
Abstract
The Hippo signaling pathway regulates cell proliferation and organ growth, and its activation is mainly reflected by the phosphorylation levels of Yes-associated protein (YAP). In this study, we show that YAP facilitates embryonic neural stem cell proliferation by elevating their responsiveness to fibroblast growth factor 2 (FGF2), one of the major growth factors for neural stem cells, in vivo as well as in vitro. Western blot and quantitative real-time PCR analyses revealed that expression of the FGF receptors (FGFRs) FGFR1 to FGFR4 were greatly increased by YAP expression upon FGF2 treatment, followed by upregulation of the mitogen-activated protein kinase and protein kinase B signaling pathways. Furthermore, as assessed by quantitative real-time PCR analyses, YAP-induced FGFR expression was found to be TEA domain transcription factor (TEAD)-independent, and transcriptional coactivator with PDZ-binding motif, the other homolog of Yorki in the Drosophila Hippo signaling pathway, was found to possess similar activity to YAP. Finally, adjustment of FGFR signaling activity in the YAP-expressing cells to control levels efficiently offset the cell proliferative effects of YAP, suggesting that the increased proliferation of YAP-expressing neural stem cells was mainly attributable to enhanced FGFR signaling. Our data indicate that YAP plays an important role in neural stem cell regulation by elevating FGFR expression, subsequently leading to enhanced cell proliferation.
Introduction
Neural cell fate is determined when specific temporal and spatial cues are given to the neural stem cells during brain development [1]. In response to those cues, neural stem cells express appropriate transcription factors, and active DNA methylation/demethylation takes place to control transcription [2]. Neural stem cells have two distinguishing features: self-renewal and multipotency [3]. Self-renewal can be further classified into symmetrical division, which results in two daughter neural stem cells, and asymmetrical division, which produces one neural stem cell and another cell with a different fate [3]. Neural stem cells have the capacity to develop into multiple specialized cell types constituting the brain such as neurons, astrocytes, and oligodendrocytes.
The fibroblast growth factor (FGF) family of mitogenic paracrine factors comprises over 20 structurally related members in mammals and regulates the proliferation and differentiation of a wide range of cell types [4 –6]. FGFs exert their effects through four distinct membrane tyrosine kinase FGF receptors (FGFRs), FGFR1 to FGFR4 [7]. Upon FGF binding, the FGFRs dimerize and their cytoplasmic tyrosine kinase domains become active. FGF signaling pathways play critical roles during normal development of both vertebrates and invertebrates, and misregulation leads to diverse developmental defects [8 –11]. Particularly, we previously showed that during the mid-neurogenic period of the mammalian embryonic brain, FGF signaling promotes radial glial character and progenitor proliferation [12].
The Hippo pathway is an evolutionarily conserved signaling system involved in organ size control [13]. Yes-associated protein (YAP) is the core transcriptional coactivator of the mammalian Hippo pathway and its proliferative activity is driven by association with YAP-interacting transcription factors in the nucleus [14,15]. When the Hippo pathway is activated, YAP is phosphorylated on a serine residue and sequestered in the cytoplasm, and when the Hippo pathway is inhibited, unphosphorylated YAP translocates into the nucleus and induces target gene expression that promotes cell proliferation and survival [16]. In terms of brain development, we and other groups reported that YAP activation increases neural stem cell pool expansion by global transcriptional upregulation of cell growth- and proliferation-related genes, and inhibition of YAP activity limits the neural progenitor pool [17 –20]. However, identification and characterization of the exact downstream YAP targets responsible for the enhanced neural stem cell proliferation have remained largely unexplored.
In this study, we show that YAP overexpression in the neural stem cells facilitates activation of the mitogen-activated protein kinase (MAPK) and protein kinase B (PKB, also known as AKT) signaling pathways by increasing FGFR expression. Our study establishes an important functional link between the Hippo and FGFR signaling pathways in the regulation of mammalian neural stem cell proliferation.
