
Abstract
Mesenchymal stem cells (MSCs) are multifunctional stem cells that exist in almost all human tissues. In addition to their self-renewal and multidirectional differentiation potential, they also have valuable immunomodulatory abilities. Bone marrow mesenchymal stem cells (BMMSCs) are the first discovered MSCs and are the most widely studied. Stem cells from the apical papilla (SCAP) are derived from the apical papilla of incompletely developed teeth and play an important role in the formation and development of tooth root. Recent studies have shown that mesenchymal stem cell-derived exosomes (MSC-exo) have similar biological functions as MSCs. Moreover, increasing evidence has highlighted the functional relationship between noncoding regulatory RNAs, especially microRNAs, and MSC-exo. However, few studies have addressed the role of PIWI-interacting RNAs (piRNAs) in MSC-exo. To develop a better understanding of the biological functions of SCAP and BMMSCs, we compared and analyzed the piRNA expression profiles of the exosomes derived from human SCAP (SCAP-exo) and the exosomes of BMMSCs (BMMSC-exo). A total of 593 and 920 known piRNAs were identified from SCAP-exo and BMMSC-exo, respectively, and 21 piRNAs were found to be differentially expressed. In addition, we predicted the target genes of the differentially expressed piRNAs, and the target genes were subjected to the Gene Ontology enrichment and the Kyoto Encyclopedia of Genes and Genomes pathway analysis, revealing the possible biological functions of these differentially expressed piRNAs. We found that the target genes of the differentially expressed piRNAs mainly involved in biological regulation, cellular processes, metabolic processes, binding, and catalytic activity, which are closely related to the biological functions of MSCs. In conclusion, this study confirmed the differential expression profiles of piRNAs in SCAP-exo and BMMSC-exo and provided useful insights for further study of their functions.
Introduction
Mesenchymal stem cells (MSCs) are self-renewing multipotent progenitors that can be isolated from various tissues [1,2]. They have multiple biological functions, especially in the area of tissue regeneration and the treatment of autoimmune diseases [3,4]. Bone marrow mesenchymal stem cells (BMMSCs) are one of the most widely studied MSCs; these mesodermal-derived stem cells exhibit multidirectional differentiation potential and play a key role in immunomodulation. They have been used in the treatment of diseases such as autoimmune diseases, hypertension, and spinal cord injuries [5]. However, acquisition of BMMSCs requires highly invasive surgery, and the longevity and potential for differentiation diminish with donor age [6].
Recently, dental stem cells (DSCs) have gained more attention, which are rich in source and retrieved through noninvasive procedures. As a type of DSCs, stem cells from the apical papilla (SCAP) were isolated from human apical papilla tissues for the first time in 2006 by Sonoyama et al. [7]. SCAP possess many similar features to BMMSCs in vitro, including the expression of certain markers, and the ability to differentiate into cells that have the characteristics of osteoblasts, chondrocytes, and adipocytes [8]. Moreover, SCAP are derived from developing tissue, apical papilla, which showed a superior tissue regeneration potential than other DSCs and BMMSCs, because they have high proliferative potential, reflected in high telomerase activity [9,10]. Our previous research found that SCAP could promote CD4+CD25− effector T cells conversion to CD4+CD25+Foxp3+ regulatory T cells, which further disclosed the immunomodulatory properties of SCAP [11]. The reasons for the functional differences between BMMSCs and SCAP are not only the focus of research but also the premise of their translation in clinic. The biological safety issues, such as tumorigenicity and retention associated with stem cell therapy, are severely limiting their clinical applications [12].
Exosomes are small extracellular vesicles of 30–150 nm in diameter and are produced by most of cell types. Recent studies have shown that exosomes derived from MSCs (MSC-exo) have similar functions to MSCs, including promoting tissue repair and regeneration, inhibiting inflammatory response, and regulating immunity [13 –16]. Compared with MSCs, MSC-exo are not only easier to store but also a safer option to covey the therapeutic potential of MSCs. Exosomes contain a variety of active components, including lipids, proteins, DNA, noncoding RNAs (ncRNAs), and RNA. ncRNAs in exosomes are considered to be one of the most important molecules responsible for regulating recipient cells [17 –19]. MSC-exo have been shown to promote angiogenesis and tissue regeneration through their ncRNAs and proteins [19 –22].
PIWI-interacting RNAs (piRNAs) are a new class of small ncRNAs that were discovered in 2006, which bind to PIWI-family subproteins to form RNA-induced silencing complexes. piRNAs play an important role in maintaining genomic stability and spermatogenesis in mammals by regulating the activity of target genes and transposons [23 –26]. Research into piRNAs has mainly focused on understanding the mechanisms that maintain germ line integrity. More recently, roles for piRNAs in gene regulation have emerged. Some scholars have found various piRNAs in the exosomes of endothelial cells and saliva [17,27]. However, there are no reports on the expression profiles of piRNAs in MSC-exo.
In this study, we analyzed and compared piRNA expression profiles in the exosomes of SCAP (SCAP-exo) and the exosomes of BMMSCs (BMMSC-exo). The aim of this study was to enrich our understanding of the functions of SCAP and BMMSCs, and the possible mechanisms involved in tissue regeneration and immune regulation, and to provide useful resources for further investigations into their functioning.
