
Abstract
Photoreceptor transplantation can rescue the retinal function of late-stage rd1 mice. Many studies have used synaptic markers to suggest that there are synaptic connections after transplantation, but how donor and host cells are connected remains unknown. Many molecules are needed for triad ribbon synapse formation in wild-type mice. Among them, pikachurin is an important extracellular matrix protein that bridges the pre- and postsynaptic components. To investigate the mechanism of the synaptic connection between donor photoreceptor and host retina, we studied the expression of pikachurin in late-stage rd1 mice before and after transplantation. The results showed that the full-length form of pikachurin could still be detected in the degenerated retina. After photoreceptors were transplanted to the subretinal space of rd1 or wild-type mice, pikachurin was detected in the cytoplasm of most donor photoreceptor cells. Pikachurin puncta may represent the cleaved form of the protein and may indicate synapse generation, but it was barely observed in the donor mass of wild-type mice (3.83 ± 3.17 puncta per 100 donor cells). In contrast, pikachurin puncta could be found in the graft of the rd1 mouse retina, but the number was low (21.35 ± 9.48 puncta per 100 donor cells). In addition, 54.12 ± 8.45% of bassoon puncta were paired with pikachurin puncta and 45.5 ± 6.33% were not, indicating that there were fewer pikachurin puncta than bassoon. These results suggest that pikachurin is involved in only a portion of the synaptic connection between the donor photoreceptor and host retina.
Introduction
Photoreceptor transplantation is a promising way to rescue retinal degeneration [1]. The key of this therapeutic method is that donor cells integrate into the host retina. However, a recent discovery about material exchange indicated that most donor cells cannot connect the retinal neural circuit in wild-type mice [2 –5]. rd1 mice, which harbor a mutation in PDE beta gene, are a naturally occurring model for retinal dystrophies [6]. Compared with the intact retinal structure, the late-stage rd1 mice almost lost the entire outer nuclear layer (ONL), which makes the physical contact between the donor cell and host inner nuclear cell easier [7]. In rd1 mice, several sources of grafts have successfully connected to the downstream neuron and restored some visual function [7 –9].
In photoreceptor transplantation studies, the typical method used to indicate new synapse generation between donor and host cells is to perform immunodetection around donor cells for synaptic proteins, such as synaptophysin [7,9], ribeye [9,10], and bassoon [11]. Evidence regarding signal transduction has also been found by electrophysiological examination [8,12,13]. However, it is still unclear how the graft and host tissue connect. In the brain, several studies have shown that transplanted neurons can form appropriate afferent and efferent projections [14 –16]. In contrast, the grafted photoreceptor seems unable to develop axon-like structures that form neural synaptic connections [11,17,18].
To form neural connections, a network of transmembrane receptors, secreted ligands, and synaptic cell adhesion molecules are needed to coordinate pre- and postsynaptic assembly [19 –25]. Determining which synaptic connection proteins are expressed may uncover the characteristics of these neural connections between donor and host cells.
Pikachurin is exclusively secreted by photoreceptor, and it is an extracellular matrix protein that is important for the proper triad ribbon synaptic structure formation [19,23,26]. In the vertebrate retina, two forms of pikachurin have been detected: the full-length form (110 kDa) and the post-translational cleaved N-terminal fragment (60 kDa) [27]. The latter is involved in synaptic connection, bridging presynaptic and postsynaptic components [28]. Furthermore, pikachurin showed a high degree of structural divergence between different species according to a computational modeling analysis, which may lead to malfunction of synaptic connections when human stem cell-derived photoreceptors are transplanted to mice retina [29]. Thus, it is critical to assess the involvement of pikachurin in new synaptic connections after photoreceptor transplantation.
In this study, we investigated the expression of pikachurin before and after conspecific photoreceptor transplantation in late-stage rd1 mice. Pikachurin puncta was found in the donor cell mass, but only a part of the synaptic connection between graft and host could be formed by pikachurin.
Methods
Animals
The mice used in this study were as follows: C57Bl/6J mice obtained from the Experimental Animal Center of Kunming Medical University; C3HPde6brd1(rd1) mice purchased from Vitalstar Biotechnology Co., Ltd. (Beijing, China); Rho-icre mice [B6;SJL-Pde6b+ Tg(Rho-icre)1Ck/Boc] (Cyagen, China) were crossed to tdTomatofloxed mice (Cyagen, China) to generate rho-tdTomato mice. Six- to eight-week-old rd1 mice were used as late-stage rd1 host mice. All animal studies were approved by the Animal Ethics Committee at the West China Hospital and were performed with adherence to the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research. The animals were raised at room temperature with a 12-h light/12-h dark cycle and standard mice food and clean drinking water.
Immunofluorescence
Eyeballs were fixed with 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) (pH 7.4) at 4°C for 60 min, and then they were dehydrated in 30% sucrose overnight, and embedded in optimal cutting temperature compound (OCT). Cryostat sections were cut at 10 μm and collected on glass slides and stored at −20°C.