Materials and Methods
Plasmid construction
The wild-type and S94A-mutated YAP vectors were purchased from Addgene (plasmid nos. 33091 and 22094) (Cambridge, MA), and subcloned into the MluI site of the retroviral vector MSIG [21]. Transcriptional coactivator with PDZ-binding motif (TAZ) was amplified using the primers mTAZ-F (5′-GGACGCGTGCCACCATGCATAATTCAACAGCTCC-3′) and mTAZ-R (5′-CCACGCGTTCAAGCGTAATCTGGAACATCGTATGGGTACAGCCAGGTTAGAAAGGGCT-3′). The amplified complementary DNA (cDNA) sequences were initially cloned into the pCR2.1 vector (Invitrogen, Carlsbad, CA) and then inserted into the multicloning sites of MSIG.
Retroviral vector production and transduction
The retroviral vector production protocol has been previously described [17]. Briefly, gag-pol (pCA-gag-pol) and env-expressing vector (vesicular stomatitis virus glycoprotein) were transfected with the retroviral construct into HEK293T cells using polyethylenimine (Sigma, St. Louis, MO). After 48 h of transfection, the supernatant was collected and filtered through a 0.45-μm filter. Viral stock concentration was conducted by ultracentrifugation at 100,000 g for 90 min at 4°C in an SW28 rotor (Beckman Coulter, Fullerton, CA). Pellets were resuspended in 100 μL of phosphate-buffered saline (PBS) at 4°C for about 12 h, and aliquoted virus stocks were stored at −80°C.
Neural stem cell preparation and neurosphere assay
Primary neural progenitor cells were prepared from the lateral and medial ganglionic eminences of E14.5 embryos. Dissected brain tissue was minced, washed three times with PBS, and incubated in 0.25% trypsin (Invitrogen) at 37°C for 5 min. DNase and ovomucoid trypsin inhibitor (both from Worthington, Freehold, NJ) were added, and samples were triturated using a fire-polished Pasteur pipette. Cells were washed twice with Dulbecco's modified Eagle's medium (DMEM)/F12 media (Invitrogen), resuspended in PBS, and run through a 40 μm cell strainer (Falcon, Franklin Lakes, NJ). Before neurosphere assays, neural progenitors were transduced with concentrated retroviral vectors. After 48 h, aggregated cells were mechanically dissociated, and 5,000 cells were seeded and incubated in DMEM/F12 media supplemented with B27/N2 (both from Invitrogen) and either FGF2 or epidermal growth factor (EGF) (both from PeproTech, Rocky Hill, NJ). After 7 days of incubation, the number of neurospheres larger than 50 μm in diameter were counted under a light microscope (Eclipse TS100; Nikon, Melville, NY).
Animals and in vivo injection into the embryonic brain
All animal procedures were approved by the Institutional Animal Care and Use Committee of Sungkyunkwan University under the animal protocol No. 201903141. All experiments were conducted in accordance with institutional guidelines and regulations for animal experiments. Timed pregnant CD1 mice (Koatech, Pyeongtaek, Korea) were used for viral injections, and embryos were considered 0.5 days old (embryonic day 0.5, E0.5) when a vaginal plug was detected in the morning. Pregnant mice were anesthetized with pentobarbital sodium (Hanlim Pharm, Gyeonggi, Korea) before the injection, and ultracentrifuge-concentrated viruses containing polybrene (final concentration 80 μg/mL) were injected into the telencephalic ventricle of E13.5 embryos.
Immunofluorescence
Standard immunofluorescence procedures were used for visualization of target gene expression in retroviral vector-injected animals. Briefly, gene-transferred brains at E18.5 were fixed in 4% paraformaldehyde and cryosectioned. Sections were washed in PBS, then blocked for 1 h with PBS containing 10% fetal bovine serum and 0.2% Triton X-100. Samples were then incubated overnight at 4°C with anti-green fluorescent protein (GFP) (A-11122; Invitrogen) and anti-FGFR2 (ab5603; Millipore, Temecula, CA) primary antibodies, washed three times in PBS, and incubated for 1 h at room temperature with Alexa Fluor-conjugated secondary antibodies (A-21206 and A-31570; Invitrogen) diluted in blocking solution. Samples were further counterstained with 4′,6-diamidino-2-phenylindole (D9541; Sigma). Images were acquired using a Zeiss LSM 700 confocal microscope (Oberkochen, Germany).
