
Abstract
Treatment options for neurodevelopmental disorders such as schizophrenia and autism are currently limited. Antipsychotics used to treat schizophrenia are not effective for all patients, do not target all symptoms of the disease, and have serious adverse side effects. There are currently no FDA-approved drugs to treat the core symptoms of autism. In an effort to develop new and more effective treatment strategies, stem cell technologies have been used to reprogram adult somatic cells into induced pluripotent stem cells, which can be differentiated into neuronal cells and even three-dimensional brain organoids. This new technology has the potential to elucidate the complex mechanisms that underlie neurodevelopmental disorders, offer more relevant platforms for drug discovery and personalized medicine, and may even be used to treat the disease.
Introduction
Schizophrenia and autism are two neurodevelopmental disorders that share a common etiology and pathology. Currently, these diseases are poorly treated by FDA-approved drugs. While antipsychotics are available for schizophrenia, these drugs do not work for everyone, can have serious adverse side effects, and have little impact on some of the most devastating symptoms of the disease. The outlook for autism is even more concerning as there are currently no FDA-approved drugs to treat the core symptoms of the disorder. To find new and more effective treatment options, a better understanding of the neural mechanisms of schizophrenia and autism is needed.
Our understanding of these complex disorders has been limited by the fact that they are difficult to model in animals. Although specific pathogenic mechanisms, for example, prenatal infection or genetic mutations, can be recapitulated in rodents [1,2], many of the symptoms associated with neurodevelopmental disorders are uniquely human, for example, hallucinations and delusions [3]. Furthermore, some of the brain regions implicated in neurodevelopmental disorders, including the prefrontal cortex, are much more complex in humans than rodents [4]. Therefore, the translational relevance of the neurobiological mechanisms underlying specific behaviors that can be modeled in rodents remains unclear. In humans, ethical considerations restrict the types of experiments that can be conducted in patients.
However, recent advancements in stem cell technology have allowed the reprogramming of adult somatic cells into induced pluripotent stem cells, which can then be differentiated into a variety of cell types, including neurons. This new technology is exciting as it will not only allow us to understand the neural mechanisms that underlie neurodevelopmental disorders but it also offers better platforms to test the efficacy of treatment strategies and may even be used to treat the disease itself (Fig. 1).
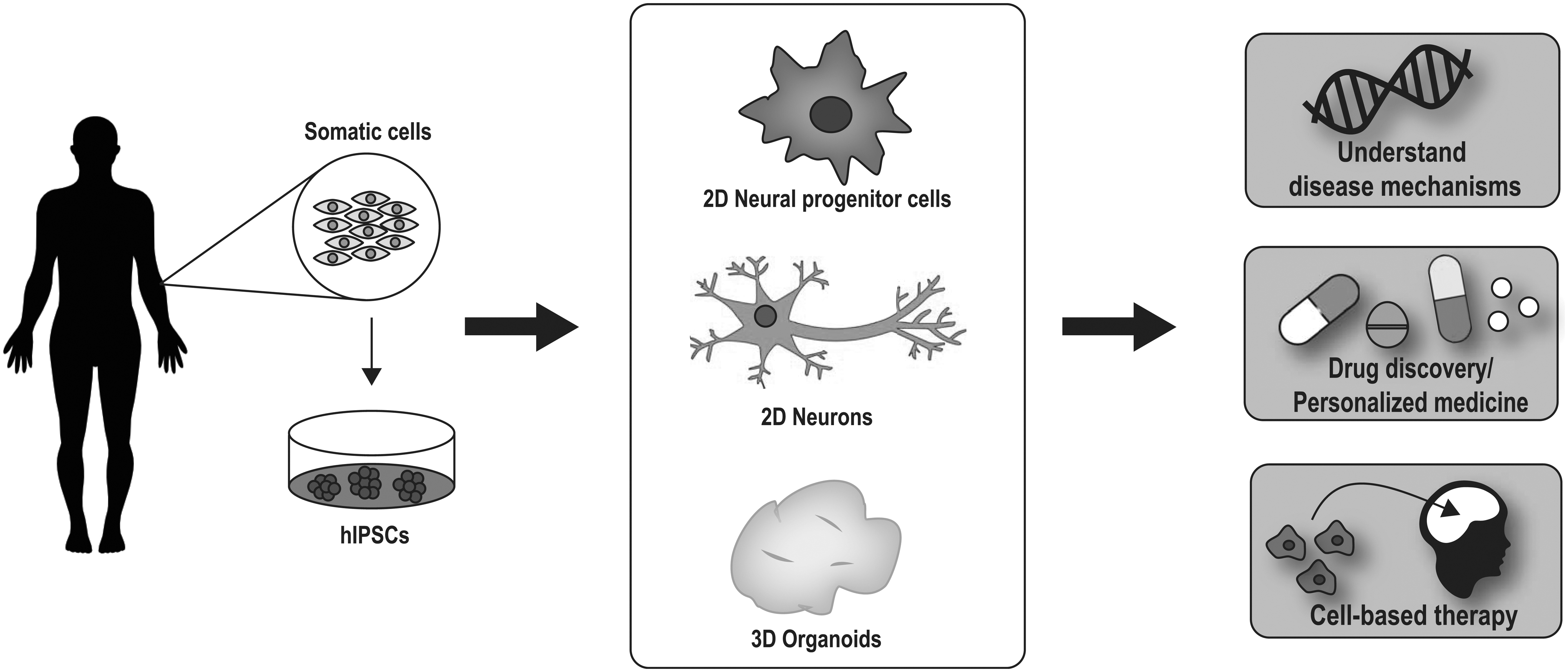
Stem cells can be used to understand neurobiological mechanisms, as drug discovery platforms, and for cell-based therapies. Somatic cells from patients can be converted to hIPSCs, which can then be differentiated into a variety of cell types, including neural progenitor cells, neurons, and even 3D brain organoids. These cells and organoids hold the potential to (1) provide insight into mechanisms underlying complex disorders; (2) be used as drug discovery platforms or for personalized medicine; and (3) even be used as therapeutics to treat neurodevelopmental disorders such as schizophrenia and autism. 3D, three-dimensional; hIPSCs, human induced pluripotent stem cells.