For immunostaining, the retinal sections were incubated in blocking solution (5% normal donkey serum, 1% bovine serum albumin, and 0.03% Triton X-100 in 1 × PBS) for 1 h and then were incubated with primary antibodies at 4°C overnight. The following primary antibodies used in this study: Pikachurin (#14578-1-AP; Proteintech, #011-22631; Wako), Ap2a (; SC-8975; Santa Cruz Biotechnology), PKC-a (P5704; Millipore Sigma), rhodopsin (SC-57433; Santa Cruz Biotechnology), Brn3 (SC-6062; Santa Cruz), glutamine synthetase (MAB302; Millipore), and Bassoon (CST; #6897). Subsequently, slides were incubated with a secondary antibody (Invitrogen) for 1 h at room temperature in the dark. Finally, slides were counterstained with 4′6-diamidino-2-phenylindole (Sigma Aldrich Corp.) and were mounted with Mowiol mounting medium. Stained sections were analyzed using a Zeiss Axio Imager Z2 fluorescence microscope or Nikon A1RMP confocal microscope.
Western blot
Total protein was extracted from retinas and from sorted retina cells using radio-immunoprecipitation assay (RIPA) lysis buffer containing protease inhibitor cocktail (Roche), or using phosphatase inhibition (IP) buffer (300 mM NaCl, 50mM Tris-HCl pH 7.4, 1% Triton X-100) with protease inhibitor cocktail and phosphatase inhibitors (Beyotime Institute of Biotechnology, China) [27]. The protein concentration was determined with a bicinchoninic acid assay kit (Beyotime). Protein (30 μg) was loaded into and separated by a 12% sodium dodecyl sulfate–polyacrylamide gel.
Next, proteins were transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA). After blocking with 5% nonfat milk, the membranes were incubated with a primary antibody. After washing, the membrane was incubated with secondary antibody for 1 h. The images were captured on the Odyssey CLx Infrared Imaging System (LI-COR Biosciences, Lincoln, NE). Western blotting bands were quantified using Odyssey CLx v2.1 software and normalized to beta-actin as loading control.
Real-time PCR
Total RNA was extracted from retina using TRIzol Reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. After quantification with a Nanodrop Spectrophotometer (NanoDrop Technologies, Wilmington, DE), cDNA was synthesized from 1 μg of total RNA with PrimeScript RT reagent kit (Takara Biotechnology, Dalian, China).
To quantify the cDNA, real-time PCR was performed on qTOWER 2.2 system (Analytik Jena, Germany). The PCR amplification was conducted in a volume of 20 μL using EvaGreen Supermix (Bio-Rad Laboratories, Inc., Hercules, CA). The primers used in our study to analyze the expression of pikachurin were AAGGTGTGACTGATCCTGCA and CCCTAGGTCCCACACAACAT. To determine mRNA expression, all samples were tested in duplicate and the average Ct values were used for quantification. The mRNA expression was normalized to the endogenous reference gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
N-methyl-N-nitrosourea treatment
1% N-methyl-N-nitrosourea (NMU; Sigma-Aldrich, Buchs, Switzerland) in saline containing 0.05% acetic acid was injected intraperitoneally. The final concentration was 60 mg/kg body weight. Control animals received a similar volume of balanced salt solution. No clinical signs of discomfort or weight loss were observed. Seven days after NMU administration, the mice were killed, and the expression of pikachurin was examined.
Donor cell preparation and fluorescent-activated cell sorting
The mice were killed according to the approved protocol, and the eyes were removed. The retinas were dissected in cold PBS. Digestion was performed by using the Papain Dissociation System (Worthington Biochemical Corp., Lakewood, NJ). In brief, the retinas were placed in a papain solution for 15 min at 37°C. After this incubation, the retinas were triturated with a 1,000 μL plastic pipette tip to dissociate them into single cells. The papain reaction was stopped with the addition of ovomucoid albumin inhibitor, and cells were collected by centrifuging at 200g for 5 min.
Cell sorting was performed with a FACSAria SORP (Becton, Dickinson and Company) fitted with a 561 nm yellow-green laser to excite tdTomato. Tomato-positive cells were collected in the 586/15 nm channel. Fluorescence-negative cells were collected as required. All the donor cells [photoreceptor precursor cells (PPCs)] were acquired from P4 to P6 mice.
Transplantation
The recipient mice were anesthetized with a mixture of ketamine and xylazine. A tunnel was made in the scleral with a 30-gauge needle, and donor cell suspension was injected through this tunnel by a blunt 32-gauge needle. One microliter cell suspension (1 × 105 cell/μL) was injected into the subretinal space slowly. The pipette was made to remain in place for 2 min and withdrawn slowly. Antibiotic ointment was applied to the eye after surgery. Two weeks after transplantation, the mice were killed to perform the following examination.
Statistical analysis
All experiments were carried out with n ≥ 3. Puncta were manually counted in three-dimensional projections using the original data files. Two different channels were examined independently and together to score both paired and single puncta. Statistical analysis was performed using the GraphPad Prism software (GraphPad Prism Software, Inc., San Diego, CA). The results were analyzed by Student's t-tests. The threshold for significance was set at P < 0.05.