XTT assay
Primary neural stem cells were initially seeded on 96-well culture plates and cultured with FGF2 or EGF. Cells were treated with 2,3-Bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxyanilide salt (XTT) activator, which was diluted with XTT solution (Biological Industries, Bet-Haemek, Israel). Cells were incubated for 2.5 h at 37°C and absorbance of the samples were measured at 490 nm wavelength, and reference absorbance was measured at 655 nm.
Western blot assay
Using radioimmunoprecipitation assay buffer (Sigma), primary neural stem cells were lysed with a protease and phosphatase inhibitor cocktail (Pierce Biotechnology, Rockford, IL). Equal amounts of protein (20–40 μg, depending on the target protein) were resolved in 8%–10% (w/v) sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride membranes (PALL, Cortland, NY). The membranes were blocked with Tris buffered saline with Tween 20 (TBST) (150 mM NaCl, 10 mM Tris-HCl, 0.1% [v/v] Tween 20, pH 8.0) containing 5% (w/v) skim milk and analyzed with the following primary antibodies: anti-FGFR2 (1:100, sc-122; Santa Cruz Biotechnology, Santa Cruz, CA), anti-p44/42 (1:1,000, 9107; Cell Signaling Technology, Danvers, MA), anti-pp44/42 (1:1,000, 9101; Cell Signaling Technology), anti-AKT1 (1:500, sc-1618; Santa Cruz Biotechnology), anti-p-AKT1 (1:500, 4058S; Cell Signaling Technology), and anti-beta-actin (1:10,000, sc-47778; Santa Cruz Biotechnology). All blots were incubated overnight with a primary antibody at 4°C with horseradish peroxidase-conjugated anti-mouse or anti-rabbit secondary antibodies for 2 h at room temperature. The enhanced chemiluminescence system (Atto, Tokyo, Japan) and X-Omat film (Kodak, Rochester, NY) were used to visualize the protein bands.
Quantitative real-time PCR
Total RNA was prepared from mouse neural progenitor cells using TRIzol reagent (Invitrogen) and cDNAs were synthesized from 500 ng of each RNA sample by using an oligo (dT) primer and Moloney-murine leukemia virus (M-MLV) reverse transcription enzyme (Promega, Madison, WI). Quantitative real-time PCR was performed according to the Smart Cycler System (TaKaRa, Kusatsu, Japan) protocol using the following primers: connective tissue growth factor (CTGF) forward (5′-CTGCCTACCGACTGGAAGAC-3′), CTGF reverse (5′-CATTGGTAACTCGGGTGGAG-3′), FGFR1 forward (5′-ACTCTGCGCTGGTTGAAAAAT-3′), FGFR1 reverse (5′-GGTGGCATAGCGAACCTTGTA-3′), FGFR2 forward (5′-GCCTCTCGAACAGTATTCTCCT-3′), FGFR2 reverse (5′-ACAGGGTTCATAAGGCATGGG-3′), FGFR3 forward (5′-CCGGCTGACACTTGGTAAG-3′), FGFR3 reverse (5′-CTTGTCGATGCCAATAGCTTCT-3′), FGFR4 forward (5′-GCTCGGAGGTAGAGGTCTTGT-3′), FGFR4 reverse (5′-CCACGCTGACTGGTAGGAA-3′), β-actin forward (5′-CAAAAGCCACCCCCACTCCTAAGA-3′), and β-actin reverse (5′-GCCCTGGCTGCCTCAACACCTC-3′).