Schizophrenia
Schizophrenia is a devastating psychiatric disorder that usually presents in early adulthood and affects ∼1% of the population [5]. Positive symptoms such as delusions and hallucinations are often the most striking features of the disorder; however, schizophrenia patients also display characteristic negative and cognitive symptoms, which can be severely debilitating. Negative symptoms, such as blunted affect, emotional withdrawal, and social avoidance, and cognitive symptoms, including disruptions in working memory, attentional deficits, disorganized thought, and cognitive inflexibility, can negatively influence social and occupational functioning and diminish quality of life [6 –8]. Currently prescribed medications for schizophrenia have been classified into first- (typical) and second-generation (atypical) antipsychotics [9]. Both classes not only have a primary effect as antagonists at the dopamine D2 receptor [10] but second-generation antipsychotics also target the serotonin system [11]. While the second-generation drugs have a reduced side effect profile, unwanted side effects, including extrapyramidal or metabolic symptoms, remain, resulting in a high rate of discontinuation [12]. Furthermore, while both typical and atypical antipsychotic drugs can be effective in treating positive symptoms, they have little to no impact on negative and cognitive symptoms of schizophrenia [12]. Even among patients who do respond to antipsychotic treatment, disability rates remain high and functional outcomes poor, highlighting the need to treat residual negative and cognitive symptoms [6]. Clearly, there is an urgent need for new and more effective treatment strategies for schizophrenia.
Development of new and more effective antipsychotics will require a better understanding of the pathological mechanisms that underlie schizophrenia. Schizophrenia is a heterogeneous disorder that is thought to result from a complex interaction between genetic and environmental factors. Twin studies have demonstrated that schizophrenia is highly heritable, with heritability estimates around 80% [13,14]. Genome wide association studies (GWAS) have identified multiple common variants, but individual loci do not confer a large risk for developing schizophrenia [15]. Similarly, several risk factors have been associated with development of schizophrenia, including increased paternal age [16,17], complications during pregnancy (eg, maternal infection and malnutrition) [18 –23], childhood adversity [24], and drug abuse [25 –27].
Imaging and postmortem brain studies have implicated specific brain regions, cell types, and neurotransmitters in the pathology of schizophrenia. For example, schizophrenia patients show overall reductions in brain volume [28] with specific losses in regions such as the prefrontal cortex, hippocampus, thalamus, and amygdala [29 –31]. Functional neuroimaging studies confirm these findings, demonstrating altered activity associated with specific symptom domains. One of the more consistent observations in schizophrenia patients is an increase in hippocampal activity at rest [32 –35] and this increase in hippocampal activity has been correlated with the severity of positive symptoms [36].
Furthermore, multiple neurotransmitter systems have been implicated in the pathology of schizophrenia. One of the first hypotheses of schizophrenia, the dopamine hypothesis, suggested that hyperactivity in the mesolimbic dopamine system was responsible for positive symptoms. Dopamine receptor agonists have been shown to induce schizophrenia-like symptoms in otherwise healthy individuals [37] and all currently available antipsychotics act as antagonists at the dopamine D2 receptor [10]. Imaging studies demonstrate that striatal dopamine synthesis is elevated in schizophrenia patients [38,39]. However, this theory is limited by the fact that no overt pathology has been identified in the dopamine system itself and that changes in dopamine signaling do not account for all symptom domains. More recently, the GABAergic system has been implicated in the pathology of schizophrenia [40,41]. Postmortem studies consistently demonstrate reductions in the GABA-synthesizing enzyme, GAD, and a loss of specific interneuron subtypes in the hippocampus [42 –45] and prefrontal cortex [46,47]. Conversely, the excitatory neurotransmitter, glutamate, has also been implicated in the pathology of schizophrenia. Agonists of the glutamatergic NMDA receptor, including ketamine, have been shown to induce a psychosis-like state [48]. Although the results have been somewhat inconsistent, there is also evidence for changes in glutamatergic cell numbers, dendritic arborization, and receptor expression [49]. Together, these studies provide clues into the pathogenic mechanisms that underlie schizophrenia, but a comprehensive understanding of the disease and effective treatment strategies remain to be established.
