
Abstract
Bone marrow-derived mesenchymal stem cells (BMSCs) have the potential to differentiate into osteoblasts or adipocytes, and an imbalance between adipogenesis and osteogenesis causes age-related bone loss. In this study, we determined the influence of tumor necrosis factor receptor-associated factor 3 (TRAF3) on senescence and osteoblastic and adipocytic differentiation of rat BMSCs. TRAF3 expression increased during osteogenic differentiation but decreased during adipocytic differentiation of rat BMSCs, and compared with day 0 cultures, on day 14, the differences were significant. Overexpression of TRAF3 significantly promoted BMSC osteogenic differentiation and suppressed adipogenic differentiation and senescence. Furthermore, Traf3 was determined to be a target gene of miR-363-3p in BMSCs, and TRAF3 expression in BMSCs was reduced by miR-363-3p overexpression. This overexpression attenuated the effects of TRAF3 on BMSC adipogenic differentiation, osteogenic differentiation, and senescence. Taken together, these results uncovered the mechanism by which TRAF3 promotes BMSC osteogenic differentiation and suppresses adipogenic differentiation and senescence, indicating that the miR-363-3p–TRAF3 axis might be a novel therapeutic target for BMSC-based bone tissue engineering in osteoporosis.
Introduction
Mesenchymal stem cells are heterogeneous progenitors that have self-renewal capacity and a potential for differentiation into various cell types, such as osteoblasts, adipocytes, and chondrocytes [1 –3]. With age, bone marrow-derived mesenchymal stem cells (BMSCs) show a decreased ability to differentiate into osteoblasts and an increased ability to differentiate into adipocytes, which together cause age-related bone loss [4]. An imbalance between adipogenesis and osteogenesis is associated with various human disorders including unhealthy aging, osteoporosis, and obesity [4]. Nonetheless, the underlying mechanism requires further elucidation.
Tumor necrosis factor receptor-associated factor 3 (TRAF3) is a relatively understudied TRAF family member that regulates the homeostasis of multiple cell types through different mechanisms [5]. Several studies have shown that TRAF3 is associated with bone diseases. For example, Zhao et al. demonstrated that overexpression of microRNA (miRNA, miR) 107 suppresses apoptosis and enhances autophagy in osteoarthritis-affected chondrocytes by targeting TRAF3 mRNA [6]. Xiu et al. generated a mouse model with conditional knockout of TRAF3 in osteoclast precursors; these mice were osteoporotic with greater osteoclast numbers, revealing that TRAF3 has a protective function in physiological bone remodeling [7]. Yao et al. suggested that upregulation of TRAF3 in osteoclast precursors may restrict bone destruction and inflammation-induced bone loss in bone diseases [8]. Recently, Li et al. reported that transforming growth factor (TGF)-β-induced degradation of TRAF3 in mesenchymal progenitor cells results in age-related osteoporosis [9]. Mice with conditional deletion of TRAF3 in mesenchymal cells also developed early-onset osteoporosis owing to a combination of increased osteoclast formation and reduced osteoblast formation [10]. These findings have revealed that TRAF3 suppresses bone loss. Nonetheless, the involvement of TRAF3 in BMSC adipogenesis and osteogenesis remains unclear. Therefore, we aimed to investigate the role of TRAF3 in BMSC adipogenesis and osteogenesis.