Materials and Methods
Source of MSCs
This study was approved by the ethics committee of the School of Stomatology, China Medical University (201315). SCAP were collected from intact, caries-free impacted third molars (n = 3) with immature roots from three healthy human patients (12–15 years of age) at the dental clinic of the School of Stomatology affiliated with the China Medical University. BMMSCs were purchased from ScienCell Company. The cells were seeded in 10-cm culture dishes and cultured with alpha minimum essential medium (α-MEM; Hyclone) supplemented with 15% fetal bovine serum (FBS; MRC BRL), 2 mM L-glutamine (Biosource/Invitrogen), 100 U/mL penicillin–streptomycin (Hyclone), and 0.1 mM L-ascorbic acid 2-phosphate (WAKO, Japan) and maintained in a humidified atmosphere containing 5% CO2 at 37°C. Cells at passage 5 were used for subsequent experiments. When the fusion degree of the 5th passage cells reached 80%, these cells were washed three times with phosphate-buffered saline (PBS) and the supernatant was collected after 48 h of continuous culture in exosome-free serum medium (SBI) and was prepared for the extraction of exosomes.
Multilineage differentiation assay
SCAP and BMMSCs at the fifth passage were seeded into six-well plates and cultured to 80% confluence. Osteogenesis culture medium supplemented with 1.8 mM monopotassium phosphate (Sigma–Aldrich) and 10 nM dexamethasone (Sigma–Aldrich) was exchanged for 6 weeks. The specimens were stained with 1% Alizarin red S. The adipogenic differentiation assay included adipogenesis culture medium with 500 μM isobutyl-methylxanthine (Sigma–Aldrich), 0.5 μM hydrocortisone (Sigma–Aldrich), 60 μM indomethacin (Sigma–Aldrich), and 10 μM insulin (Sigma–Aldrich) replaced two or three times per week for 3 weeks. Oil red O was used to stain the oil droplets.
Surface markers expressed on SCAP and BMMSCs by flow cytometry
Anti-human CD73 antibody labeled with phycoerythrin (PE) and anti-human CD31, CD34, CD45, CD90, and CD105 (1:100; Abcam) antibody labeled with fluoresceine isothiocyanate (FITC) were used to detect the surface markers expression on SCAP and BMMSCs. SCAP and BMMSCs were incubated with CD31, CD34, CD45, CD73, CD90, and CD105 antibodies, respectively, on ice for 1 h. The expression of each surface marker was tested by flow cytometry (Becton Dickinson, Islandia, NY).
Isolation and identification of exosomes
The exosomes were purified according to the ExoQuick-TC™ kit (SBI) instructions. Ten milliliters of cell culture supernatant was collected from each sample and centrifuged at 3,000 g for 15 min at 4°C to remove cells and cell debris. The supernatant was transferred to a 15-mL centrifuge tube, placed on ice, and an appropriate volume of ExoQuick-TC exosome precipitation solution was added. The centrifuge tube was inverted to mix the liquid and refrigerated overnight. The ExoQuick-TC/supernatant mixture was centrifuged at 3,000 g for 15 min at 4°C. After centrifugation, the exosomes appeared as a white pellet at the bottom of the tube. The supernatant was aspirated, and the residual ExoQuick-TC solution was rotated by centrifugation at 1,500 g for 5 min, and all fluid traces were removed by aspiration. The exosomal pellet was resuspended in 200 μL of PBS.
The extracted exosomes were diluted at a ratio of 1:20. Ten microliters was dropped onto a copper net, placed in a 65°C oven for 30 min, dried, and labeled with phosphotungstic acid, before being dried naturally and viewed by transmission electron microscopy (TEM). The size of these exosomes was analyzed by nanoparticle tracking analysis.
Exosomes were incubated with RIPA lysis buffer (Beyotime Biotech Co., Shanghai, China) containing 1 mM/L phenylmethane-sulfonyl fluoride on ice for 1 h. A bicinchoninic acid protein assay kit (Beyotime Biotech Co.) was used to quantify the concentration of protein. An equal amount of exosome lysates was separated with 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Millipore Corporation, Billerica, MA). The membranes were blocked in 4% bovine serum albumin for 1 h at room temperature and then incubated with Alix anti-rabbit primary antibody (1:500; Abcam), CD9 anti-rabbit primary antibody (1:500; Abcam), CD63 anti-rabbit primary antibody (1:500; Proteintech), or Calnexin anti-rabbit primary antibody (1:500; Abcam) at 4°C overnight. The membranes were washed with tris buffered saline with tween 20 and exposed to goat anti-rabbit IgG IRDyel 800CW secondary antibody (1:1,000; Abbkine, Inc., Redlands, CA) at room temperature for 1 h. The protein bands were detected with an Odyssey CLx instrument (LI-COR, Lincoln, NE), and grayscale analysis was performed using the ImageJ software (1.50i; National Institutes of Health, Bethesda, MA).