Autism
Autism spectrum disorders (ASDs) include several developmental disorders that are usually diagnosed within the first or second year of life. ASDs are characterized by three core symptom domains: deficits in social interaction, impairments in communication, and repetitive stereotyped patterns of behavior or thought [50]. Along with varying degrees of dysfunction in these core behavioral domains, many autistic patients also experience other symptoms, including intellectual disabilities, disruptions in motor coordination, and gastrointestinal disturbances [51]. This heterogeneous group of disorders affects ∼1 in every 59 US children [52], yet there are no drugs currently available for the treatment of ASD. Rather, pharmacological interventions only target comorbid symptoms, and these treatments often have serious adverse side effects [53]. Therefore, new therapeutic strategies targeting the core behavioral deficits associated with ASD are urgently needed.
Like schizophrenia, autism is thought to result from a complex interplay between genetic and environmental factors. Twin studies have demonstrated that the heritability of autism is up to 90% [54]. However, highly penetrant copy number variants remain relatively rare and more common individual gene variants do not significantly increase the risk of developing autism [55]. Environmental risk factors have also been associated with ASD, including prenatal infection [56,57] and exposure to drugs such as valproic acid (VPA) [58,59].
Postmortem and imaging studies have begun to identify brain regions and neurotransmitter systems involved in autism. Structural imaging studies have demonstrated that autism is associated with an increase in brain volume during early childhood, specifically in regions such as the cerebellum, cerebrum, amygdala, and hippocampus, but this effect is often lost by late childhood or adolescence [60,61]. Functional imaging studies have also identified changes in neural activation associated with specific behavioral symptoms. For example, deficits in social perception have been associated with reduced activity in the amygdala [62,63].
Alterations in inhibitory/excitatory balance have also been implicated in the pathology of autism [64]. For example, copy number variants have been identified on the 15q11-q13 chromosome, a locus that contains genes related to GABA function, including multiple GABAA receptor subunits. Mutations in these GABAA genes are associated with an increased risk for autism [65,66]. Furthermore, mutations in other genes associated with ASD, such as CNTNAP2 and PLUAR, have been shown to decrease the number of GABAergic interneurons in the cortex [64]. Postmortem and imaging studies in autistic patients demonstrate reductions in the GABAA receptor [67,68], the GABA-synthesizing enzymes, GAD65 and GAD67 [69], and in GABA itself [70]. A recent analysis of multiple genetic and environmental mouse models of autism suggests that these animals have reduced GABAergic interneurons [71], further implicating the GABAergic system in the pathology of autism.
Stem Cells for Modeling Psychiatric Disease
The recent discovery that transcription factors can be used to reprogram adult somatic cells into human induced pluripotent stem cells (hIPSCs), cells that maintain the properties of embryonic stem cells, has led to the advent of a new era of stem cell research [72]. Indeed, research from this rapidly growing field has found that hIPSCs can be generated from patient fibroblasts, keratinocytes [73], hair follicles [74], blood [75], and other tissues [76]. The development of integration-free viruses, plasmids, or small molecules now ensures that the reprogrammed hIPSCs contain the entire genetic background of the donor [77], which is particularly important for neurodevelopmental disorders with a strong genetic component, including schizophrenia and autism [15,54]. These pluripotent cells have the ability to differentiate into a variety of cell types that have been used to study neurodevelopmental disorders, such as autism and schizophrenia. Neural progenitor cells have been shown to have a similar gene signature to hIPSC-derived neurons [78], but can be grown much more rapidly. More recently, three-dimensional (3D) brain organoids have been used to study neurodevelopmental disorders. This model system has been shown to express a transcription factor profile that models that of the developing human brain [79 –81]. Within 3D brain organoids, cells have a high degree of organization, including fluid-filled cavities that resemble ventricles surrounded by defined proliferative zones [82]. Recently, protocols have been developed to direct the differentiation of organoids into specific brain regions, including the cerebral cortex [79,83], midbrain [84,85], and subcortical regions [83]. Together, two-dimensional (2D) and 3D cell culture systems have already begun to provide insight into the neurobiological mechanisms underlying schizophrenia and autism (Table 1).