miRNAs are short (∼22 nt) noncoding RNAs that repress gene expression through mRNA degradation or translational suppression after binding to the 3′-untranslated region (3′-UTR) of mRNA [11]. Clarification of the molecular mechanisms by which miRNAs regulate osteogenesis and adipogenesis will facilitate development of more effective therapeutic methods for osteoporosis. Several miRNAs involved in the regulation of osteogenic and adipocyte differentiation have been identified. For example, miR-450b accelerates osteogenic differentiation and promotes bone formation by targeting BMP3 mRNA [12]; miR-149-3p reduces adipogenic differentiation and enhances osteogenic differentiation by repressing FTO expression in BMSCs [13]; and miR-188 modulates age-related osteoblastic and adipocytic differentiation [14]. Despite these findings, the participation and molecular mechanisms of action of miRNAs in osteogenic and adipogenic differentiation of BMSCs still require further research. Some studies have shown that miR-363-3p is related to cell differentiation. For instance, miR-363-3p is involved in T helper 17 (Th17) cell differentiation [15] and modulates the transition from mitotic clonal expansion to terminal differentiation [16]. Moreover, miR-363-3p promotes osteoclastogenesis and inhibits osteogenic differentiation [17]. However, its specific functions and molecular mechanisms in osteogenic and adipogenic differentiation of BMSCs remain unclear.
In this study, we demonstrated that overexpression of TRAF3 promotes osteogenic differentiation of BMSCs and suppresses their adipogenic differentiation and senescence. Moreover, we revealed that miR-363-3p overexpression attenuates the effects of TRAF3 on BMSC adipogenic differentiation, osteogenic differentiation, and senescence. These results suggest that TRAF3 may serve as a potential therapeutic target in metabolic bone disorders such as osteoporosis.
Materials and Methods
BMSC culture
Rat BMSCs were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). The BMSCs were grown at 37°C with 5% CO2 and passaged in α-Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS), glutamine, and penicillin–streptomycin sulfate (Invitrogen, Life Technologies, Carlsbad, CA). When the BMSCs reached proper confluence, passages 3–5 were subjected to differentiation in adipogenesis induction medium or osteogenesis induction medium.
Cell transfection
A TRAF3-overexpressing lentivirus and a control empty lentiviral vector were purchased from Hanbio Biotechnology Co., Ltd. (Shanghai, China). The Traf3 sequence was cloned into the pCDH-CMV-MCS-EF1-copGFP vector (System Biosciences, Mountain View, CA), followed by packaging in 293T cells using plasmids psPAX2 and pMD2.G, according to the manufacturer's protocol. Transfection was performed using Lipofectamine 2000 (Invitrogen), and cells were prepared 48 h after transfection. When the cells reached a certain degree of confluence, adipogenic or osteogenic induction was carried out. An miR-363-3p mimic and control mimic (miR-NC) were acquired from RiboBio Co., Ltd. (Guangzhou, China). BMSCs were transfected with RNA using Lipofectamine 2000 (Invitrogen) in accordance with the manufacturer's instructions.
Adipogenic differentiation
For adipogenic differentiation, BMSCs were cultured in adipogenesis induction medium (DMEM containing 10% FBS, insulin, indomethacin, dexamethasone, 3-isobutyl-1-methylxanthine, and dexamethasone), which was refreshed every 3 days. Oil Red O staining was performed to quantify lipid droplets after 14 days of adipogenic induction. BMSCs were fixed in 4% paraformaldehyde, washed, and then stained with an Oil Red O solution. To quantify the retention of Oil Red O, the absorbance was measured at 520 nm after the addition of isopropanol to dissolve the stain.
Osteogenic differentiation
For osteogenic differentiation, BMSCs at 80%–90% confluence were maintained in osteogenesis induction medium, which composed of DMEM with 10% FBS, ascorbic acid, β-glycerophosphate, and dexamethasone. BMSCs were cultured in this medium for 14 days to induce osteoblast mineralization. Alizarin Red S (ARS) staining was performed to assess the mineralization of the cell matrix. ARS was quantitated at 540 nm by spectrophotometry [18].
Alkaline phosphatase activity assay
BMSCs were washed with PBS and lysed using radioimmunoprecipitation assay (RIPA) lysis buffer. The supernatant was incubated with the Alkaline Phosphatase (ALP) Yellow Liquid Substrate System reagent after centrifugation of the cell lysate. The reaction was quantified on a spectrophotometer at 405 nm after blockage with NaOH. ALP activity was normalized to total protein in the cell lysates [19].