Results
Detection of pikachurin in rd1 mice
In wild-type mice, pikachurin was predominantly expressed in the outer plexiform layer (OPL), as previously reported (Fig. 1A). In rd1 mice, most of pikachurin was detected in the ganglion cell layer (GCL), and the apical side of ONL and inner nuclear layer (INL) at postnatal day 14 (Fig. 1B). After photoreceptor degeneration, the pikachurin was found in the INL and GCL at p42 (Fig. 1C). This observation in rd1 mice was unexpected, as pikachurin was previously found to be exclusively expressed by photoreceptors [26]. Next, we immunostained pikachurin in another retinal degeneration model, which is induced by treatment with NMU [30]. Pikachurin was also found in the INL and GCL at 7 days after NMU administration (Fig. 1D).
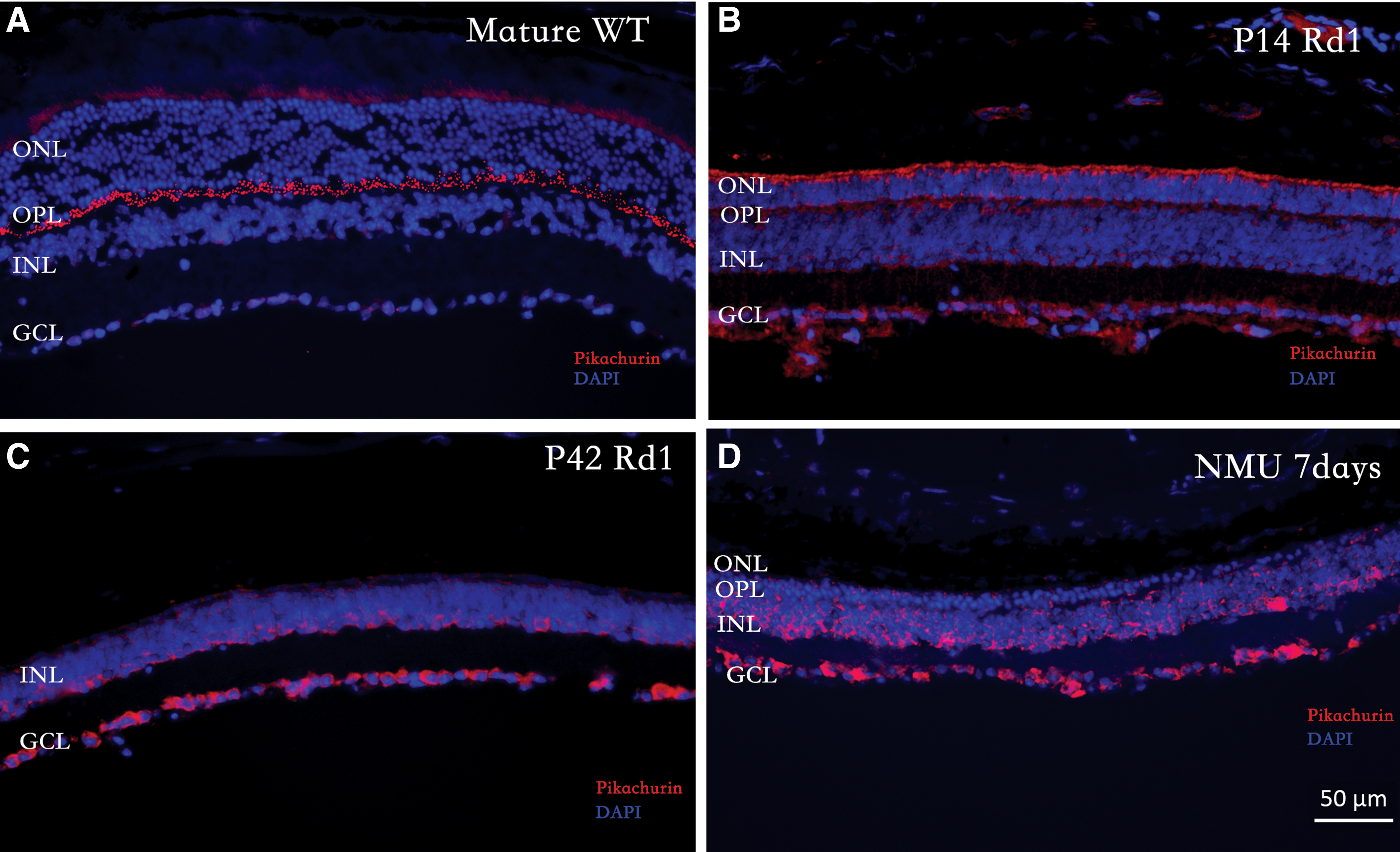
Expression of pikachurin.
To further confirm this observation, we performed western blotting to detect pikachurin in fluorescent-activated cell sorting (FACS) photoreceptor cell/nonphotoreceptor cell, wild-type retina tissue, and rd1 retinal tissue. In Fig. 2A, we find a 60 kDa band (cleaved form) in the photoreceptor cell fraction (PF), but not in the non-PF. This form was also found in the wild-type retina at different time points. However, in rd1 retinal tissue, we detected the full-length form of pikachurin (110 kDa) instead of the cleaved form (60 kDa). The expression of the 110 kDa fragment was significantly increased from age p14 to p42. Real-time PCR results showed that the mRNA level was significantly decreased in rd1 mice compared with that of wild-type mice.
