
Abstract
Many persons with diabetes mellitus have limb ischemia, which is a major clinical problem. A subset of human monocytes that expresses TIE-2 may enhance neovascularization. We performed 179 phlebotomies on 142 patients (or donors), including 61 patients/donors without diabetes or ischemia (controls), 39 diabetic nonischemic patients (controls), and 42 diabetic patients with severe limb ischemia requiring amputation. We compared these groups for the presence of TIE-2-positive proangiogenic monocytes. The proportion of proangiogenic monocytes in the venous blood (on hospital admission) was significantly increased in diabetic patients without ischemia (9.22% ± 1.19%), compared to controls (6.53% ± 0.58%) or ischemic diabetic patients (5.44% ± 0.56%) (P < 0.05). In this pilot evaluation, we succeeded in extracting potential proangiogenic TIE-2 monocytes from the blood of diabetic patients without ischemia, but less in patients with ischemia. The implications for therapeutic neoangiogenesis require further studies.
Introduction
Many persons with diabetes mellitus of long duration develop advanced peripheral arterial disease. For a substantial percent of these patients, especially those with a long-standing atherosclerotic disease or severe peripheral microangiopathy, neither vascular surgery nor angioplasty treatment is feasible. Thus, alternative strategies for improving blood supply to the lower extremities are needed. Because most previous studies of innovative methods have used angiogenic growth factors alone and reported limited efficacy [1], many research groups have turned to investigations of cell-based therapies. Results of local injections of unselected bone marrow- or peripheral blood-derived mononuclear cells have been encouraging, but insufficiently efficacious, likely related to the limited delivery of proangiogenic cells within a complex cell population [2]. Investigations using autologous bone marrow cell transplantation [3] have also met with limited success. Furthermore, on both methods, harvesting of angiogenic stem cells remains a cumbersome procedure.
Unlike embryonic or hematopoietic stem cells, blood monocytes are abundant, readily replenished, and collected by a simple peripheral phlebotomy. A subset of monocytes endowing proangiogenic capacity is identifiable by the expression of TIE-2, a tyrosine-kinase receptor for angiopoietins that enables neovascularization, at least in tumors and a limb ischemia model in mice [4]. Overall, there are three major populations of circulating human monocytes: classical inflammatory (CD14highCD16−); intermediate (CD14highCD16+); and nonclassical (CD14lowCD16+) [5]. Both the intermediate and nonclassical monocyte types express TIE-2, and are considered the most proangiogenic [4,6,7], even though pan-monocytes exist as a continuous spectrum rather than distinct populations, and not all CD16+ monocytes may promote angiogenesis. Patel et al. detected increased levels of peripheral proangiogenic monocytes in three patients with critical limb ischemia [8]. However, designing therapeutic interventions requires understanding the (local) homing of these proangiogenic monocytes in diabetic patients with limb ischemia. We therefore designed a pilot study to compare the proportion of circulating proangiogenic TIE-2 monocytes in different patient populations and to characterize their recruitment capacity in presence of an inflamed vascular endothelium.
Materials and Methods
Sampling and study participants
We enrolled healthy donors (or patients with various orthopedic infections but no evidence of limb ischemia); diabetic patients without clinical evidence of limb ischemia; and diabetic patients with critical limb ischemia requiring amputation. We defined ischemia based on clinical signs and symptoms (eg, claudication, diminished or absent pedal pulses, delayed capillary refill time, and low ankle-brachial index) present on limb examination. Included patients had diabetes mellitus and were not being treated with colony growth factors. Patients with an infection could be enrolled if they were not bacteremic. Infected patients received antibiotic therapy according to international guidelines [1] and we did not change any of the other medications of any participants. We obtained venous blood samples on all enrolled patients from the arm; for the diabetic subjects, these were collected immediately before any orthopedic surgery procedure they were scheduled to undergo. We planned that in a subset of up to 20% of the diabetic patients, we would obtain blood samples multiple times, either during another hospitalization or the recovery period, to see changes over time. Our Ethical Committee (Regional Ethics Committee; République et canton de Genève, Geneva, Switzerland) approved the study (NAC 14-183). The work was performed in accordance with the Declaration of Helsinki and the guidelines for Good Clinical Practice.