Stem Cells to Understand the Neurobiology of Schizophrenia and Autism
ASD, autism spectrum disorder; CHD8, chromodomain helicase DNA-binding protein 8; DISC1, disrupted in schizophrenia 1; hIPSC, human induced pluripotent stem cell; RNASeq, RNA sequencing.
Schizophrenia
As the use of stem cells for understanding neurodevelopmental disorders grows, many studies have focused on confirming the validity of the model by corroborating and extending data found in patient studies. One of the most common findings from GWAS and postmortem studies of schizophrenia patients is a loss of synaptic proteins and dendritic spines [86]. In one of the first uses of stem cells to model schizophrenia, results from human studies were confirmed using neurons differentiated from schizophrenia hIPSCs. Rabies virus was used to demonstrate that neurons from schizophrenia hIPSCs had reduced synaptic connectivity. This effect was reversed by treatment with loxapine, a typical antipsychotic drug. Furthermore, the neurons derived from schizophrenia hIPSCs also showed reduced dendritic arborization and fewer neurites. RNA sequencing (RNASeq) demonstrated that the schizophrenia hIPSC-derived neurons had changes in the expression of genes related to glutamate, cAMP, and WNT signaling and these transcriptional changes could also be reversed by treatment with loxapine [87]. Similar results were obtained in neural progenitor cells (NPCs) derived from schizophrenia hIPSCs. RNASeq demonstrated differences in glutamate receptor signaling, insulin signaling, neuronal migration, neuron differentiation, synaptic transmission, and synaptic vesicle function [78]. Around the same time, Chiang et al. generated hIPSCs from patients with a mutation in the disrupted in schizophrenia 1 (DISC1) gene [88], which has been strongly associated with schizophrenia as well as other psychiatric disorders [89,90]. Consistent with the findings above, neurons from DISC1 mutant hIPSCs had reduced expression of SV2+, a synaptic vesicle protein that has been associated with mature synaptic vesicles and regulates presynaptic release [91]. Furthermore, patch clamp recordings found decreased frequency of spontaneous excitatory synaptic currents, with no change in amplitude, suggesting a presynaptic mechanism. FM1–43 imaging was used to show that the DISC1 mutant neurons had a significant defect in depolarization-induced vesicle release. These deficits were reversed when the DISC1 mutation was corrected and could be induced in hIPSCs from healthy controls by mutating DISC1, suggesting that the effects are a direct result of the DISC1 mutation. In addition, RNASeq identified alterations in genes related to synaptic transmission, nervous system development, and dendritic spines [91]. Together, these studies point to specific mechanisms contributing to disrupted synaptic transmission in schizophrenia, which are in line with findings from schizophrenia patients [86].
Schizophrenia has also been associated with deficits in cellular plasticity and migration [92]. In a study of neurons derived from schizophrenia hIPSCs, RNASeq demonstrated that neurons from schizophrenia hIPSCs had increased expression of genes associated with organization of the extracellular matrix and cell adhesion [93]. In NPCs derived from schizophrenia hIPSCs, changes were also found in cellular adhesion, cytoskeletal remodeling, and oxidative stress pathways. Furthermore, NPCs from schizophrenia patients had reduced migration and outgrowth, an effect that was not rescued by loxapine or clozapine [78]. These results may be associated with alterations in WNT signaling, which has been associated cell migration [94] and is disrupted in schizophrenia (SZ) [95]. NPCs derived from schizophrenia hIPSCs demonstrate altered WNT signaling [96].
A loss of hippocampal and cortical interneurons has also been observed in postmortem brains of schizophrenia patients [42 –47]. To test interneuron function, cortical interneurons were derived from hIPSCs from schizophrenia patients. The cells were transplanted into mice, where they survive, express interneuron markers, and display firing patterns that resemble that of endogenous interneurons. Furthermore, the cells integrate into circuitry, as evidenced by spontaneous excitatory post-synaptic potentials (sEPSPs), and optogenetic activation of transplanted interneurons increases inhibitory postsynaptic potenials (IPSPs) in nontransplanted cells. Further examination of these cells, however, demonstrated that the schizophrenia-derived interneurons had decreased the neurite number from soma, total branch number, neurite length, and inhibitory synapse number. RNASeq found that although interneuron markers, such as GAD1, VGAT, SST, and Lhx6, did not differ between control and schizophrenia-derived interneurons, protocadherin (PCDHA) gene expression was decreased. PCDHA gene expression is also altered in schizophrenia patients [97]. Using a PCDHA knockout (KO) mouse, the authors demonstrated that similar to the transplanted interneurons, the PCDHA KO mice had changes in interneurons in the prefrontal cortex, including a decreased neurite number from soma, decreased total branch number, and decreased neurite length. These findings are in line with postmortem brain tissue in which the interneuron neurite number from soma, total branch number, and total neurite length are decreased in the medial prefrontal cortex (mPFC) of schizophrenia patients [98].