Senescence-associated β-galactosidase staining
The activity of senescence-associated β-galactosidase (SA-β-gal) in BMSCs at passage 18 was measured with the Senescence β-Galactosidase Staining Kit (Yeasen, Shanghai, China). In brief, cells were washed with PBS and fixed in 4% paraformaldehyde and then washed and incubated with an SA-β-gal staining solution for 24 h. The cells with SA-β-gal activity were stained blue, and images were captured using an inverted microscope (Olympus, Tokyo, Japan).
RNA extraction and quantitative reverse-transcription PCR analysis
Total RNA was extracted from cells using TRIzol Reagent (Invitrogen), and then cDNA was synthesized with the cDNA Reverse Transcription Kit (Invitrogen). Quantitative reverse-transcription PCR (qRT-PCR) was carried out with Hieff™ qPCR SYBR® Green Master Mix (Yeasen, Shanghai, China) on a StepOnePlus Real-Time PCR Detection System (Applied Biosystems, Foster City, CA). The fold change in miRNA or relative expression of mRNA was determined using the formula 2−ΔΔCt. The levels of β-actin or U6 served as an internal control.
Western blot assay
Total protein was extracted from BMSCs using ice-cold RIPA lysis buffer, resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and transferred to polyvinylidene difluoride membranes. The membranes were incubated with primary antibodies and then with a secondary antibody. After washing, protein expression levels were visualized with an ECL detection system. GAPDH was used as an internal control [20].
Luciferase reporter analysis
BMSCs were seeded in a 24-well plate and then cotransfected with a reporter plasmid [carrying either the wild-type (WT) or mutated (MT) Traf3 3′-UTR] and with either the miR-363-3p mimic or miR-NC. After 48 h of incubation, luciferase assays were performed using the Dual Luciferase Reporter Assay System (Promega, Madison, WI) to measure the firefly and Renilla luciferase activities. The ratio of firefly fluorescence to Renilla fluorescence was calculated and considered the relative luciferase activity.
Statistical analysis
GraphPad Prism software (version 5.0) was used for statistical analyses, and all data are presented as mean ± SD. Student's t-test and one-way analysis of variance were carried out to assess differences between groups. Differences with P < 0.05 were considered statistically significant.
Results
TRAF3 inhibits the adipogenic differentiation of BMSCs
A previous study showed that upregulation of TRAF3 can limit bone destruction and bone loss in common bone diseases [8]; therefore, we hypothesized that TRAF3 may perform some functions in BMSCs. We first examined TRAF3 expression in BMSCs after adipogenic induction and found that mRNA and protein expression of TRAF3 was lower than that in day 0 cultures, and the difference reached significance on day 14 (Fig. 1A, B). To evaluate whether TRAF3 participates in the adipogenic differentiation of BMSCs, we overexpressed TRAF3 in BMSCs using lentivirus. qRT-PCR and western blot assays were performed to confirm the transduction efficiency (Fig. 1C, D). Increased TRAF3 expression markedly reduced lipid droplet formation in BMSCs in the adipogenesis condition (Fig. 1E). We next determined the expression of the adipocyte-related markers peroxisome proliferator-activated receptor γ (Pparγ) and fatty acid-binding protein 4 (Fabp4) in TRAF3 lentivirus-transduced or empty vector-transduced BMSCs after adipogenic induction for 14 days. The results of qRT-PCR and western blot analysis revealed that the expression of Pparγ and Fabp4 was dramatically lower in the TRAF3 lentivirus-transduced BMSCs than in empty vector-transduced BMSCs after adipogenic induction (Fig. 1F, G). These results indicated that TRAF3 suppressed the adipogenic differentiation of BMSCs.

TRAF3 was decreased during adipogenic differentiation, and overexpression of TRAF3 suppressed the adipogenic differentiation of rat BMSCs.