Reagents and antibodies
We used Lympholyte-H (Cedarlane Laboratories) or Ficoll-Paque Plus (GE Healthcare) gradients to isolate human peripheral blood mononuclear cells (PBMC) and the Pan-Monocyte isolation kit (Miltenyi Biotech) for monocyte isolation. We applied various commercial antibodies for flow cytometry, the live-imaging of cells, or the immune-fluorescence examinations (Table 1). We did, however, not specifically stain the TIE-2 expression, because they are already included in the (CD14highCD16+) and (CD14lowCD16+) assessments, as previously described by our and other groups [4].
List of Antibodies Used in the Study
Analysis of the proportions of monocyte subpopulations
For targeting monocyte subpopulations, we slightly modified our previously published methods [4]. Briefly, we stored phlebotomy blood samples in refrigerators and processed them within 24 h. The red blood cells of the venous blood samples were first lysed by red blood cell lysis buffer (8.3 g NH4Cl, 1 g NaHCO3, and 1 mL ethylenediaminetetraacetic acid (EDTA) [100 mM]). Afterward, we analyzed the PBMC with a BD Accuri C6 cytometer (BD Biosciences). We incubated the various antibody combinations with FcR blocking reagents in staining buffer, which contained 0.5% bovine serum albumin (BSA) and 2 mM EDTA in phosphate-buffered saline (PBS). After staining, we washed the PBMC in a7-AAD buffer to exclude nonviable or dead cells. We identified proangiogenic monocytes by flow cytometry. Cells expressing the lineage markers CD15, CD2, CD3, CD56, NKp46, CD1c, and CD141 removed the granulocytes, lymphocytes, and immature dendritic cells. We identified all the monocytes collected in the human leukocyte antigen—DR isotype (HLA-DR) and CD300e-positive windows. All monocytes were negative for granulocyte, lymphocyte, natural killer (NK), and immature dendritic cell markers (CD15−, CD2−, CD3−, CD56−, NKp46−, CD19−, CD1c−, and CD141−) and positive for the monocyte markers (HLA-DR+ and CD300e+). Within the pan-monocyte population, we distinguished three subpopulations according to their expression of CD14 and CD16 using the Galios cytometer (Beckman Coulter). We have previously shown that both CD14highCD16+ and CD14lowCD16+ monocytes express TIE-2 at different levels, but present similar proangiogenic activity [4]. Both markers are similar in the identification of TIE-2 blood monocytes [9,10]. However, CD16+ monocytes contain the primary population of TIE-2 monocytes, which are capable of promoting vascularization through the secretion of various factors [9 –11]. Other groups have also reported that CD16+ monocytes are TIE-2-positive proangiogenic monocytes in human [6,9 –12]. Thus, in this study, we focused on CD16+ monocytes.
Monocyte recruitment under flow
We assessed the recruitment potential of proangiogenic monocytes according to previous reports [4,13 –16]. As proangiogenic monocytes must find their way to inflamed tissues for wound healing, we tested the recruitment capacities of these proangiogenic monocytes by measuring their adhesion to experimental vascular endothelium [primary human umbilical vein endothelial cells (HUVEC)]. Briefly, we coated six chambers of μ-Slide VI 0.1 ibi Treat (IBIDI) with 0.2% gelatin (Sigma-Aldrich), and 1 mg/mL Collagen G (Biochrom AG) in PBS, and then cultured HUVEC (3 × 104 cells/chamber) in M199 medium (20% Fetal Calf Serum, 1% Endothelial Cell Growth Supplement (Thermo-Fisher Scientific), 1% penicillin, 1% streptomycin, 0.1 mg/mL heparin sodium, 0.1 μM hydrocortisone, and 10 μg/mL L-ascorbic acid) for 7 days, with medium change every other day. We then stimulated the resulting monolayers with 1 μg/mL vascular endothelial growth factor (VEGF; Peprotech) and 1,000 U/mL tumor necrosis factor α (TNF-α; Peprotech) for 6 h to induce an angiogenic inflammation. For this, we generated flow over the HUVEC monolayer by perfusing washing buffer (containing 0.2% BSA in M199 prewarmed at 37°C) over the HUVEC monolayers using a calibrated pump (74,900; Cole Parmer). The flow rate was representative of shear rates in postcapillary venules (0.05 Pa). An image of a fixed field was generated every 15 s during 25 min (Axiovert 200 microscope; Carl Zeiss). After 1 min of recording with Openlab software, we used a switch system to deliver prestained pan-monocytes (3 × 106 cells/mL) under continuous uninterrupted flow for 5 min.