So far, few published studies have used hIPSCs from schizophrenia patients to generate 3D brain organoids. However, a recent study found that cortical organoids from schizophrenia patients show disorganization of proliferating cells within ventricular zones. Interestingly, the schizophrenia-derived organoids also had a reduction in mature neurons in cortical areas, while at the same time showing an increase in mature neurons in subcortical areas. Nuclear fibroblast growth factor receptor 1 (nFGFR1) has been implicated in neuronal differentiation and schizophrenia-derived organoids that express less nFGFR1 in cortical zones [99]. Together, these results suggest that NPCs, neurons, and even cerebral brain organoids may be used to model pathological mechanisms associated with schizophrenia.
Autism
To date, studies using stem cells to model autism have used hIPSCs derived from patients with both idiopathic and syndromic forms of the disorder. Rett syndrome is a form of autism caused by mutations in the X-linked MeCP2 gene [100]. In one of the first studies to use stem cells to understand autism, neurons were derived from hIPSCs from patients carrying the MeCP2 mutation. As has been observed in postmortem tissue from autism patients [101], hIPSC-derived neurons with the MeCP2 mutation had a reduction in excitatory synapses. Knockdown of MeCP2 from control hIPSC-derived neurons produced a similar effect, while MeCP2 overexpression increased excitatory synapses. Furthermore, hIPSC-derived neurons from Rett syndrome patients had fewer dendritic spines, smaller cell bodies, and reduced activity, as measured by calcium transients and spontaneous postsynaptic currents (both spontaneous excitatory postsynaptic potentials [sEPSCs] and spontaneous inhibitory postsynaptic potentials [sIPSCs]) [102]. Similar results were found in another study comparing hIPSC-derived neurons from autism patients with X-linked neuroligin mutations. hIPSC-derived neurons with the NLGN4 delta E 4 (skipped exon 4 splice variant) mutation had an impaired ability to form synapses [103]. Neurons derived from ASD-hIPSCs also form fewer excitatory synapses, as evidenced by a reduction in synapsin and VGLUT1 puncta. This resulted in mature neuronal networks that had less bursting [93]. As described above, the excitatory/inhibitory balance seems to be disrupted in autism [104], so it is not surprising that hIPSC-derived neurons from autism patients had less GABA and reduced expression of GABAergic receptors [102]. Using cocultures, Russo et al. suggest that these defects in synapse formation may be a direct result of deficits in astrocyte function. Neurons derived from hIPSCs from autism patients had a significant decrease in synaptic proteins, reduced glutamate release, and reduced spontaneous firing rate, which was caused by astrocytes derived from autism patients. Astrocytes from autism patients released more IL-6, a proinflammatory cytokine, and inhibiting IL-6 restored synaptogenesis in hIPSC-derived neurons from autism patients [105].
Stem cells from autism patients have also been used to generate 3D brain organoids. hIPSCs from autism patients were used to generate telencephalic organoids, and RNASeq demonstrated downregulation of genes associated with vascular development and lipid metabolism and upregulation of genes associated with synaptic transmission, gated channel activity, transcription factors associated with determining neural cell fate, and precursor cell proliferation [106]. In another study, the CRISPR-Cas9 gene editing system was used to generate CHD8 (chromodomain helicase DNA-binding protein 8) ± lines from control hIPSCs. CHD8 is a candidate gene in autism, as determined by multiple exome-sequencing studies [107 –109]. Using cerebral organoids derived from these cell lines, RNASeq identified differential expression of genes in pathways associated with nervous system development, neurogenesis, neuron differentiation, forebrain development, axon guidance, and WNT-beta catenin signaling [110].
Changes in excitatory/inhibitory balance have also been observed in 3D organoids derived from autism hIPSCs. Morphological analysis of neuronal maturation and synapse formation demonstrated an increased density of neurons and an increase in total synapse number in autism organoids. This effect may be driven by changes in GABAergic cells as there was an increase in the number of GABAergic synapses and an upregulation of the expression of transcription factors associated with the development of GABAergic cells [106]. Of the differentially regulated genes, FOXG1 is an important transcription factor for development of the telencephalon [111,112] and loss-of-function mutations have been found in patients with Rett syndrome [113,114]. FOXG1 downregulation using lentiviral vectors expressing FOXG1 small hairpin RNA (shRNA) decreased expression of markers of GABAergic cells without affecting transcription factors associated with general forebrain development or those associated with excitatory neuron differentiation. Interestingly, there was a positive correlation between autism symptom severity and FOXG1 expression in patients from whom the hIPSCs were derived, suggesting that this transcription factor may contribute to the pathology of autism through disruption of inhibitory cell function [106]. In another recent study using brain organoids to model autism, migration of GABAergic cells into cortical tissue was examined. Birey et al. generated cortical and subcortical organoids from patients with Timothy syndrome, a severe neurodevelopmental disorder characterized by autism and epilepsy. Cortical and subcortical organoids were fused and the migration of GABAergic interneurons into the cerebral cortex was analyzed. They found that in the Timothy syndrome organoids, there was an increase in saltation frequency, but the saltation length and speed were reduced, suggesting that interneurons migrated less efficiently [83]. Together, these results confirm that autism is associated with synaptic changes and excitatory/inhibitory balance and provide potential mechanisms by which these deficits may occur.