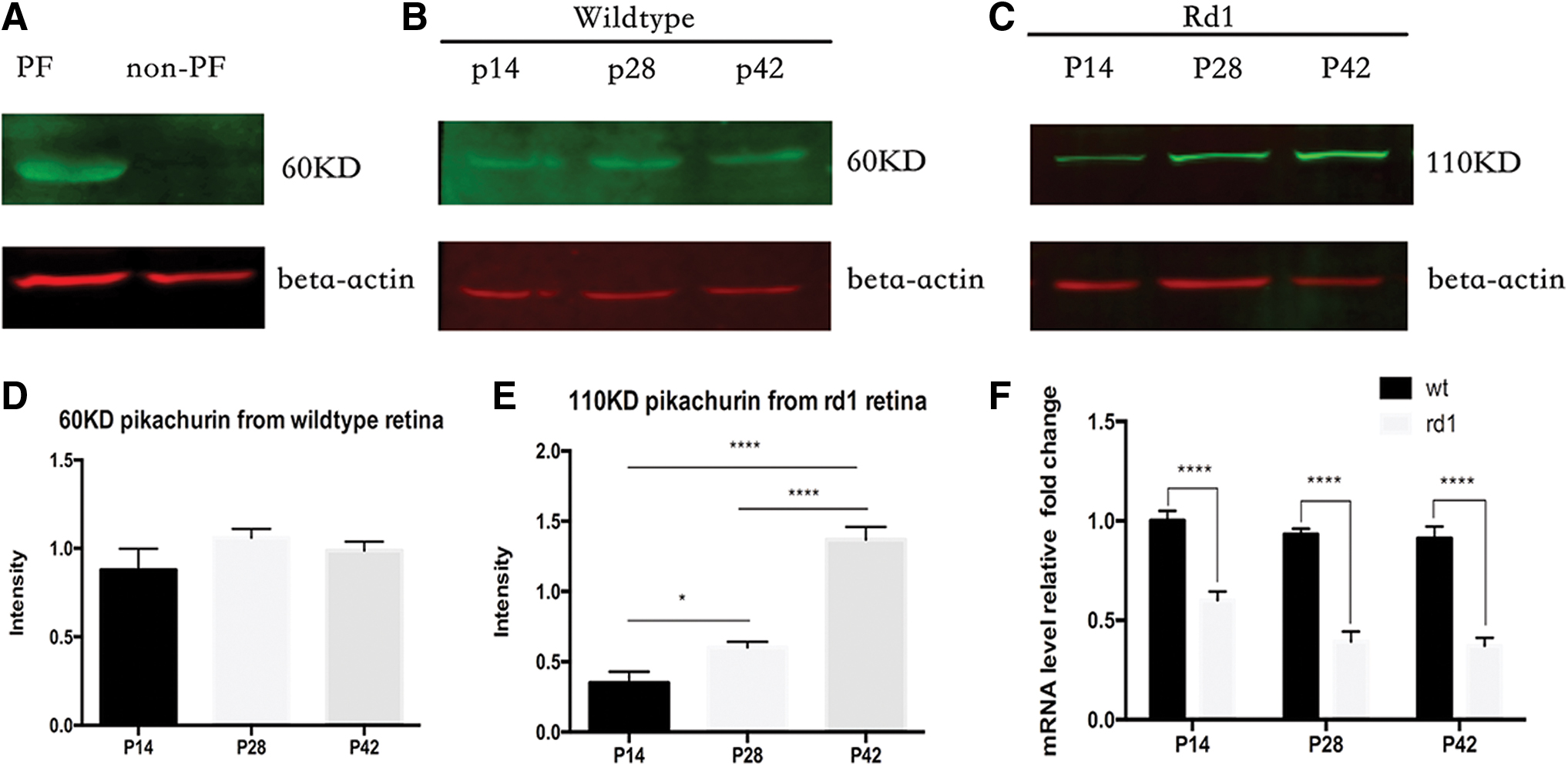
Detection of pikachurin by western blot and real-time PCR.
Distribution of pikachurin in rd1 mice
We used confocal images to analyze the distribution of pikachurin in rd1 mice. At p14 in rd1 mice, rhodopsin-labeled photoreceptor cells and outer segments were immunoreactive for pikachurin (Fig. 3A). At p42 in rd1 mice, many retinal cells were positive for pikachurin, including a large population of bipolar cells (PKCa), amacrine cells (AP2a), and ganglion cells (Brn3a) (Fig. 3B, C, and E). Only Müller cells were almost negative for pikachurin staining (Fig. 3D).