Transendothelial locomotion of proangiogenic blood monocytes
We furthermore evaluated the transendothelial locomotion of the blood proangiogenic monocytes. First, we isolated the CD16+ monocytes by using anti-CD16 (clone: 3G8) coupled with anti-mouse IgG Microbeads (Miltenyi Biotech). Proangiogenic monocytes were retained within an LS column (Miltenyi Biotech) on the magnet (Miltenyi Biotech). We then used the isolated cells in the transmigration assay by fixing them with 4% paraformaldehyde for 10 min and permeabilized them with 0.2% Triton-X-100 for another 10 min. The Slan+ cells were stained with anti-Slan-FITC for 30 min. Finally, we determined the proportions of adherent and transmigrated Slan+ monocytes for all three groups of patients.
Statistical analyses
Our statistical analyses were basic. Data were presented as mean ± standard deviation or mean ± standard error of mean, or dot-plot with median values. The analyses were performed using one-way analysis of variance (ANOVA) with Holm-Sidak adjustment (Graphpad).
Results
Patients
From March 2015 to December 2017, we collected blood (6 mL) from the arm veins of 142 different persons. As planned, we sampled blood from 29 participants multiple times to determine the levels of proangiogenic monocytes over time; thus, we performed a total of 179 phlebotomies. Among the enrolled patients, 61 were healthy donors (or nonischemic patients with various orthopedic infections); 39 were diabetic patients without clinical evidence of limb ischemia; and 42 were diabetic patients with critical limb ischemia requiring amputation. The corresponding numbers of phlebotomies were 80, 42, and 57, respectively. Overall, 113 patients were sampled once, 22 were sampled twice, 6 three times, and 1 four times (Table 2). The patients' median age was 64 years (range, 26–84 years), and 53 were female.