RNA extraction and construction of small RNA library
The total RNA of SCAP-exo and BMMSC-exo was isolated separately using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. The concentration and quality of each RNA sample were evaluated using a NanoDrop spectrophotometer (NanoDrop ND-1000; Thermo Fisher Scientific) and 1% agarose gel electrophoresis. Then, reverse transcription–polymerase chain reaction (RT-PCR) was performed to create cDNA based on the small RNA (sRNA), and the libraries were amplified to produce stable and easy-to-use libraries for sequencing. Polyacrylamide gel electrophoresis was used to recover the sRNA library of about 150 bp. One microliter of RNA library was assessed with a Qubit Fluorometer (Invitrogen) to determine the concentration, and only libraries with a concentration greater than 1.0 ng/L were qualified. Bio-Q100 was used to detect the length distribution and concentration of the library.
sRNA sequencing and annotation
Each sRNA library was added to the Illumina HiSeq X Ten-sequencing platform for high-throughput parallel sequencing. The sequences obtained by Illumina HiSeq sequencing were filtered through removing joints and low-quality sequences to obtain clean sequences. The clean sequences were compared with the Rfam database (
Analysis of differentially expressed piRNAs
The piRNA expression profiles in each library were normalized to identify differentially expressed piRNAs. piRNA expression levels were estimated by the number of reads per million (RPM) using the following formula: RPM = each piRNA reads/all piRNA reads × 106. The results were analyzed by using DESeq2 (Version 1.20.0) according to the expression of the piRNAs. The standard of difference was | log2 (fold change) | > 1 as well as P < 0.05. The differentially expressed piRNAs were exported as volcano plots, and heat maps were generated based on their expression levels.
Prediction and enrichment analysis of target genes
The target genes of differentially expressed piRNAs were predicted using miRanda (v3.3a,
Validation of crucial differentially expressed piRNAs via quantitative PCR
To validate the sRNA sequencing results, we selected two differentially expressed piRNAs between SCAP-exo and BMMSC-exo to perform stem-loop reverse transcription–quantitative polymerase chain reaction (qPCR). The GAPDH gene sequence was used as a control. The primer sequences were designed by primer bank (Supplementary Table S1).
A chromo4 RT-PCR system (Bio-Rad) was used to perform the qPCR with a SuperReal PreMix Plus Kit (Tiangen, China) following the manufacturer's instructions. A total of 1,000 ng RNA was used to synthesized cDNA, and 200 ng cDNA was used in a 20 μL qPCR mixture containing 10 μM FW primer, 10 μM RV primer, and 10 μL SYBR Green Supermix for each reaction. Amplification was performed as follows: 95°C for 10 min, followed by 40 cycles at 94°C for 15 s, 60°C for 30 s, and 70°C for 30s. All reactions were performed in triplicate, and the data were analyzed by the 2−▵▵CT method.
Results
Identification of SCAP and BMMSCs
The fifth passage SCAP presented with a spindle-like appearance (Supplementary Fig. 1A), and the fifth passage BMMSCs appeared as short spindle or polygon (Supplementary Fig. 2A). Both SCAP and BMMSCs possessed osteogenic and adipogenic differentiation potential (Supplementary Figs. 1B, C and 2B, C). In addition, flow cytometry revealed that SCAP and BMMSCs expressed MSC markers (CD73, CD90, and CD105) but failed to express hematopoietic stem cell markers (CD31, CD34, and CD45) (Supplementary Figs. 1D and 2D).

Characterization of exosomes derived from SCAP and BMMSCs.
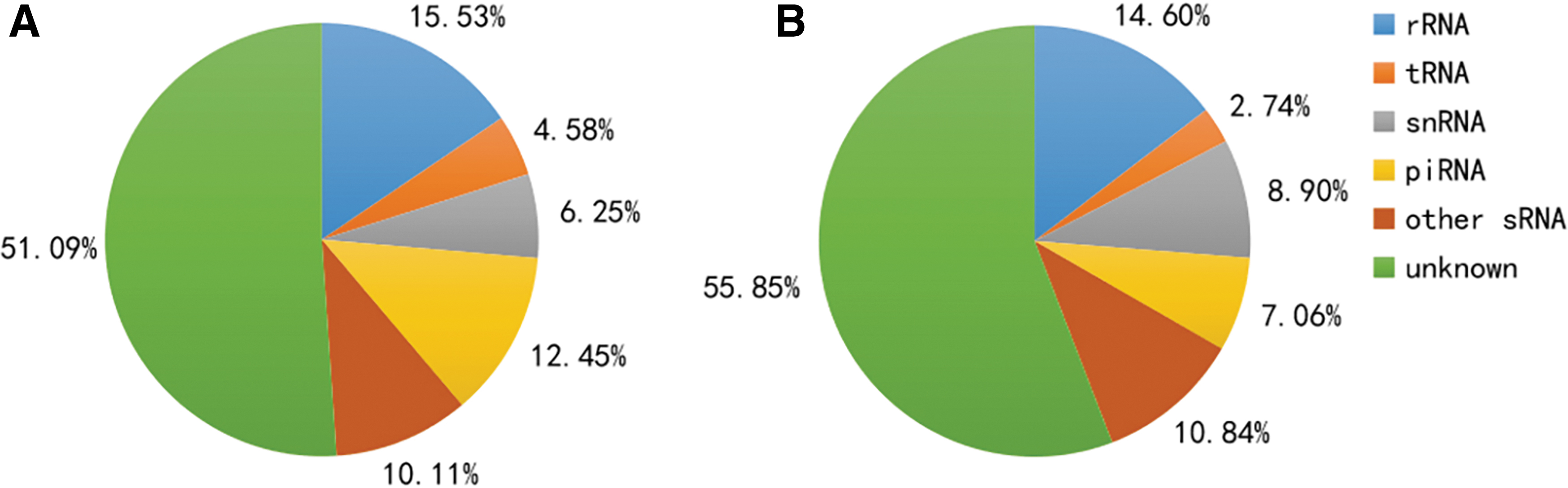
Small RNA classification. Each color represents a kind of small RNA, which includes rRNA, tRNA, snRNA, piRNA, other sRNA, and unknown sRNA.