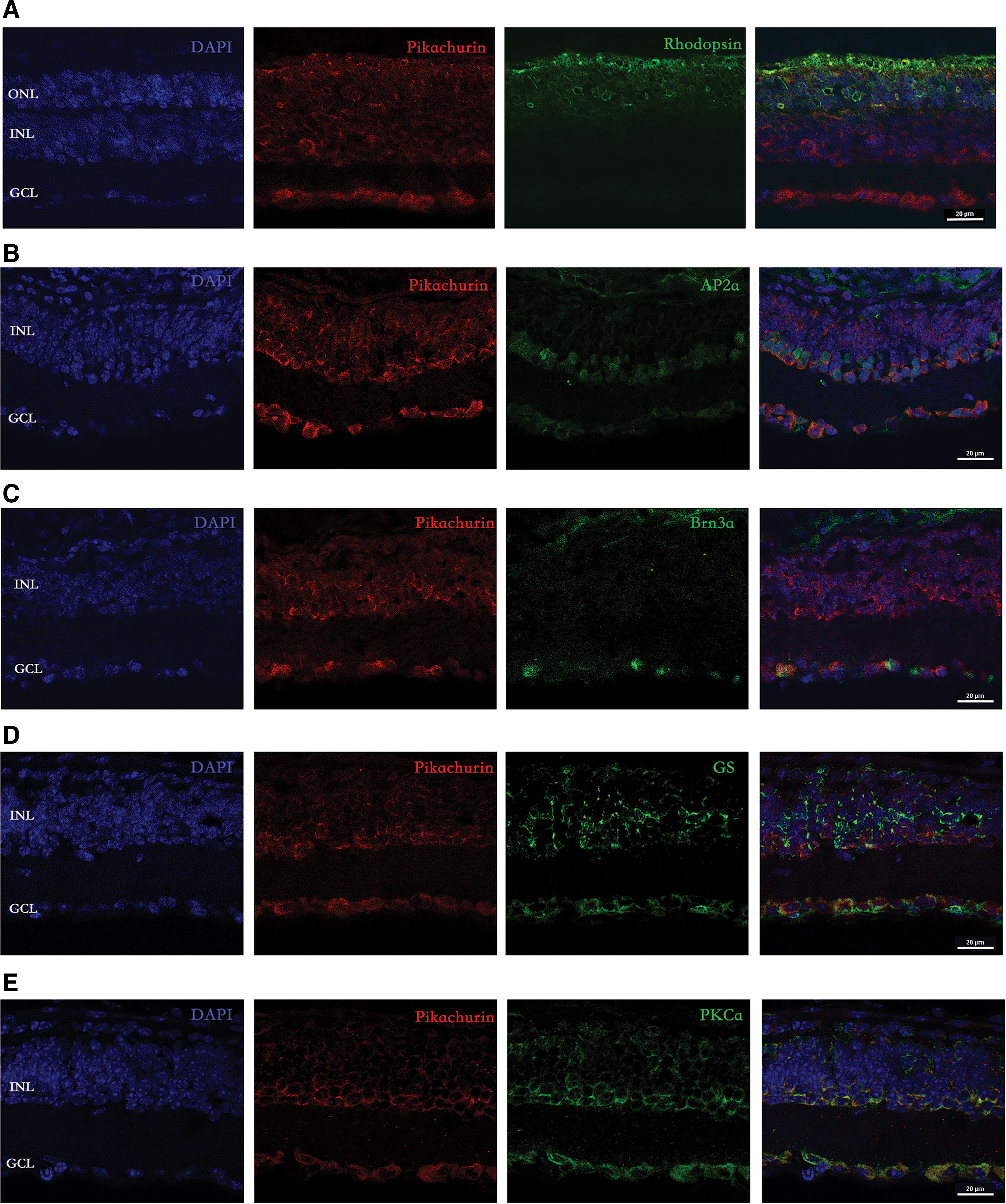
Confocal micrographs showing the pattern of pikachurin labeling in rd1 mice.
Immunodetection of pikachurin in late-stage rd1 mice after photoreceptor transplantation
To determine whether pikachurin was involved in new synapse formation after photoreceptor cell transplantation, P4–P6 PPCs were injected into the subretinal space of late-stage rd1 mice. After transplantation, surviving donor cells could be traced by fluorescence protein, and rhodopsin was well expressed in these photoreceptor cells (Supplementary Fig. S1). Pikachurin could be detected in both the host retinal cells and the donor cells. In the donor photoreceptor mass, we also noticed punctate staining of pikachurin, which may be the post-translational cleaved form of pikachurin involved in synaptic connection (Fig. 4). To verify this possibility, we costained bipolar cells and found pikachurin puncta between donor cell and host bipolar cell (Supplementary Fig. S2).
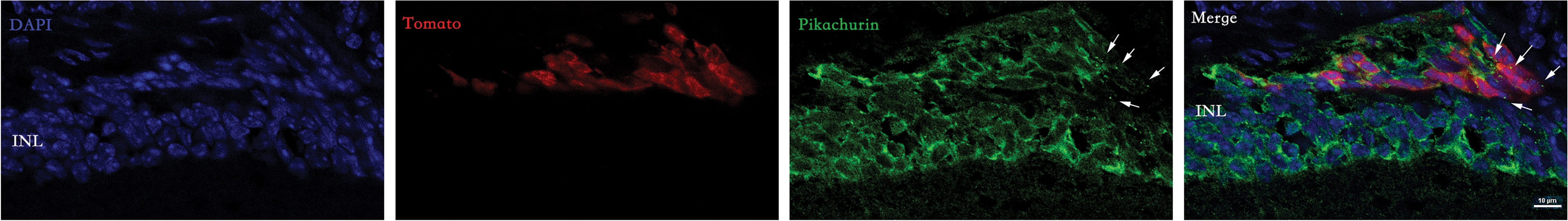
Detection of pikachurin in rd1 mice after photoreceptor transplantation. Donor cells formed a mass under the retinal space, and pikachurin was found in the cytoplasm of donor cell. Some punctate staining of pikachurin was also observed (arrows). Color images are available online.