TRAF3 promotes the osteogenic differentiation of BMSCs
To determine whether TRAF3 is involved in osteogenic differentiation, BMSCs were cultured in osteogenesis induction medium for 14 days. During the osteogenic differentiation of BMSCs, the endogenous expression of TRAF3 significantly increased with time compared with that of TRAF3 in day 0 cultures (Fig. 2A, B). As indicated by ARS staining, overexpression of TRAF3 obviously promoted osteoblastic differentiation of BMSCs, resulting in greater calcium deposition than that in cells transfected with the empty vector control (Fig. 2C). In addition, ALP activity was higher in TRAF3 lentivirus-transduced BMSCs than in empty vector-transduced BMSCs (Fig. 2D). Moreover, expression levels of osteogenic differentiation markers ALP, OPN, and RUNX2 were obviously increased by TRAF3 overexpression, as revealed by qRT-PCR and western blot analyses (Fig. 2E–H). These observations suggested that TRAF3 promoted the osteogenic differentiation of BMSCs.
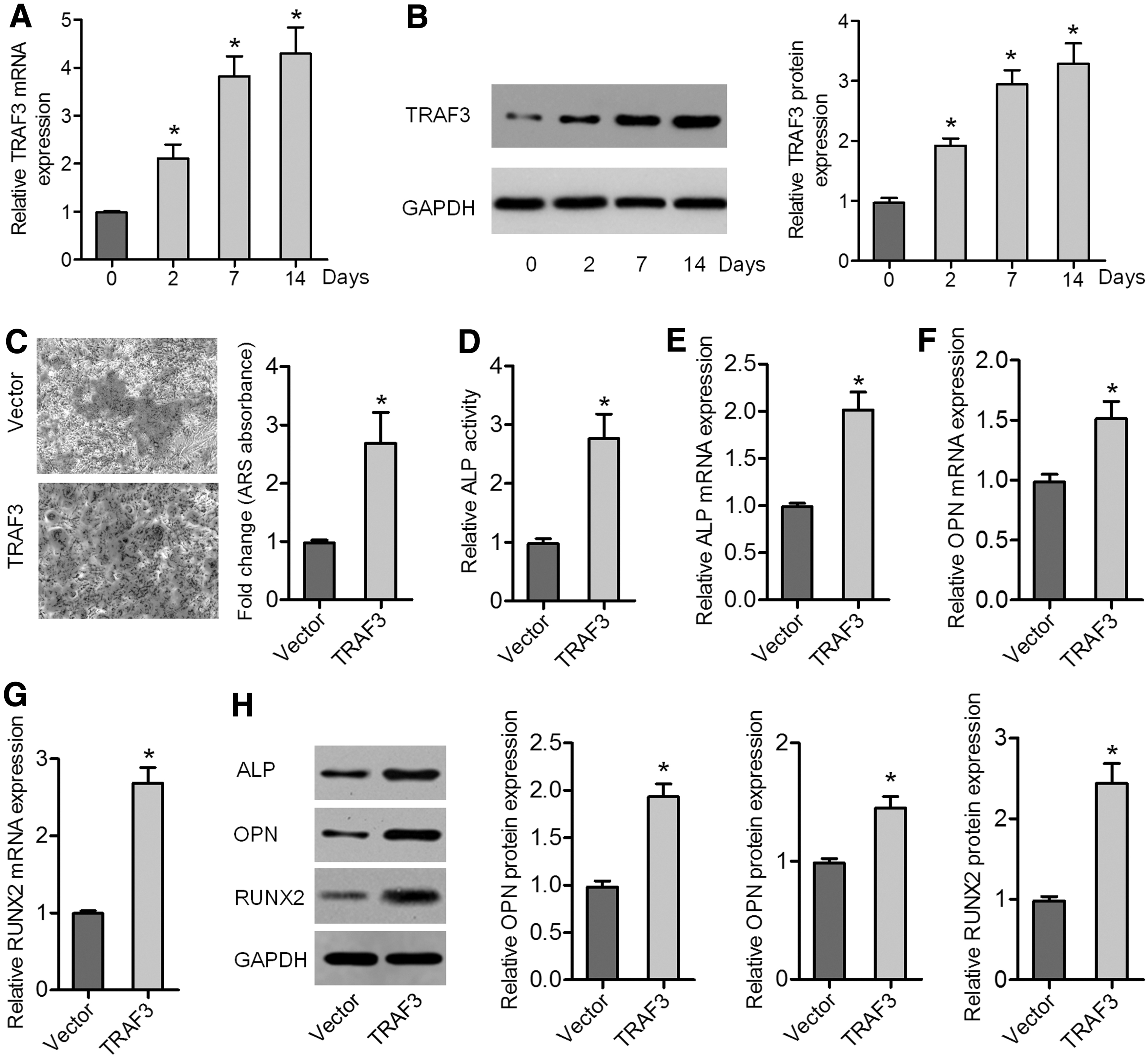
TRAF3 was increased during osteogenic differentiation, and overexpression of TRAF3 increased the osteogenic differentiation of BMSCs.