Key Demographic Variables of the Study Population (Hospitalized Patients and Controls)
Quantification of proangiogenic TIE-2 monocytes
Nonclassical proangiogenic, CD16+ monocytes were distinguished from the CD16−/CD14+ nonangiogenic, inflammatory monocytes by flow cytometry (Fig. 1A). We renounced on supplementary staining methods for identification or quantification for practical reasons, because we judged the flow cytometry method as sufficient for our study goals. The median blood monocyte quantification showed that blood from nonischemic diabetic patients contained 80% or more proangiogenic monocytes compared to both other study groups (Fig. 1B). The proportion of proangiogenic monocytes was significantly increased in diabetic patients without ischemia (9.22% ± 1.19%) compared to controls (6.53% ± 0.58%) or ischemic diabetic patients (5.44% ± 0.56%) (one-way ANOVA-test, P < 0.05). In cases of multiple sampling, this quantification of samples from the same patient did not significantly differ over time in the samples.
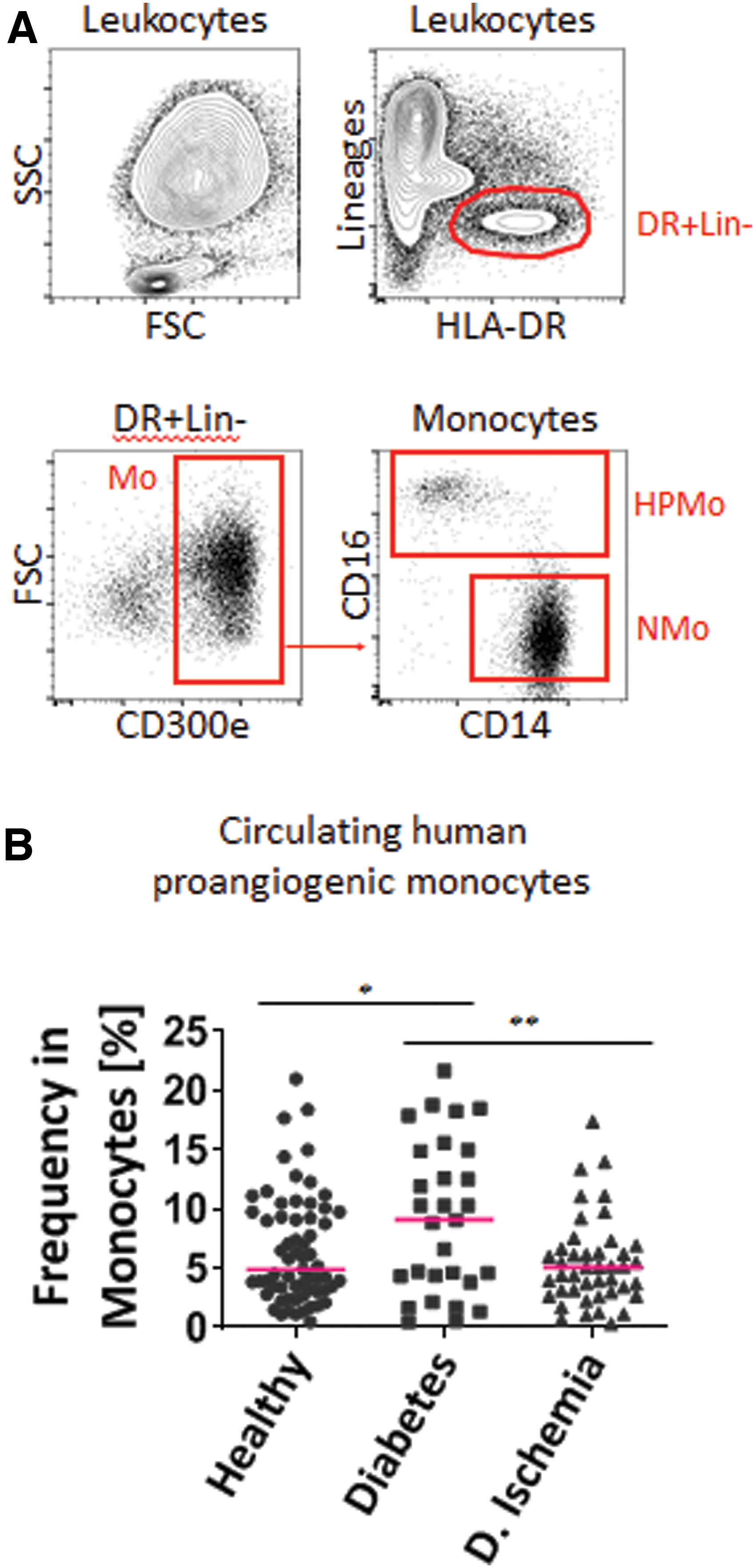
Analysis of HPMo in blood of healthy donors and patients.
Recruitment capacity of TIE-2 monocytes and their transendothelial migration
For the recruitment of proangiogenic monocytes, we induced a proangiogenic inflammation to HUVEC by stimulating them with the combination of VEGF and TNF-α. Between the three study groups, there was no difference regarding the adhesion potential (Fig. 2A). However, the number of adherent cells in diabetic patients with ischemia was twice as high as those in both of the other comparator groups. In testing the transmigration of adherent monocytes, we found that their numbers in ischemic diabetic patients were again higher compared to those in the other two groups (Figs. 2B, C). To further explore this difference, we assessed the expression of molecules involved in the adhesion among the study groups, as described below.