Next, we performed photoreceptor transplantation in wild-type retina. Although donor photoreceptor was close to bipolar cell in late-stage rd1 mice, graft was separated from INL by endogenous photoreceptor in wild-type retina. As it shown in Fig. 5, we found the “isolated” donor mass above the slightly detached retina, and no synaptic connection could form between donor cell and host downstream neuron (connections that support material exchange may exist between donor and host photoreceptor, but they are not synaptic connections). In this situation, pikachurin puncta were barely observed. If the number of puncta in the donor mass was counted, we found that it was significantly higher in rd1 mice than it was in wild-type mice (21.35 ± 9.48 vs. 3.83 ± 3.17 per 100 tomato-positive cells, P < 0.05).
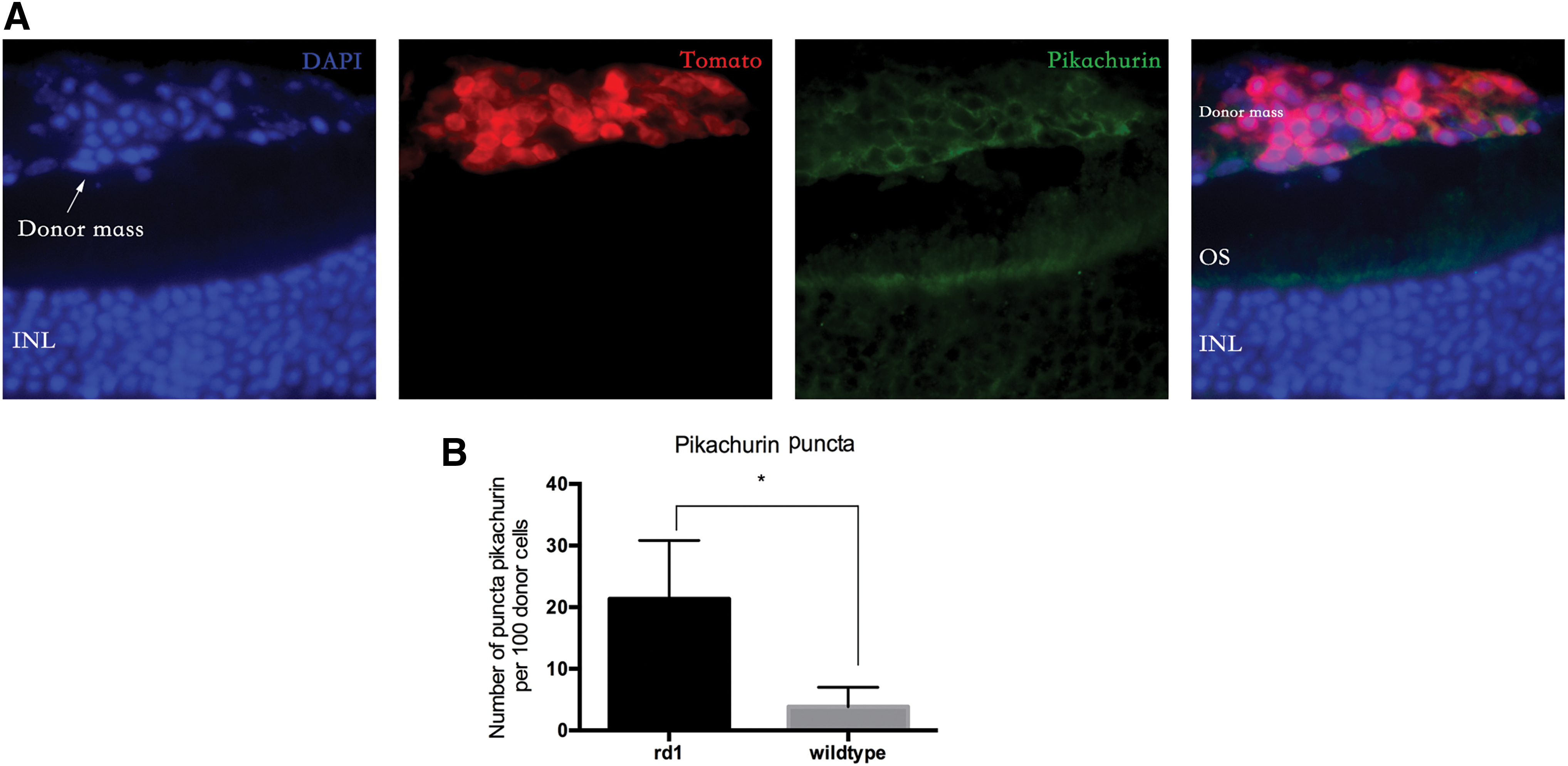
Detection of pikachurin in wild-type mice after photoreceptor transplantation.