Identification of exosomes derived from SCAP and BMMSCs
Exosomes were characterized according to MISEV 2018 guidelines [28]. TEM imaging showed that SCAP-exo and BMMSC-exo were cup-shaped, and there were no morphological differences between them (Fig. 1A, B). Nanoparticle tracking analysis (NTA) indicated that the diameters of SCAP-exo and BMMSC-exo ranged approximately from 30 to 150 nm, with a major peak in particle size at 119.1 and 120.5 nm, respectively (Fig. 1C, D). Western blot analysis showed that exosomes had immunoreactivity with specific antibodies against CD9, CD63, and Alix but failed to express Calnexin (Fig. 1E, F).
Overview of the sRNAs sequencing data
After Illumina HiSeq X Ten sequencing, 17.30 × 106 and 20.18 × 106 raw reads were obtained from the two sRNA libraries of SCAP-exo and BMMSC-exo, respectively. The low-quality reads were removed, and the 3′- and 5′-adaptor sequences were trimmed. In total, we obtained 11.78 × 106 and 2.10 × 106 clean reads from the two libraries, respectively. Then, the clean reads were compared with the Rfam database and piRNA bank to obtain the sRNA classification and annotation results for each sample and counts separately. The average length of these sRNA ranged from 15 nt to 28 nt.
The classification results are summarized in Fig. 2; they showed the proportion of rRNA, tRNA, small nuclear RNA (snRNA), piRNA, other sRNAs, and unknown RNA in the sRNA libraries of SCAP-exo and BMMSC-exo. The distributions of sRNA-mapping reads between the two groups were similar, and more than 50% of the sRNA was unknown sRNA. Among the known sRNAs, the percentage of tRNA was the highest. The proportion of piRNA was 12.45% and 7.06% in SCAP-exo and BMMSC-exo, respectively.
piRNA expression profiles
Overall, 593 and 920 known piRNAs were identified from the SCAP-exo and BMMSC-exo. The length of these piRNAs ranged from 15 nt to 28 nt, with the distribution peak at 21 nt. In SCAP-exo, the read counts of hsa-piR-020326 were much higher than other piRNAs, and hsa-piR-016735 and hsa-piR-017716 also had high abundances. However, in BMMSC-exo, the highest read count was hsa-piR-016735. The 10 most highly expressed piRNAs of SCAP-exo and BMMSC-exo are shown in Table 1.
Ten Most Highly Expressed PIWI-Interacting RNAs in Stem Cells from the Apical Papilla-exo and Bone Marrow Mesenchymal Stem Cell-exo
BMMSC, bone marrow mesenchymal stem cell; SCAP, stem cells from the apical papilla.