TRAF3 inhibits the senescence of BMSCs
We next investigated the effect of TRAF3 on BMSC senescence using SA-β-gal staining and qRT-PCR and western blot analyses. We observed a decreased number of SA-β-gal-stained cells after TRAF3 overexpression (Fig. 3A). qRT-PCR and western blot analyses revealed that TRAF3 lentivirus-transduced BMSCs showed lower levels of the senescence markers p16 and p21 than the empty vector-transduced BMSCs (Fig. 3B–D). These data confirmed that TRAF3 is a negative regulator of BMSC senescence.
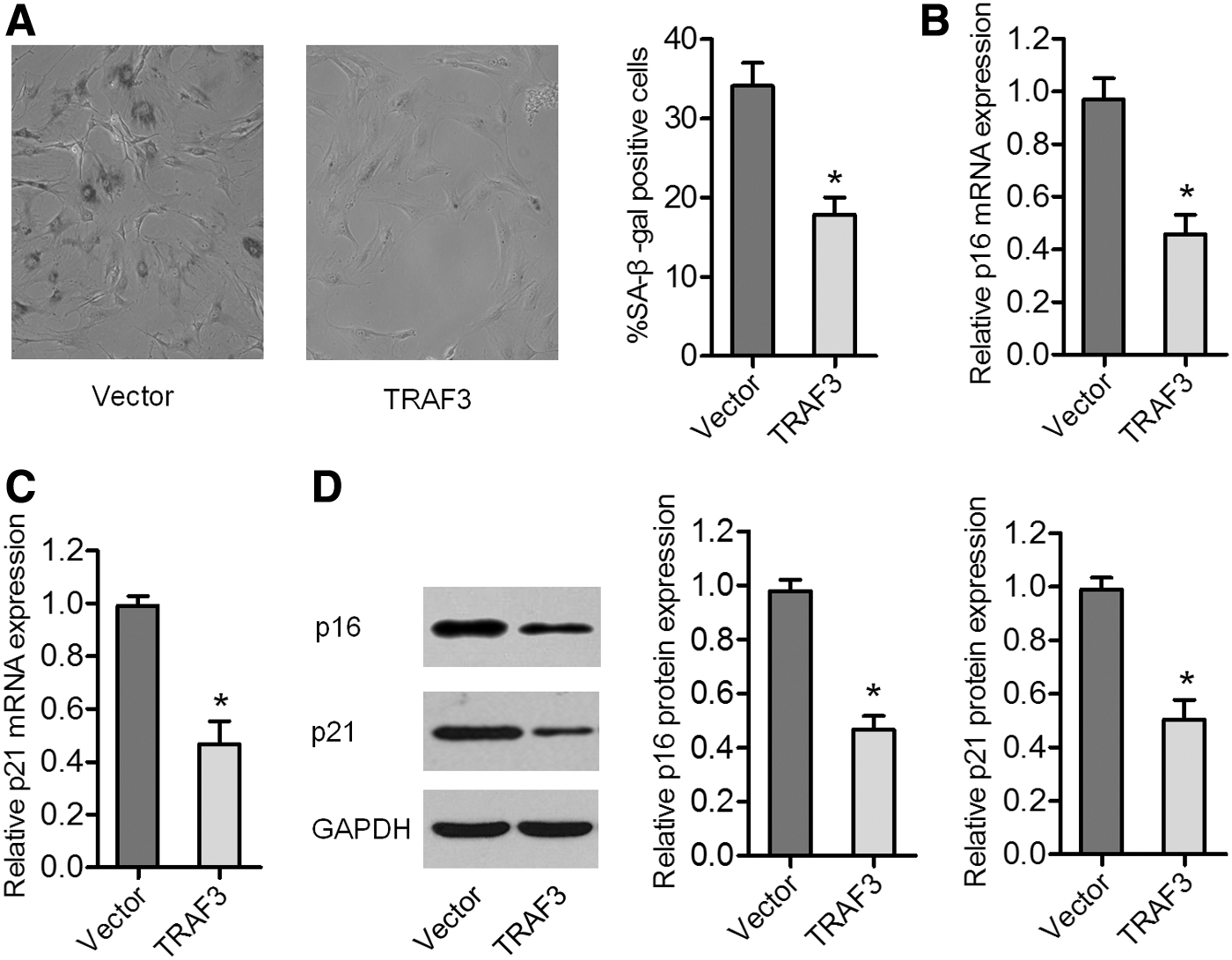
Overexpression of TRAF3 inhibited the senescence of BMSCs.
Traf3 mRNA is targeted and TRAF3 expression is reduced by miR-363-3p in BMSCs
Next, we investigated the mechanism of action of TRAF3 in BMSCs. The miRNAs targeting Traf3 mRNA were predicted by bioinformatics analysis. We screened for specific miRNAs that were simultaneously identified by several databases and focused on miR-363-3p, which is known to be related to osteogenic differentiation [17]. To assess the binding of miR-363-3p to the Traf3 3′-UTR, we carried out a luciferase reporter assay