In addition, we found that the number of pikachurin puncta was much lower than the number of donor cells. The amount of pikachurin puncta may not be sufficient for all the donor cells to form synapses. Thus, we costained pikachurin with another synaptic marker, bassoon. In confocal images, we found that although most pikachurin puncta were colocalized with bassoon, the bassoon puncta were not always paired with pikachurin (Fig. 6). Quantitative analysis showed that 54.12 ± 8.45% of the bassoon puncta were paired with pikachurin, and 45.5 ± 6.33% of bassoon puncta were not paired. This result indicated that there were fewer pikachurin puncta than bassoon puncta.
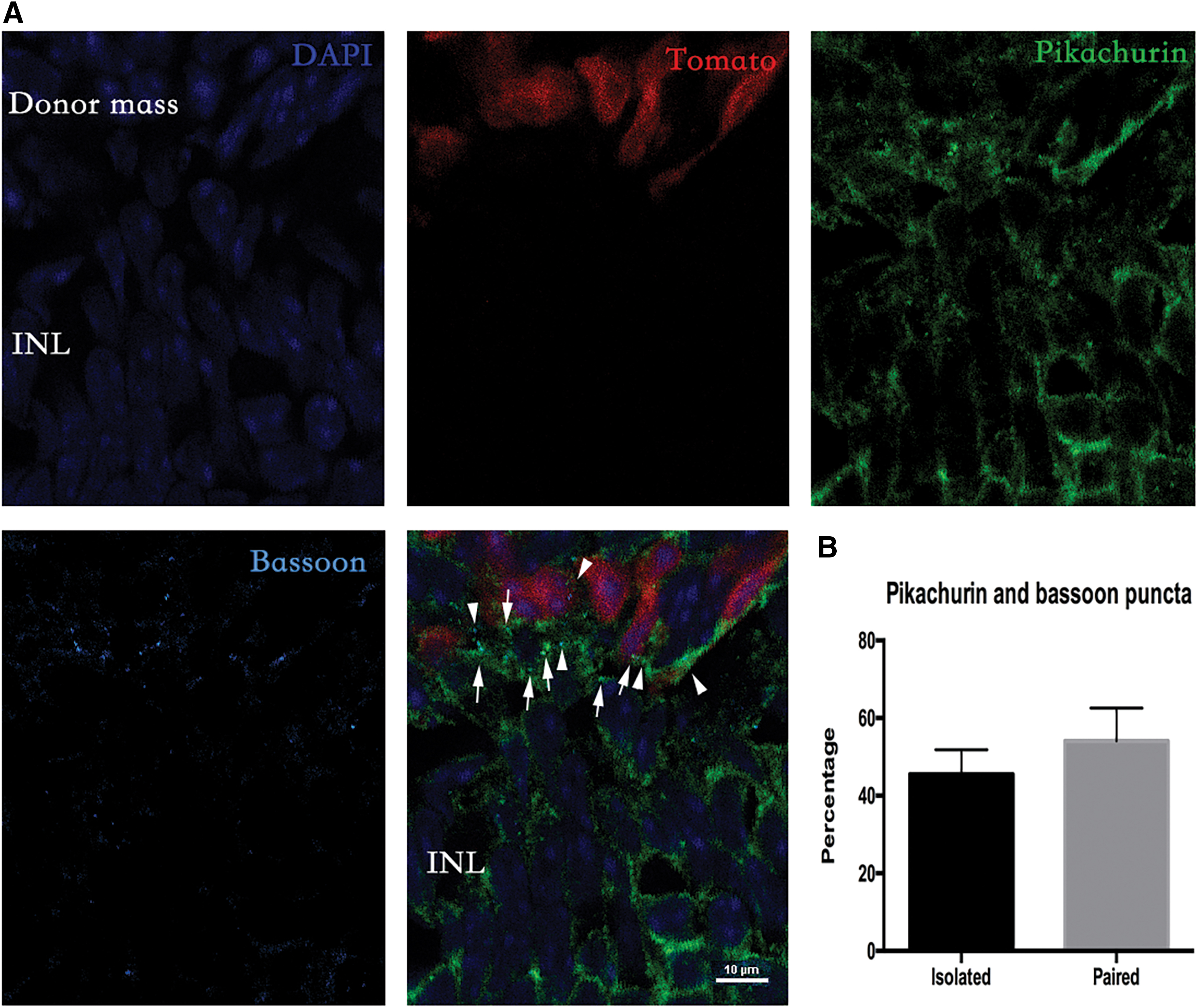
Detection of pikachurin and bassoon in rd1 mice after photoreceptor transplantation.
Discussion
In this study, we investigated whether pikachurin was involved in synapse formation between donor and host cells after photoreceptor transplantation.