Differentially expressed piRNAs in SCAP-exo and BMMSC-exo
Based on the criteria of log2 (fold change) greater than 1 or less than −1, as well as P-values less than 0.05, we detected 21 piRNAs that were differentially expressed in SCAP-exo and BMMSC-exo. Compared with BMMSC-exo, 15 piRNAs were upregulated and 6 piRNAs were downregulated in SCAP-exo (Table 2). Volcano plots and a heat map were used to better visualize the differential expression of piRNAs (Fig. 3). Each differentially expressed piRNA was shown in the heat map, which represents a comparison between the piRNA expression levels.
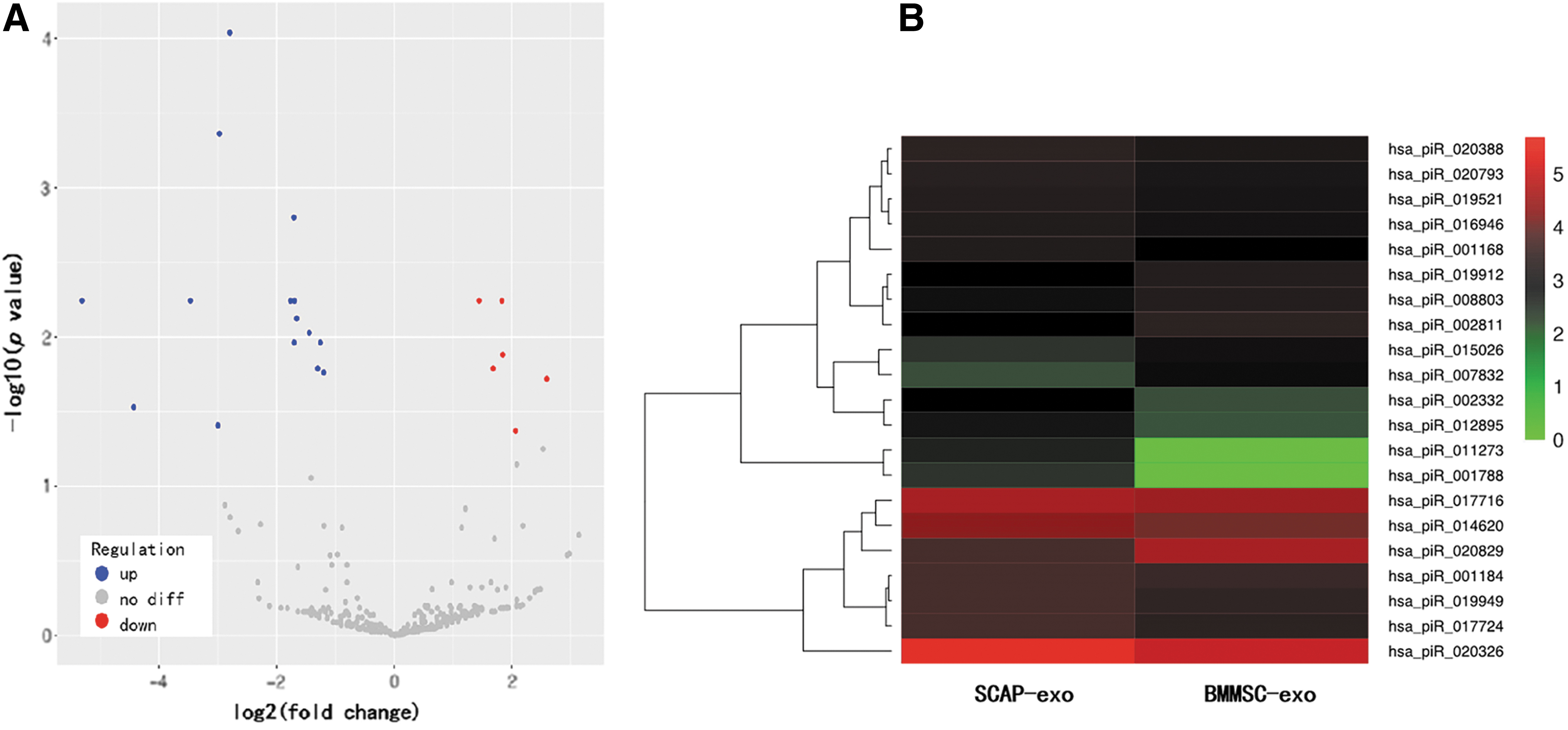
Volcano plots and heat map of differentially expressed piRNAs. The abscissa corresponds to log2 (fold change), ordinates represent P values. Each point represents a piRNA. Blue points represent significantly upregulated piRNA expression, red points represent significantly downregulated piRNA expression, and gray points represent no differentially expressed piRNAs.
Differentially Expressed PIWI-Interacting RNAs Between Stem Cells from the Apical Papilla-exo and Bone Marrow Mesenchymal Stem Cell-exo
(log2 (fold change) >1.0 or log2 (fold change) <−1.0, P < 0.05).
GO analyses of differentially expressed piRNAs
The GO analysis results showed that the target genes of the differentially expressed piRNAs were significantly involved in a broad range of biological processes, cellular components, and molecular functions. First, in regard to the target genes of those upregulated piRNAs in SCAP-exo compared with BMMSC-exo, GO enrichment results showed that single-organism processes were the most significant enrichment term in the biological process category. Within the cellular component category, most of the target genes were associated with cell parts. Meanwhile, in the molecular function category, binding was identified as the most significant enrichment term (Fig. 4A). Second, target genes of those downregulated piRNAs in SCAP-exo were most significantly enriched in cellular processes, cell parts, and binding, in that order (Fig. 4B).
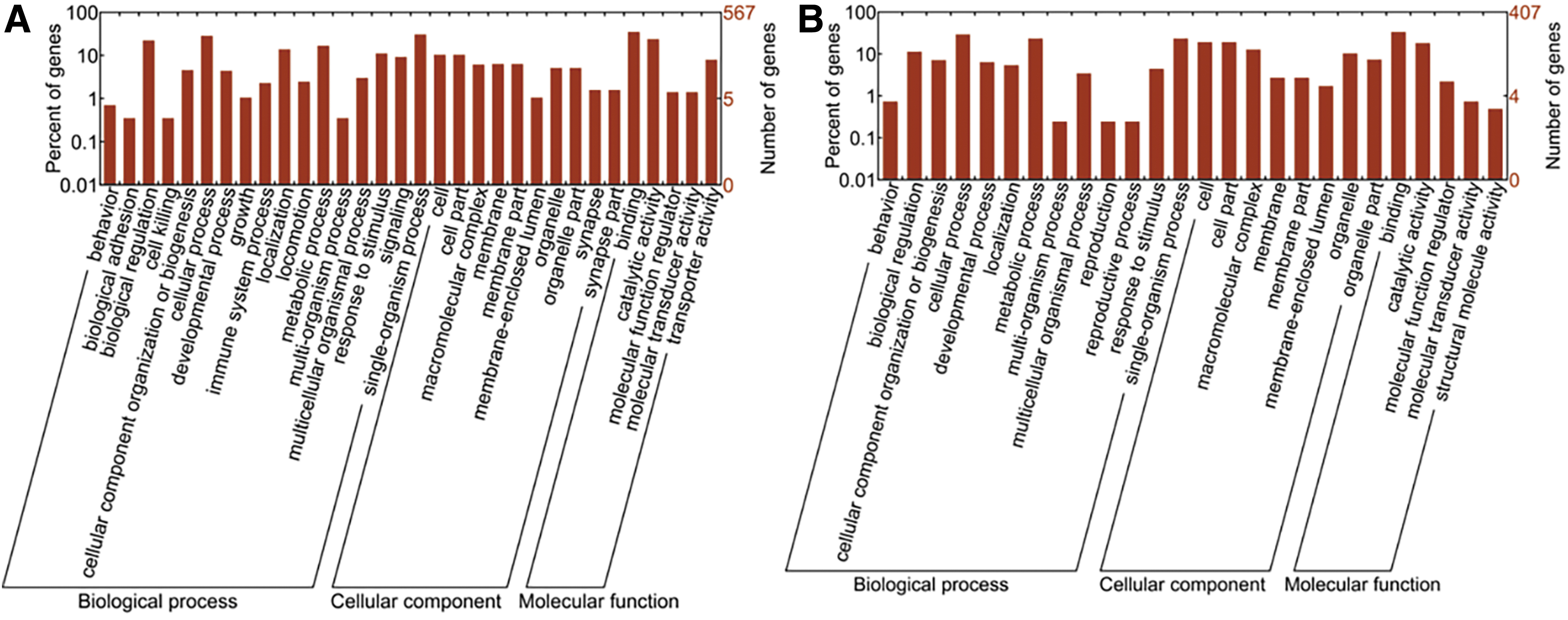
GO enrichment analysis of the target genes of the differentially expressed piRNAs. The abscissa is the GO classification, which is the secondary function of GO. The left side of the ordinate is the percentage of the target genes, and the right side is the number of target genes.