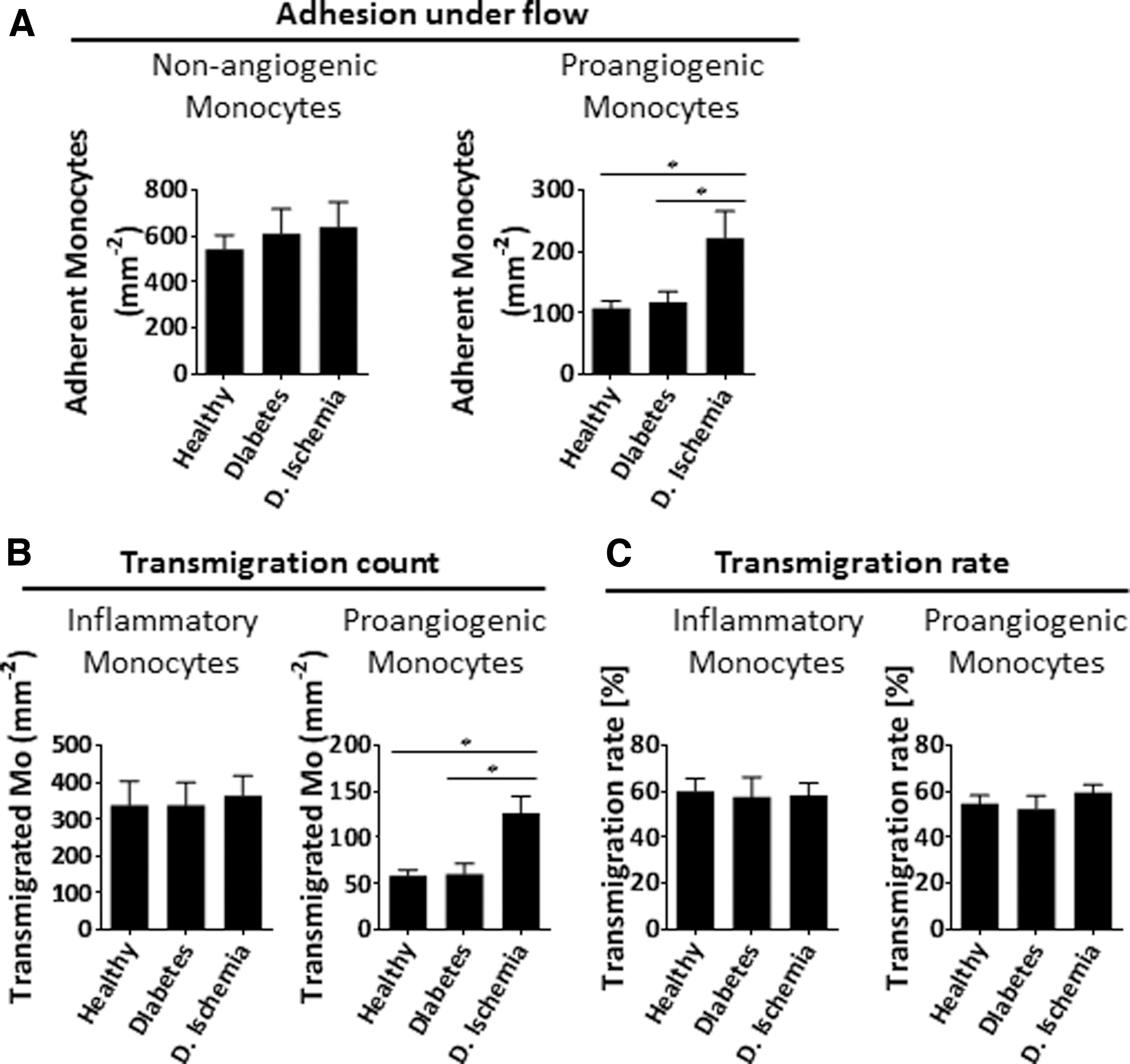
Recruitment capacity of HPMo derived from healthy donors and patients.
Expression of adhesion molecules and chemokine receptors by proangiogenic monocytes
As we wished to determine the expression of adhesion molecules on the protein level, we performed flow cytometry using the windows highlighted in Figs. 1A and 3A. Among possible relevant molecules, we analyzed L-selectin, the integrins LFA-1 and Mac-1, the fractalkine receptor CX3CR1 (a key chemokine receptor on angiogenic monocytes), and the selectin receptor PSGL-1. None of these showed significant differences among the different study populations. We then searched for the expression of the PSGL-1 protein that could be post-translationally modified by 6-Sulfo LacNAc glycosylation (Slan). This modification inactivates PSGL-1 as a ligand for vascular P- and E-selectin [17,18]. We found that the Slan-PSGL-1 expressions were drastically reduced in diabetic patients with ischemia compared to both of the other groups (Fig. 3B, C). The Slan-PSGL-1 cells did not transmigrate and were encountered in the blood only, but did not adhere to endothelium. Finally, we quantified the proangiogenic monocytes in the blood, their adhesion to inflamed HUVEC, and their transendothelial migration (Fig. 4A–C). Figure 5 illustrates the impact of PSGL-1 and its Slan-modified version on monocyte capture on vascular endothelial cells.

Analysis of the expression of adhesion molecules and chemokine receptor by HPMo derived from healthy donors and patients.