Pikachurin is predominantly located at OPL and is exclusively expressed by photoreceptor in wild-type mice [26]. Thus, its expression should decrease as photoreceptors are lost in rd1 mice. However, we found that pikachurin could still be detected in degenerated retina by immunostaining. To confirm what we observed in immunofluorescence examination, western blotting was performed using several different sets of samples. First, we collected the PF and non-PF sorted by FACS from rho-tomato adult mice retina. Comparing these two fractions, the 60 kDa cleaved fragment was found in the former, but not the latter. This result is consistent with previous report [26]. Next, we examined the protein extracted from wild-type and rd1 retinas. We only found the 60 kDa band in wild-type retina. In the rd1 retina, the full-length form of pikachurin was detected (110 kDa) instead. As rd1 mice aged, the amount of the 110 kDa band increased significantly.
We also investigated the pikachurin expression in another retinal degeneration model induced by NMU. Pikachurin was found in INL and GCL, which is similar to the change observed in rd1 mice. These results indicated that the full-length form of pikachurin remained in the retina of rd1 mice after most photoreceptors were lost. Confocal imaging showed that pikachurin was colocalized with a large population of retinal cells, including bipolar cells, amacrine cells, and ganglion cells. However, the reason why the pikachurin could be found in these cells is unknown. Retinal degeneration may lead to unexpected changes of synaptic proteins, for example, the expression level of synaptophysin was increased after most photoreceptors died [31]. In general, we have limited information about the expression of synaptic proteins in degenerated retina.
Next, we transplanted PPCs into the subretinal space of late-stage rd1 mice. As we have observed in degenerated retina before transplantation, pikachurin could be detected in the host retina. Donor cells formed a cell mass in the subretinal space and pikachurin was also found in the donor cells.
Furthermore, we found pikachurin puncta in the donor photoreceptor mass, which was probably its cleaved form. The cleaved N-terminal fragment of pikachurin, not the full-length form, worked as an important matrix protein in triad ribbon synapse [19]. The presence of pikachurin puncta may indicate synapse formation. To verify this speculation, we performed subretinal transplantation in age-matched wild-type mice. Although the graft was close to the INL in rd1 mice, the donor mass was “isolated” from bipolar cell by the ONL in wild-type mice. In the latter situation, pikachurin could be observed in the donor cell cytoplasm, yet puncta-like expression was barely observed. These results probably indicate that the pikachurin puncta found in the subretinal space of rd1 mice was the cleaved form of pikachurin and that it was involved in synapse formation.
However, we noticed that the number of pikachurin puncta was relatively low. It was only one-fifth of the number of donor cells. Thus, we costained pikachurin and another synaptic marker, bassoon. Although most pikachurin puncta was colocalized with bassoon, the bassoon puncta were not always paired with pikachurin. The number of pikachurin puncta was lower than that of bassoon. This result indicates that pikachurin may only involve in a portion of the synaptic connection between donor and host cells.
Several studies have reported functional recovery after photoreceptor transplantation in late-stage rd1 mice, which supports synapse formation between donor and host cells. Singh et al. reported that the end-stage rd1 mice had pupillary light reflex and blood flow in visual cortices after photoreceptor transplantation [7]. According to the authors, ∼2.37 ± 1.95 × 104 cells (7.9 ± 6.5% of 3 × 105 transplanted cells) survived in the subretinal space. Mandai et al. and Iraha et al. transplanted retinal sheets into the degenerated retina, and electrophysiological responses were recorded by multi-electrode array (MEA) [8,13]. Although the number of integrated photoreceptors in the retinal sheet was not accounted for, it should be greater than that using suspension of isolated photoreceptor cells.
Garita-Hernandez et al., recently reported that transplantation of optogenetically engineered photoreceptors could restore the visual function of rd1 mice [12]. However, the host retina grafted with unmodified photoreceptors (control group) failed in the MEA test in this study. The possible reason was that the number of surviving donor cells was insufficient (2830 ± 493 cells per retina, 1.42 ± 0.25% of all transplanted cells) to activate retinal ganglion cells. Thus, the electrophysiological response seems to be donor cell number-dependent in photoreceptor transplantation studies.
With a similar number of optogenetically transformed photoreceptors, however, the function of degenerated retina was restored. Light stimuli greatly promoted the release of glutamate, which may offset the limitation of insufficient cell number. Thus, the recorded electrophysiological response was probably glutamate amount dependent, rather than donor cell number dependent. Alternatively, the activation of the downstream neural circuit is more likely the result of glutamate spillover, but not the signal transmission between matched/paired donor and host cell through mature synaptic connection. In addition, the functional test seems not to be affected in xenotransplantation [8,12], despite interspecific protein structural divergence, such as pikachurin [29].
Taken together, there is a high possibility that the synaptic connections between donor and host cells are immature. This analysis is consistent with our observation that pikachurin was only partially involved in synapse formation after photoreceptor transplantation.
Footnotes
Acknowledgments
The authors thank Christopher R.J. Laver (University of British Columbia) for providing us many advices during the study. The authors are grateful to Yi Zhang and staff from Research Core Facility for the help provided to them in this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants to C.L. from the National Natural Science Foundation of China (81700850) and China Postdoctoral Science Foundation (2018M631085).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.