using luciferase reporters containing either the WT or MT Traf3 3′-UTR (Fig. 4A). As given in Fig. 4B, miR-363-3p overexpression dramatically decreased the luciferase activity of the TRAF3-3′UTR-WT reporter plasmid in BMSCs but not that of the TRAF3-3′UTR-MT reporter, in which the putative binding site for miR-363-3p was mutated. In addition, we observed that TRAF3 mRNA and protein expression was decreased in BMSCs by miR-363-3p mimic transfection (Fig. 4C, D). Moreover, miR-363-3p expression significantly increased during adipogenic differentiation and decreased during osteogenic differentiation (Fig. 4E, F). Collectively, these data suggested that the effects of TRAF3 on adipogenic and osteogenic differentiation of BMSCs may be weakened by miR-363-3p.
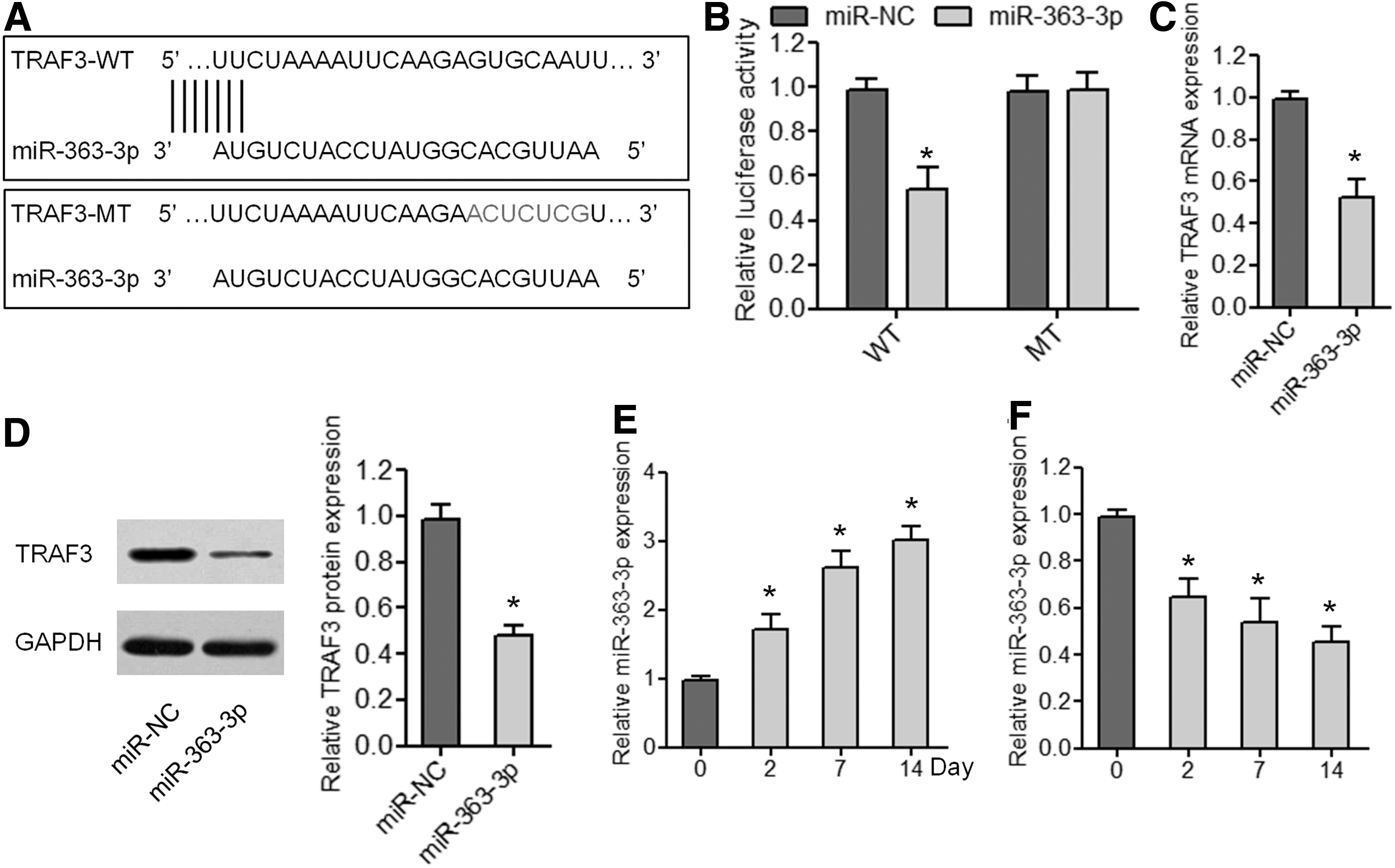
Traf3 is a target gene of miR-363-3p in rat BMSCs.
MiR-363-3p attenuates the effects of TRAF3 on BMSC adipogenic differentiation, osteogenic differentiation, and senescence
To determine whether the TRAF3-induced suppression of BMSC adipogenic differentiation and whether senescence and promotion of osteogenic differentiation were indeed downregulated by miR-363-3p, we transfected the cells with miR-363-3p mimic followed by TRAF3-overexpressing lentiviral vector. We found that TRAF3 overexpression suppressed adipogenic differentiation, but overexpression of miR-363-3p attenuated the inhibitory effect of TRAF3 on the adipogenic differentiation of BMSCs as indicated by restored Oil Red O staining and restored expression of adipocyte-related markers (Fig. 5A, B, H). MiR-363-3p overexpression in BMSCs weakened the promotion of osteogenic differentiation by TRAF3 as confirmed by decreased ARS staining and lower expression of osteogenic differentiation markers (Fig. 5C–E, H). Likewise, miR-363-3p overexpression abrogated the inhibition of BMSC senescence by TRAF3 overexpression as revealed by restored SA-β-gal staining and restored expression of senescence-associated markers (Fig. 5F–H). In addition, upregulation of miR-363-3p promoted adipogenic differentiation and cell senescence and inhibited osteogenic differentiation (Fig. 5A–H). These results suggested that miR-363-3p inhibits the expression and functions of TRAF3 in BMSCs.