Stem Cells for Drug Discovery/Personalized Medicine in Psychiatric Disease
Currently, in vitro drug discovery, even for mental illnesses, uses immortalized cell lines and biological readouts, such as receptor binding assays [115]. Unsurprisingly, the success rate for psychiatric drug discovery is notoriously low [116], leading drug companies to focus on creating iterations of currently prescribed medications with reduced side effect profiles and increased adherence. With improvements in hIPSC technology, drug discovery may take a more personalized approach, using cells from patients who actually have the disease. For example, hIPSC-derived neurons have been used to identify novel therapeutic targets for the treatment of amyotrophic lateral sclerosis (ALS) [117,118] and Parkinson's [119]. Although not currently in practice, the research described below suggests that stem cells from patients may even be used for personalized medicine to find the most effective treatment strategy for a particular individual.
Schizophrenia
As described above, neural progenitor cells have the benefit of being relatively easy to culture and faithfully recapitulating the transcriptional profile of schizophrenia [78]. In a study by Readhead et al., NPCs from control and schizophrenia patients were compared with the immortalized cancer cell lines typically used in drug discovery. Using in silico drug screen methodologies, the authors screened drugs based on their ability to alter the transcriptome of patient-derived NPCs or immortalized cancer cell lines. Interestingly, specific drugs altered expression of schizophrenia-related genes only in schizophrenia-derived NPCs when compared with control NPCs and immortalized cancer cell lines. The authors identified 18 drugs that were able to attenuate transcriptional signatures related to schizophrenia in NPCs and differentially regulate these genes in control versus schizophrenia NPCs. These results suggest that using patient-derived NPCs to screen drugs may provide more relevant information than the immortalized cell lines used in traditional drug screens [120].
Patient-derived stem cells may also have the utility for testing the efficacy of new treatment strategies. For example, valproate, which has been proposed as an adjunct treatment in patients [121,122], has been tested on NPCs derived from schizophrenia hIPSCs. As has been observed in schizophrenia patients [123], the schizophrenia-derived NPCs showed an increase in reactive oxygen species compared with controls and valproate was able to return reactive oxygen species to control levels [124]. In another study, NPCs from a clozapine-resistant schizophrenia patient were shown to have increased concentrations of potassium and zinc. This effect was also reversed by valproate treatment [125]. Together, these results suggest that hIPSCs from patients may be used to test the efficacy of novel treatment strategies and, excitingly, may provide insight into how well individual patients will respond to different drugs.
Autism
Stem cell technology has also been used to test the efficacy of potential therapeutics for the treatment of autism. Neurons derived from hIPSCs from Rett syndrome patients were used to test two unique therapeutic strategies for autism. First, IGF1, which is currently in clinical trials for Rett and Phelan–McDermid syndromes and ASDs, was tested. IGF1 was able to increase the excitatory synapse number in hIPSC-derived neurons from Rett syndrome patients [126]. A follow-up study also demonstrated that neurons derived from autism hIPSCs treated with IGF1 have an increase in GABAergic interneurons, suggesting that this treatment may have its effect by restoring the excitatory/inhibitory balance [102]. In addition, the authors used gentamicin, an aminoglycoside antibiotic that can bind 16S ribosomal RNA (rRNA) and disrupt ribosomal proofreading, to suppress nonsense mutations that are common in Rett syndrome. Gentamicin treatment was able to increase MeCP2 protein expression and normalize the excitatory synapse number in hIPSC-derived Rett syndrome neurons [126].
SHANK3 is a synaptic protein localized to the postsynaptic density where it acts as a scaffolding protein. SHANK3 mutations have been linked to autism [127,128] and SHANK3 KO mice show autism-like behavioral deficits and reduced excitatory synapses [129]. Using hIPSC-derived neurons from control patients, compounds were screened for their ability to increase SHANK3 messenger RNA (mRNA) expression. Of these compounds, lithium, VPA, and fluoxetine significantly increased SHANK3 protein and increased colocalization between SHANK3 and the synaptic protein PSD-95. To determine if these drugs produced functional changes in neuronal activity, spontaneous calcium oscillations were measured. Lithium and VPA both increased spontaneous calcium oscillations, an effect that was blocked by SHANK3 shRNA, suggesting that the effects are dependent on this protein. Then, the authors used hIPSC-derived neurons from patients with SHANK3 mutations to determine if lithium and VPA could rescue autism-like deficits. Both drugs increased SHANK3-containing synapses and spontaneous calcium oscillations. Interestingly, one of the SHANK3 mutation patients was enrolled in a clinical trial to test the efficacy of lithium to control her mood. Lithium was able to improve her manic symptoms, ASD symptoms, and cognitive performance. Although very preliminary, these results suggest that hIPSCs may be beneficial for predicting clinical efficacy of drugs in individual patients [130].
