higher in the HUCMSC group than in the other two groups at 24-h follow-up. Serum proinflammatory markers, including IL-6 and CRP, were lower in the HUCMSC group than in the control group, showing remission of the intense inflammatory response, whereas added CsA caused more severe inhibition of the inflammatory response and eliminated cardiac protection. These results showed that regulating the inflammatory response rather than simply blocking inflammatory responses may be important.
We also found that Prussian blue-stained particles were distributed in the area surrounding T lymphocyte clusters, revealing a close relationship between transplanted HUCMSCs and T lymphocytes. To explore the mechanism of the interaction of HUCMSCs and T lymphocytes in an inflammatory environment, a novel Transwell co-culture model of PBMCs and HUCMSCs was used. The release of anti-inflammatory cytokines IL-10 and TGF-β1 was increased from the first 24 h of co-culture, which was approximately consistent with the detection of serum cytokines in animal experiments. This result also indicates that this intercellular interaction was realized through a paracrine mechanism to regulate inflammatory responses. These results showed that cardiac protection by HUCMSCs was triggered by cell-cell contact with T lymphocytes to regulate the inflammatory process.
In the cell model, CD4/CD8 among PBMCs was increased by co-culture with MScs, although the results were not significant, demonstrating the promotion effects of MSCs on the generation of CD4+ T lymphocytes. In the cell proliferation experiment, the proliferation rate of T lymphocytes was higher in the HUCMSC group than in the other two groups. Two weeks after myocardial infarction reperfusion is the myocardial repair phase, during which T lymphocytes are mostly anti-inflammatory T lymphocytes, such as Th2 or Treg cells. This result suggests that HUCMSCs promoted the activation of anti-inflammatory T lymphocytes.
Treg cells include CD4+CD25+Foxp3+Treg, NK Treg, CD8+ Treg, and Th3 cells, among which CD4+CD25+Foxp3+Treg is the main subtype. Transgenic animal experiments suggested that eliminating Treg cells increases the degree of myocardial injury, and local infusion of Treg cells is beneficial for protecting an ischemic myocardium [36]. MSCs promoted the generation of CD4+CD25+Foxp3+Tregs significantly in indirect co-culture experiments. Our study revealed more Foxp3-positive cells in the infarct area after HUCMSC transplantation than in the control group. The number of Foxp3-positive cells was decreased when CsA was added. This result showed that regulating rather than completely suppressing the acute immune response was beneficial for subsequent myocardial repair after IRI.
One limitation of this study is that the differentiation of homing cells was not further analyzed. Some studies demonstrated that an intercellular interaction between stem cells and T lymphocytes is achieved through a paracrine mechanism [37,38]. In this study, whether T lymphocytes influence the differentiation of homing HUCMSCs through a paracrine mechanism in vivo was not explored. Another limitation of this study is that the pure effects of CsA on myocardial IRI were not explored. CsA is an immunosuppressive agent that can influence the inflammatory process after myocardial IRI.
In general, immediate coronary artery transplantation of HUCMSCs can reduce AMI reperfusion injury and promote myocardial repair through the interaction of T lymphocytes. Added CsA diminishes cardiac protection by HUCMSCs, by blocking the function of T lymphocytes. Further studies are required to explore the mechanisms of T lymphocyte involvement in this process.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported, in part, by the Excellent Youth Foundation of Nanjing Scientific Committee (JQX15002).