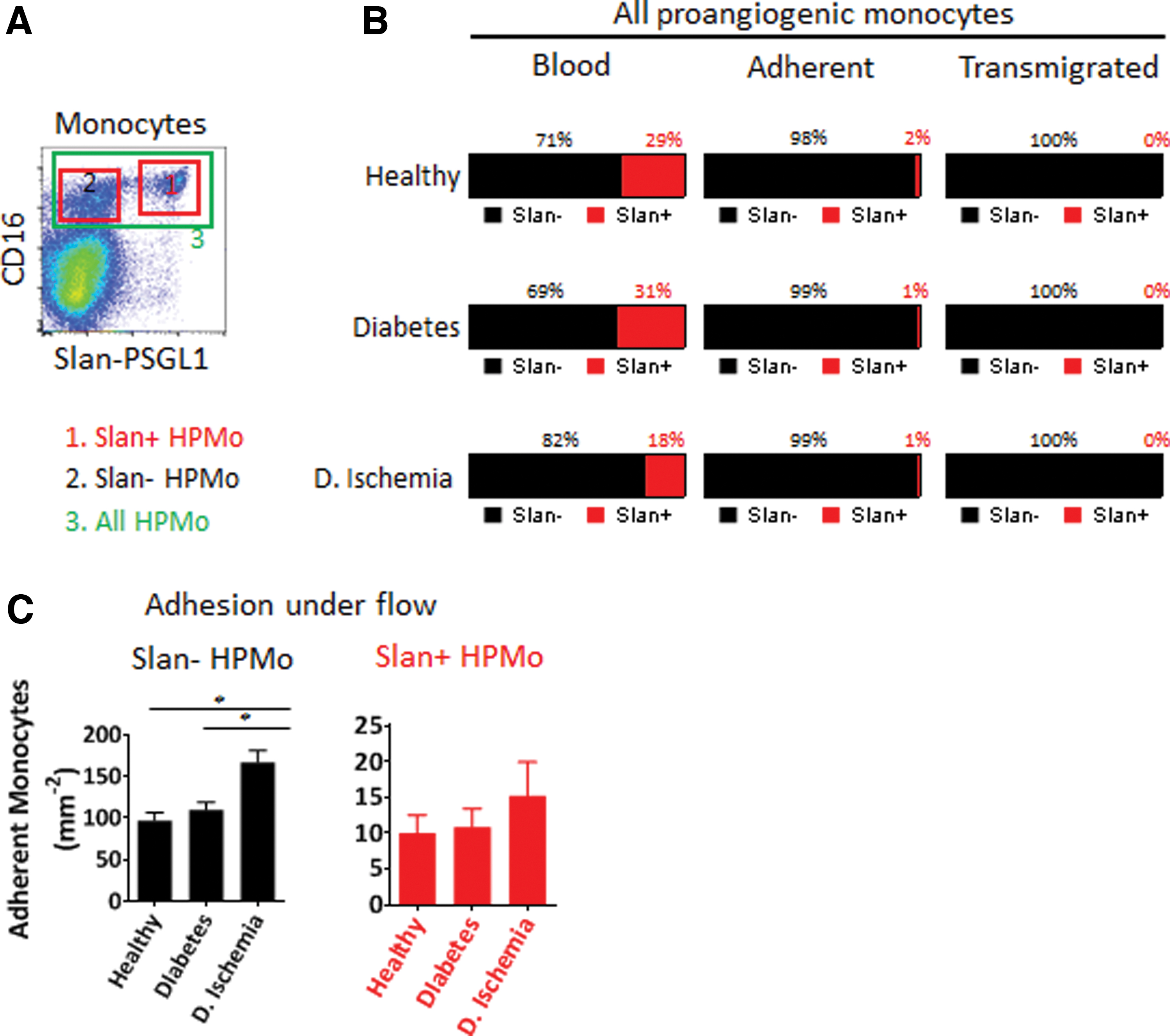
Effect of PSGL1 inactivation by 6-Sulfo LacNAc glycosylation of human proangiogenic monocyte on adhesion and transmigration.