Stem Cells as Therapy for Psychiatric Disease
One of the greatest promises of stem cell technology is the potential to cure devastating diseases such as autism and schizophrenia. With the advent of hIPSC technology, cells could be harvested from healthy relatives or patients themselves to reduce the chance of rejection. Furthermore, this strategy may allow us to actually cure diseases by replacing defective cells, rather than just by treating the symptoms. While this strategy has begun to be used in the clinic, the majority of research to date remains in animal models.
Schizophrenia
As described above, deficits in inhibitory interneuron function have been observed in postmortem tissue from schizophrenia patients, particularly in hippocampal regions. Therefore, recent studies in animal models have aimed to restore interneuron function in the hippocampus, to reduce hyperactivity in this brain region, and to alleviate the resulting behavioral correlates of schizophrenia. During fetal development, interneurons are born in subcortical regions of the brain called the medial ganglionic eminence (MGE) before they migrate tangentially into the hippocampus. Early studies in this field demonstrated the remarkable ability of MGE interneurons to survive and migrate [131] and enhance inhibition [132] when transplanted into an adult brain. Therefore, the first proof-of-concept studies transplanted fetal MGE tissue into the ventral hippocampus of rodent models of schizophrenia. Using the methylazoxymethanol (MAM) model of schizophrenia, Perez and Lodge demonstrated that MGE transplants into the ventral hippocampus reduce the pyramidal cell firing rate in the vHipp. Furthermore, these cells also normalize firing of dopamine cells in the downstream ventral tegmental area (VTA) and reduced amphetamine-induced locomotor activity, a behavioral test that is used as a correlate of positive symptoms [133]. Similarly, in a genetic model of hippocampal disinhibition, the Cyclin D2 KO mouse (ccnd 2−/−), MGE transplants were also able to normalize hippocampal activity and dopamine cell activity in the VTA. Furthermore, amphetamine-induced locomotor activity was also normalized by MGE transplants in this model. Interestingly, Gilani et al. also found that MGE transplants improve contextual fear conditioning, one type of hippocampal-dependent cognitive function that is disrupted in SZ [135]. The primary interneuron subtypes that have been implicated in schizophrenia are parvalbumin- and somatostatin-positive cells [134] and these experiments found that the majority (56%) of transplanted MGE cells mature into parvalbumin (PV)-positive interneurons, while about 35% become somatostatin (SST)-positive interneurons [135]. To better understand the role of these specific interneuron subtypes, more recent work has used a dual-reporter, mouse, embryonic stem cell line to grow enriched populations of PV- or SST-positive interneurons. These cells were transplanted into the vHipp of the MAM model of schizophrenia. Both PV- and SST-positive interneurons were able to decrease the firing rate and increased sIPSC amplitudes in vHipp pyramidal cells. Furthermore, both cell types reduced hyperactivity in the dopamine system and attenuated deficits in dopamine-dependent cognitive function [136]. Currently prescribed antipsychotic medications have little to no impact on negative or cognitive symptoms [12], therefore it is of particular interest that the PV-positive interneuron transplants were also able to improve social interaction [136], a model of negative symptoms, and extradimensional set-shifting [136], a form of cognitive flexibility that depends on prefrontal cortical function [137] and is disrupted in schizophrenia [138,139]. Together, these results suggest that using stem cells to restore interneuron function in the vHipp may be a useful strategy for treating schizophrenia and, importantly, to target multiple symptom domains.
Autism
Similar to schizophrenia, deficits in GABAergic function have also been implicated in the pathology of autism. Using a rodent model of autism, the potential utility of transplanting interneurons was also tested. PV-positive interneurons were transplanted into the mPFC of the polyinosine:cytosine (Poly I:C) model of autism. Viral infection during pregnancy has been associated with an increased incidence of autism [56,57]. Similarly, injecting pregnant female rats with the viral mimetic, Poly I:C, on gestational day 12 has been shown to produce autism-like deficits in offspring, including reduced communication, social avoidance, and stereotyped or repetitive behaviors [1]. The cell transplants integrated into the circuitry and decreased the pyramidal cell firing rate in the mPFC. Behaviorally, PV-positive transplants also increased social interaction time and perseveration in a test of cognitive flexibility [140], suggesting that this strategy may be useful for alleviating some of the core symptoms of autism.