KEGG pathway enrichment of differentially expressed piRNAs
The KEGG pathway analysis revealed that those target genes of differentially expressed piRNAs were significantly enriched in multiple pathways involved in carbohydrate, lipid, and amino acid metabolism. In addition, target genes of the upregulated piRNAs in SCAP-exo compared with BMMSC-exo were significantly enriched in the mitogen-activated protein kinase (MAPK) signaling pathway, Ras signaling pathway, and citrate cycle signaling pathway (Fig. 5A). The target genes of those downregulated piRNAs in SCAP-exo were enriched in the p53 signaling pathway and Epstein–Barr virus infection signaling pathway (Fig. 5B).
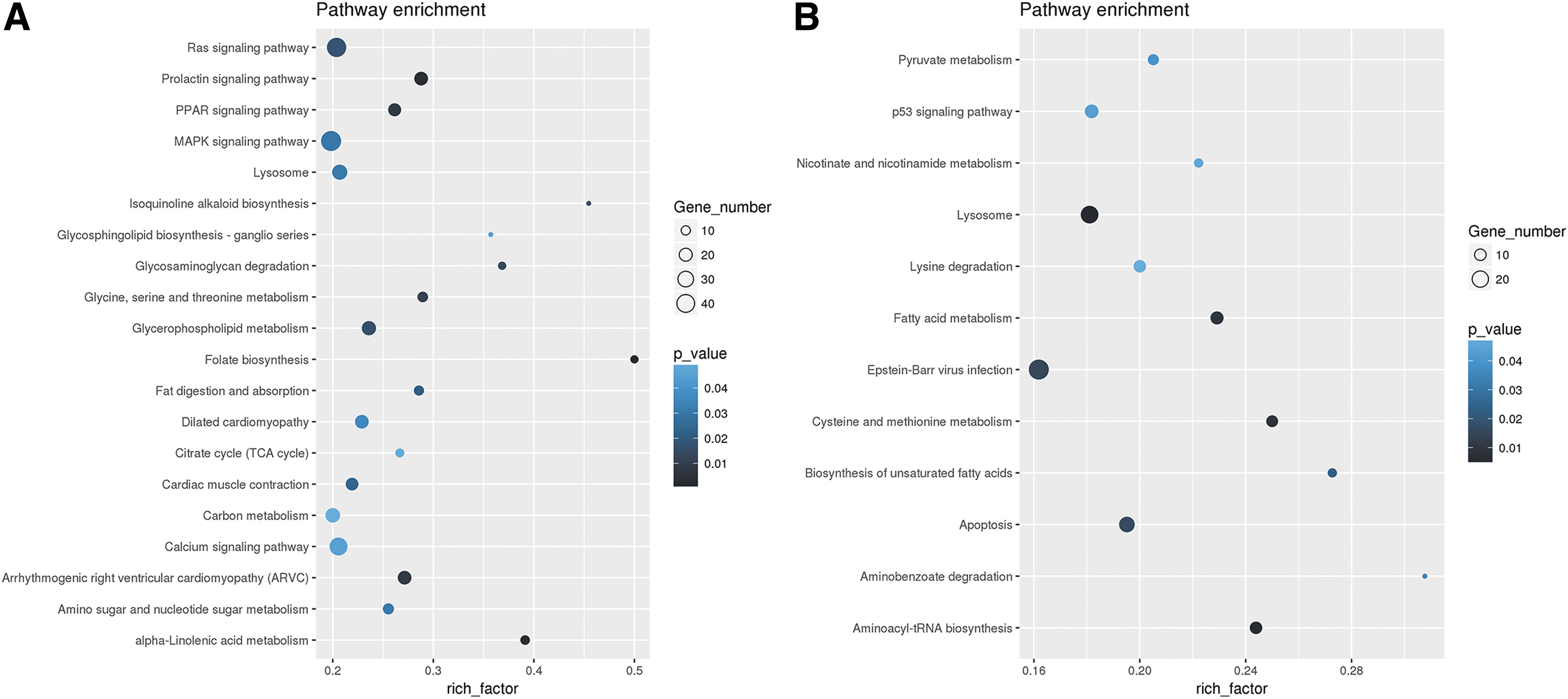
KEGG enrichment analysis of the target genes of the differentially expressed piRNAs. The abscissa is the enrichment factor (the number of different piRNA target genes enriched in the pathway/the number of background genes in the pathway); the larger the number, the greater the enrichment degree. The ordinate is the corresponding pathway. The bubble size is the number of target genes; the larger the bubble, the more genes there are. The darker the bubble color, the smaller the P value, which indicates that the enrichment is more obvious.