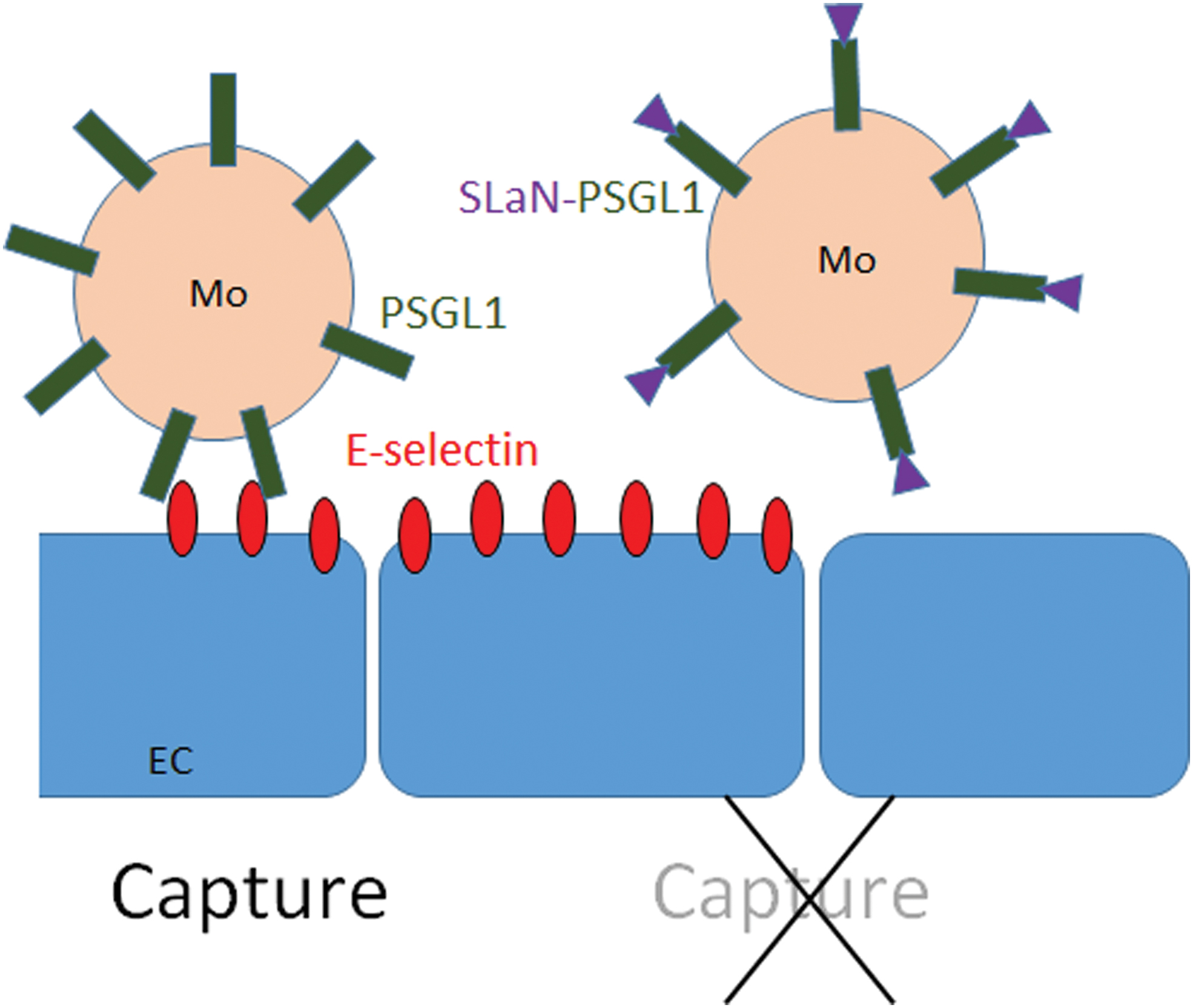
Scheme illustrating the effect of PSGL-1 and its Slan-modified version on monocyte capture by E-Selectin on vascular EC. EC, endothelial cells. Color images are available online.
Discussion
In this pilot evaluation, we found that diabetic patients with severe lower limb ischemia had quantitative and functional changes of circulating blood proangiogenic monocytes that affect their local recruitment and their local proangiogenic role. Overall, we successfully derived TIE-2 monocytes from the blood of healthy donors and nonischemic diabetic patients and expanded them in vitro in animal-free real-life conditions. However, the levels of these monocytes in blood were reduced in the most affected group, that is, diabetic patients with limb ischemia, even when their individual transmigration was higher than for the two comparator groups. Previous in vitro or in vivo studies suggested a role of proangiogenic (TIE-2) monocytes in wound healing [8]. We found that in diabetic patients without ischemia, the level of circulating proangiogenic monocytes was increased, suggesting that ischemia affects the proangiogenic monocytes more than the diabetes itself [19]. While the increase of circulating proangiogenic monocytes in patients with diabetes without ischemia may be a systemic response in favor of the repair of dysfunctional vessels, this assumed advantage may be lost during ischemia.