To date, the majority cell-based therapies for treatment of autism have focused on mesenchymal stem cells (MSCs) [141]. MSCs are multipotent progenitor cells of mesodermal origin that can be isolated from the bone marrow, adipose tissue, liver, tendons, synovial membrane, amniotic fluid, placenta, umbilical cord, and teeth [141]. These cells, which can migrate to the site of tissue injury, have a high expansion rate and demonstrate genetic and phenotypic stability [141]. In the BTBR genetic model of autism, it was shown that intracerebroventricular (i.c.v.) injection of MSCs reduced stereotyped patterns of behavior, improved cognitive flexibility, and increased vocalizations and social behaviors [142]. In a follow-up study, the authors extended their findings to show that the behavioral effects of a single i.c.v. injection of MSCs persisted for at least 6 months after testing [143]. Similar results were found using a developmental model of autism, the VPA model. MSCs injected into the 3rd ventricle reduced repetitive grooming behaviors and increased social preference. Furthermore, the MSCs were also able to decrease anxiety-like behavior on the social interaction test [144]. Excitingly, a few small open-label trials using stem cells in autistic patients have shown promising results. For example, Lv et al. found that intravenous (i.v.) and intrathecal injections (one infusion per week for 4 weeks) of human cord blood mononuclear cells and umbilical cord-derived MSCs produced overall improvements on the Childhood Autism Rating Scale, Clinical Global Impressions (CGI) Scale, and Aberrant Behavior Checklist (ABC) [145]. Similar results were found in another small open-label trial, in which autologous, bone marrow mononuclear cells were transplanted intrathecally. Patients were monitored for over 2 years and overall improvements were observed on the CGI Scale and Indian Scale for Assessment of Autism (ISAA). Specifically, improvements were found on the social relationship and reciprocity, emotional responsiveness, speech, language, and communication, behavior patterns, sensory aspect, and cognitive component subscales [146]. In another small clinical trial, a single infusion of fetal stem cells was administered i.v. to autistic patients and a significant overall improvement was seen on the Autism Treatment Evaluation Checklist and ABC for up to 12 months postinfusion. These improvements were driven primarily by the sociability, cognitive, and health/behavior subscales [147]. Together, these results suggest that stem cell therapy may be beneficial for treating the core symptoms of autism.
Unanswered Questions
While stem cells hold the potential to help us understand and better treat neurodevelopmental disorders such as schizophrenia and autism, multiple questions remain. The first is the validity of 2D or 3D cultures for understanding complex disorders that involve multiple cell types and circuits. Both schizophrenia and autism have been associated with dysfunction of the immune system, and to date, stem cell models do not include the primary immune cells of the brain, microglia. Furthermore, epigenetic mechanisms, or nonpermanent alterations to the 3D structure of DNA that causes changes in gene expression, have been implicated in both schizophrenia [148] and autism [149]. The process of reprogramming somatic cells can erase the epigenetic signature. However, newer process reprogramming, called transdifferentiation, may be able to get around this problem [150,151]. Finally, complex disorders such as schizophrenia and autism involve multiple brain regions and the connections between these regions have been shown to be disrupted. While 3D brain organoids allow regional specification, to date, complex brain circuits have not been formed. Fusion of cortical and subcortical organoids and migration of interneurons into cortical areas suggest that this may be possible [83]. However, especially as brain organoids become more complex, ethical issues will need to be considered. For example, it will be important to know whether the organoids can experience consciousness. Even now, ethical issues such as ownership and consent should be considered [152].
As stem cells move into the clinic as potential therapies, multiple issues will need to be addressed. First and foremost, extensive testing will be required to determine the safety of cell transplants and the duration of their effect. Although MSCs for treatment of autism have already begun to make their way into the clinic, multiple questions still exist related to this highly controversial cell-based therapy [153]. First, MSCs are an extremely heterogeneous population of cells with no specific cell markers [154]. In addition, without targeted delivery, it is impossible to ensure that the cells will reach the target site, especially in the case of autism where there is no overt tissue injury [154]. Furthermore, little is known about the behavior of these cells in vivo, specifically the immunologic properties of the cells. Finally, although MSCs are thought to have their beneficial effects through their ability to suppress immune signaling, promote neurogenesis and plasticity, and release neurotrophic factors, their mechanism has not been completely elucidated. In addition, highly standardized and efficient protocols for growing stem cells will need to be developed. In the case of using interneurons, for example, techniques for generating a highly pure population will be necessary to prevent the possibility of tumorigenesis. Furthermore, the process or growing human interneurons in vitro is currently in line with the time course of human development, suggesting that more efficient protocols need to be developed.
Although questions remain, stem cell technology still has the potential to help us understand complex neurodevelopmental disorders, such as schizophrenia and autism, to develop better methodologies for screening drugs and personalizing medicine, and most excitingly, they may even be used to treat the disease.
Footnotes
Author Disclosure Statement
The authors do not have any conflicts of interest to disclose.
Funding Information
This work was supported by the San Antonio Life Sciences Institute, R01 MH090067 (DL) from the National Institute for Mental Health, TL1TR002647 (JD) from the National Center for Advancing Translational Science, and by K99 MH121355 (JD) from the National Institute for Mental Health.