Validation of sequencing data by qPCR
To verify the existence and expression patterns of the piRNAs from the sequencing data, hsa-piR-020388 and hsa-piR-020326, with different expression patterns from the deep sequencing results, were randomly selected for qPCR analysis. The qPCR analyses showed that the log2 (fold change) value between SCAP-exo and BMMSC-exo of hsa-piR-020388 and hsa-piR-020326 was 3.81 and 5.25, respectively (Fig. 6). Therefore, the qPCR data were consistent with the deep sequencing results. Both hsa-piR-020388 and hsa-piR-020326 were upregulated in SCAP-exo.
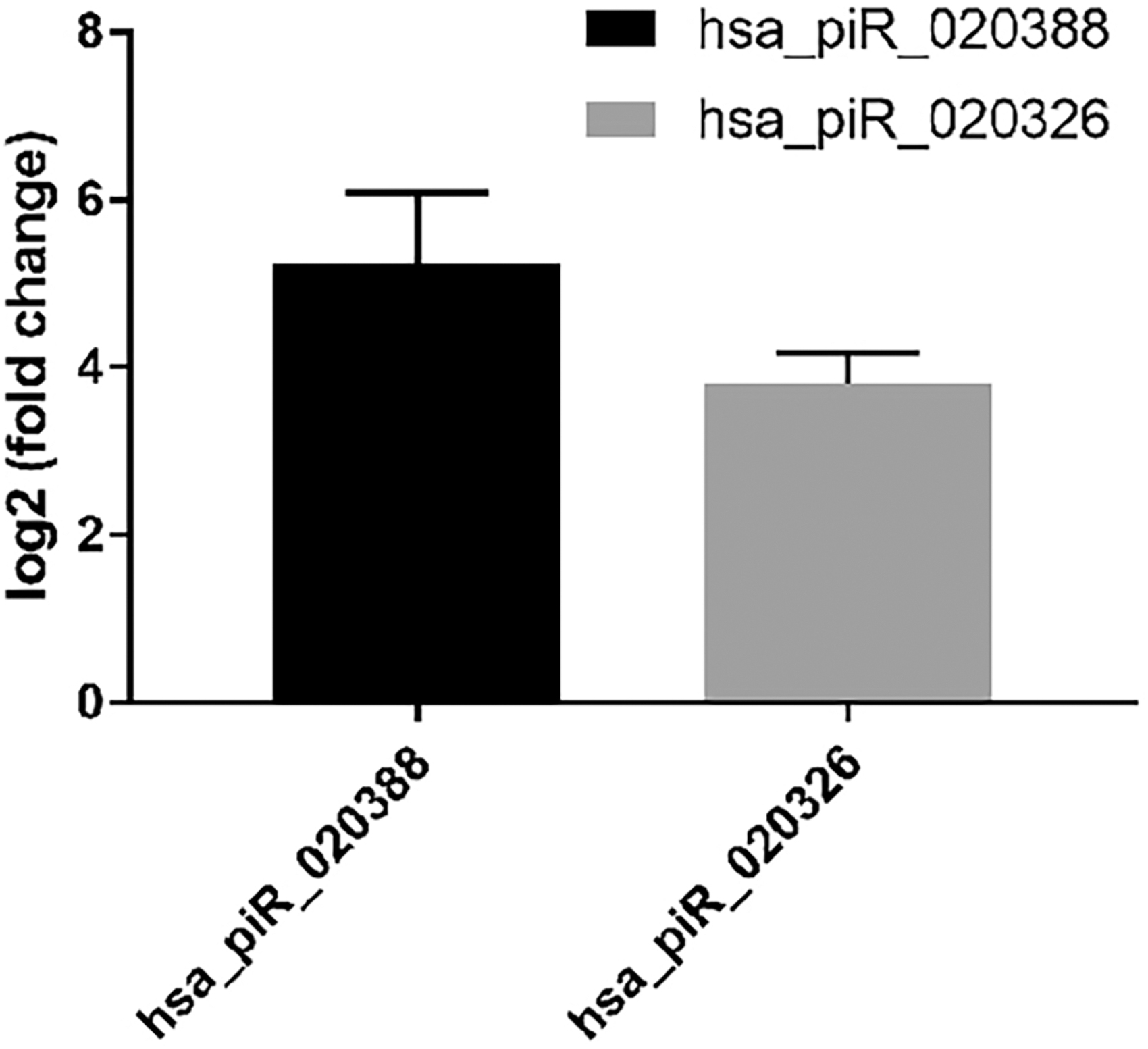
Validation of differentially expressed piRNAs by qPCR. qPCR, quantitative polymerase chain reaction.
Discussion
As an available and promising source for tissue regeneration, SCAP display multiple differentiation potential, with the capacity to give rise to distinct cell lineages, including osteo/odontogenic, adipogenic, and neurogenic lineages. In addition, recent studies have found that SCAP have valuable immunomodulatory capabilities [29 –31]. These biological characteristics of SCAP facilitate their application in tissue regeneration and provide an alternative source of MSCs for tissue engineering. Genes are an important factor in determining the functions of stem cells. With the development of next-generation sequencing technology, we were able to acquire gene expression profiles in MSCs in this study; the functions of MSCs were further predicted by analyzing the differences in gene expression.