Overexpression of miR-363-3p attenuated the effects of TRAF3 on BMSC adipogenic differentiation, osteogenic differentiation, and senescence.
Discussion
In this study, we showed that TRAF3 promotes osteogenic differentiation of BMSCs and suppresses their adipogenic differentiation and senescence. Next, we explored the mechanisms underlying the functions of TRAF3 in the regulation of senescence and osteogenic and adipogenic differentiation. We revealed that miR-363-3p overexpression attenuated the effects of TRAF3 on BMSC adipogenic differentiation, osteogenic differentiation, and senescence. These results indicated that TRAF3 may serve as a potential therapeutic target in osteoporosis.
BMSCs from patients with osteoporosis have a dramatically lower capacity to differentiate into osteoblasts than into adipocytes, resulting in the progression of bone loss [21]. An imbalance between osteogenic differentiation and adipogenic differentiation is related to osteoporosis [22]. Liu et al. reported that miR-214-3p directly targets TRAF3 mRNA to increase osteoclastic and bone resorption activity [23]. Boyce et al. found that TRAF3 limits RANKL-induced osteoclast formation by facilitating the proteasomal degradation of NF-κB-inducing kinase in a complex with TRAF2, indicating that drugs that prevent TRAF3 degradation may be a novel therapeutic approach to bone loss [10]. We speculated that TRAF3 influences bone formation by modulating osteogenic and adipogenic differentiation. In this study, we showed that TRAF3 expression increased during osteogenic differentiation but decreased during adipogenic differentiation of BMSCs. It is known that decreased osteoblastic activity and increased osteoclastic activity contribute to age-related bone loss [24]. The multilineage differentiation potential of senescent BMSCs is impaired, thus decreasing the rate of bone formation and increasing predisposition to osteoporosis [25]. When compared with those of earlier passages, late-passage human adipose-derived mesenchymal stem cells (P15 or later) displayed a senescent phenotype and decreased osteogenic potential [19]. Thus, we speculated that TRAF3 may play a role in BMSC senescence. Our study demonstrated that TRAF3 overexpression dramatically enhanced osteogenic differentiation of BMSCs and suppressed their adipogenic differentiation and senescence.
We next explored the mechanisms underlying the functions of TRAF3 in the modulation of senescence and of osteogenic and adipogenic differentiation. We found that Traf3 is a target gene of miR-363-3p. Moreover, the luciferase reporter, qRT-PCR, and western blot assays suggested that miR-363-3p overexpression decreased TRAF3 expression by binding of miR-363-3p to the Traf3 mRNA 3′UTR. Some studies have shown that miR-363-3p is associated with cell differentiation. For example, miR-363-3p promotes pluripotency maintenance through repression of NOTCH1 and PSEN1 and suppression of Notch-induced differentiation of human pluripotent cells [26]. Dendritic cells from peripheral blood of patients with rheumatoid arthritis promote Th17 cell differentiation through the miR-363-3p–integrin αv–TGF-β axis [27]. Moreover, during adipogenesis of adipose tissue-derived stromal cells, miR-363-3p modulates the transition from mitotic clonal expansion to terminal differentiation by suppressing E2F3 [16]. Our study suggests that miR-363-3p overexpression dramatically reduces the effects of TRAF3 on BMSC senescence and osteogenic and adipogenic differentiation. In addition, we demonstrated that miR-363-3p overexpression suppresses osteogenic differentiation, consistent with the recent findings of Li et al. [17]. These data suggest that miR-363-3p may suppress the expression and functions of TRAF3 in BMSCs.
In conclusion, our findings indicate that miR-363-3p modulates BMSC differentiation and promotes senescence by downregulating TRAF3. These data reveal a new mechanism behind the osteoporosis-related imbalance between osteogenic differentiation and adipogenic differentiation and indicate that the miR-363-3p–TRAF3 axis may be a novel therapeutic target for BMSC-based bone tissue engineering in osteoporosis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.