We have previously shown that both CD14highCD16+ and CD14lowCD16+ monocytes express TIE-2 at different levels, but present similar proangiogenic activity [4]. In this study, we focused on CD16 expression to identify proangiogenic monocytes. Although TIE-2 expression is very relevant, in humans, the CD14highCD16+ and CD14lowCD16+ monocytes are similarly proangiogenic, despite different levels of TIE-2 expression [4]. This suggested that in humans, the CD16 expression is more relevant in blood circulating monocytes than TIE-2 staining. Accordingly, we quantified CD16+ monocytes in this study rather than TIE-2.
Our study also provided new insights on the adhesion of proangiogenic monocytes to the inflamed endothelium and the transendothelial migration in diabetic persons. The reduction of the blood level of TIE-2 monocytes paralleled a twofold adhesion to experimental inflamed human endothelium. The transmigration of proangiogenic monocytes requires angiogenic inflammation by the simultaneous actions of angiogenic factors and inflammatory cytokines, such as TNF-α [4,20]. In addition, the adhesion molecule PSGL-1, critical during the capture process of monocytes, can be subject to the 6-sulfo-N-acetyl-lactosamine glycosylation in proangiogenic monocytes [17 –22]. This modification usually alters the adhesion capacity of proangiogenic monocytes to vascular endothelium. In our study, we were surprised to find that both the frequency of Slan+ monocytes and the glycosylation level of PSGL-1 strongly decreased in diabetic patients with critical ischemia. Possibly, the monocytes of ischemic diabetes patients are more adhesive to the vascular endothelium and that is the reason we see less of them circulating. In diabetic rats, the injection of autologous monocytes, together with TNF-α and/or interferon-gamma, significantly improved wound healing through angiogenesis [23]. Based on currently available scientific knowledge and in contrast to studies in rodents, future therapies in humans based on the use of autologous, proangiogenic monocytes is a complex field. We need further understanding of the proangiogenic monocyte differentiation from progenitor cells in bone marrow, their expansion in vitro, and the prevention of PSGL-1 Slan-glycosylation to improve their capacity to selectively migrate to ischemic regions. Alternatively, proangiogenic monocytes may be isolated from immune-compatible healthy donors and be used for therapy. For example, monocytes have been derived from human embryonic stem cells and shown to increase angiogenesis in xenotransplanted human tumors [24]. The long-term risks of these proangiogenic monocyte enhancements in humans have not been determined. In the future, it could be possible to use these cells to improve wound healing in diabetic patients with limb ischemia, if we can find ways of avoiding the potential long-term neoplastic effects of these cell lines. In healthy donors, we estimate infusing about 250–1,000 proangiogenic monocytes/mL would be necessary for neoangiogenesis in ischemic areas [24], but the available literature lacks benchmark numbers. Moreover, we currently ignore the minimal, or optimal, quantity for monocyte therapy in diabetic patients. We also do not know if we should inject proangiogenic monocytes alone or concomitantly with colony-stimulating factors, as many basic researchers advocate [25]. Likewise, we ignore so far how these monocytes behave inside specific patient populations such as diabetic patients with and without ischemia and controls, or if the cells remain basically the same. Our literature search did not find any particular hints that the blood monocytes would be altered per se in their structure, only because of the underlying chronic comorbidity, which, however, would be logical given the differences in our group comparisons. Finally, the future will show if the approach of autologous proangiogenic monocyte administration through the venous blood phlebotomy is a better way to go than, for example, the classical angiogenesis factor harvesting from pluripotent (adipose) stem cells. This approach is another field of increasing research, more advanced than monocyte harvesting, but also much costlier and more cumbersome [26].
In conclusion, we clearly succeeded in extracting proangiogenic TIE-2 monocytes from the venous blood of diabetic patients without clinical ischemia, but were less successful with the target population of diabetic patients with critical ischemia. Possibly, the monocytes of severely ischemic diabetes patients are more adhesive to the vascular endothelium and that is the reason we see less of them circulating. The implication of these findings for therapeutic neoangiogenesis will require further studies. There is equally still room for further preliminary studies before the first therapeutic trials.
Footnotes
Acknowledgment
We are indebted to the Nursing Teams of the Orthopedic Service of Geneva University Hospitals for their invaluable help.
Data Availability
Anonymized data can be obtained on reasonable request upon the corresponding author.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.