Exosomes are extracellular vesicles secreted by most nucleated cells, which are also an important paracrine way for MSCs to exert their biological functions. Intercellular delivery of exosomes has now been demonstrated for a number of different MSCs. When exosomes enter the recipient cells through membrane fusion, they can regulate the phenotypes and functions of the recipient cells by releasing ncRNAs [32]. Many studies have reported that miRNAs and long ncRNAs are contained in exosomes [33,34]. However, few studies have investigated the role of piRNAs in exosomes. This study revealed a distinct piRNA expression pattern in exosomes from SCAP and BMMSCs. To the best of our knowledge, this is the first report on the exosomes in these stem cells that employed an sRNA sequencing strategy.
As the largest class of small ncRNAs, piRNAs can repress the functions of their targets with the assistance of PIWI proteins, which play an important role in regulating tissue development [35]. Our results indicated that the piRNA expression profile of SCAP-exo was statistically different from that of BMMSC-exo, and the proportion of piRNAs was 12.45% in SCAP-exo, which was higher than that in BMMSC-exo. It may be related to the origin of SCAP, which were isolated from early-stage tissues during tooth root development. Among these piRNAs, expression of hsa-piR-011273 and hsa-piR-007832 was the most significantly different. However, these piRNAs have not been previously reported in MSCs, which highlights the importance of our analysis. hsa-piR-020388 and hsa-piR-020326 were randomly selected for qPCR, and the qPCR analyses showed similar results to those detected piRNAs identified with deep sequencing. These data also provide evidence that deep sequencing is a more sensitive and reliable method for identifying differentially expressed piRNAs in MSC-exo.
To further explore the functions of piRNAs in SCAP-exo and BMMSC-exo, GO and KEGG pathways were analyzed to identify the target genes and pathways of the differentially expressed piRNAs. GO enrichment results revealed that these differentially expressed piRNAs targeted genes that function mainly in biological regulation, cellular processes, metabolic processes, single-organism processes, binding, and catalytic activity. These biological functions are closely related to the proliferation, differentiation, and self-renewal ability of MSCs. As one of the mechanisms regulating the function of MSCs, piRNAs are likely to be involved in many key processes.
The KEGG pathway analysis indicated that these differentially expressed piRNAs were associated with multiple carbohydrate, lipid, and amino acid metabolic pathways, which was consistent with previous studies reporting that stem cells play an important role in the body's metabolism [36,37]. In addition, the higher expression of piRNAs in BMMSC-exo was associated with the p53 signaling pathway, which was consistent with a recent report that the p53 signaling pathway may be involved in the osteogenic differentiation of BMMSCs [38]. The KEGG analysis also suggested that the higher expression of piRNAs in BMMSC-exo might be involved in apoptosis. BMMSCs, as an immunoregulatory MSC, are known to regulate apoptosis in immune cells [39,40]. However, the role of piRNAs in apoptosis is unclear.
The upregulated expression of piRNAs in SCAP-exo was associated with the citrate cycle (TCA cycle) and carbon metabolism, the Ras signaling pathway, and the MAPK signaling pathway. The citrate cycle and carbon metabolism are the main metabolic processes used to obtain energy in the body [41,42]. The Ras signaling pathway is a very common cell signaling pathway and is associated with excessive cell proliferation and tumorigenesis [43,44]. hsa-piR-001788, hsa-piR-012895, hsa-piR-014620, hsa-piR-020326, and hsa-piR-020388 were upregulated in SCAP-exo; the KEGG pathway analysis showed that these piRNAs could regulate multiple target genes on the MAPK signaling pathway. The MAPK signaling pathway is known to regulate a variety of important cellular physiological and pathological processes, such as cell growth, differentiation, environmental stress adaptation, and inflammatory response [45]. Several studies have reported that the MAPK signaling pathway is related to the odonto/osteogenic differentiation of MSCs. For example, Wang et al. found that estrogen receptor α regulated the odonto/osteogenic differentiation of SCAP via the MAPK signaling pathway [46].
In summary, we found that the higher expression of piRNAs in BMMSC-exo may be involved in the regulation of apoptosis and osteogenic differentiation. In addition, the higher expression of piRNAs in SCAP-exo was mainly related to metabolism, cell proliferation and differentiation, and other signaling pathways that were closely related to the development of teeth and the formation of bone tissue. These results may increase the understanding of the functional differences between SCAP and BMMSCs and provide novel insight into the molecular mechanisms underlying stem cell therapy. Furthermore, it will be interesting to validate target RNAs and clarify whether these piRNAs actually have a role in cellular functions in the future study.
Conclusions
This study analyzed the expression profiles of piRNAs in SCAP-exo and BMMSC-exo through high-throughput sequencing, and the expression profiles were different. Furthermore, the GO enrichment and KEGG pathway analysis for the differentially expressed piRNAs provided additional information about the biological functions of SCAP and BMMSCs. These results provide a useful resource for further study of the functions and applications of MSCs.
Footnotes
Author Disclosure Statement
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (no. 81771059 and no. 81900963) and the Science and Technology Project of Shenyang (no. F16-102-4-00).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.