Results
YAP confers FGF2-dependent growth advantages to embryonic neural stem cells
Typically, either FGF2 or EGF is added to serum-free media to culture neural stem cells derived from embryonic brains. To test the effects of YAP expression on the proliferation of neural stem cells, primary neural stem cells isolated from mouse embryonic day 14.5 (E14.5) brains were infected with retroviral vectors expressing YAP, and incubated in media containing FGF2 or EGF for 3 days. Interestingly, we found that YAP expression enhances neural stem cell growth in the presence of FGF2 (Fig. 1A), but not EGF (Fig. 1B), assessed by a cell proliferation assay using the tetrazolium salt of XTT. These data suggest that YAP-expressing neural stem cells respond distinctly to different growth factors, and that the neural stem cell proliferating activity of YAP is FGF2 dependent.
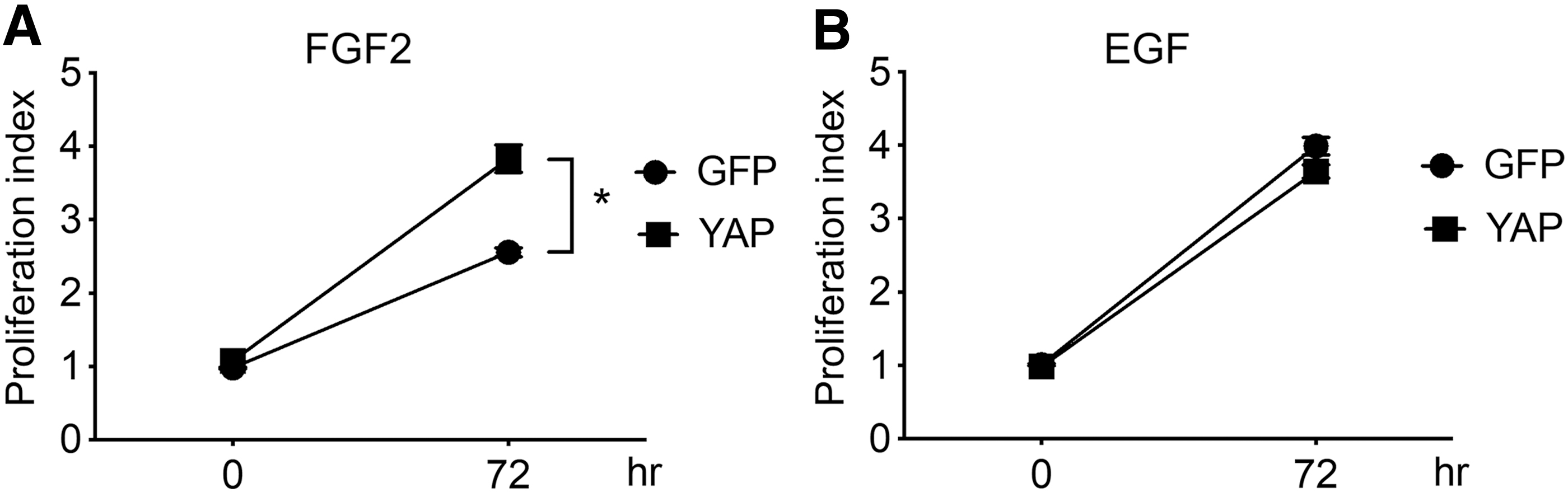
YAP expression enhances FGF2-dependent neural stem cell growth. XTT assays were performed using mouse E14.5 primary neural stem cells transduced with a retroviral vector expressing YAP and then cultured for 3 days in serum-free neural stem cell media containing
YAP increases stemness of neural stem cells in the presence of FGF2, but not EGF
We further tested the effects of YAP expression on neural stem cell properties using the neurosphere assay. The neurosphere assay is a cell survival assay based on the principle that only neural stem cells form a colony (neurosphere) in the defined stem cell culture media [22,23]. As shown in Fig. 2A–C, YAP expression resulted in significant increase in both neurosphere formation frequency and size suggesting that YAP enhances the self-renewal capability and proliferative activity of neural stem cells, respectively, whereas YAP expression did not substantially affect EGF-dependent neurosphere formation. These data indicate that YAP expression enhances neural stem cell properties when FGF2, not EGF, is used as a growth factor.
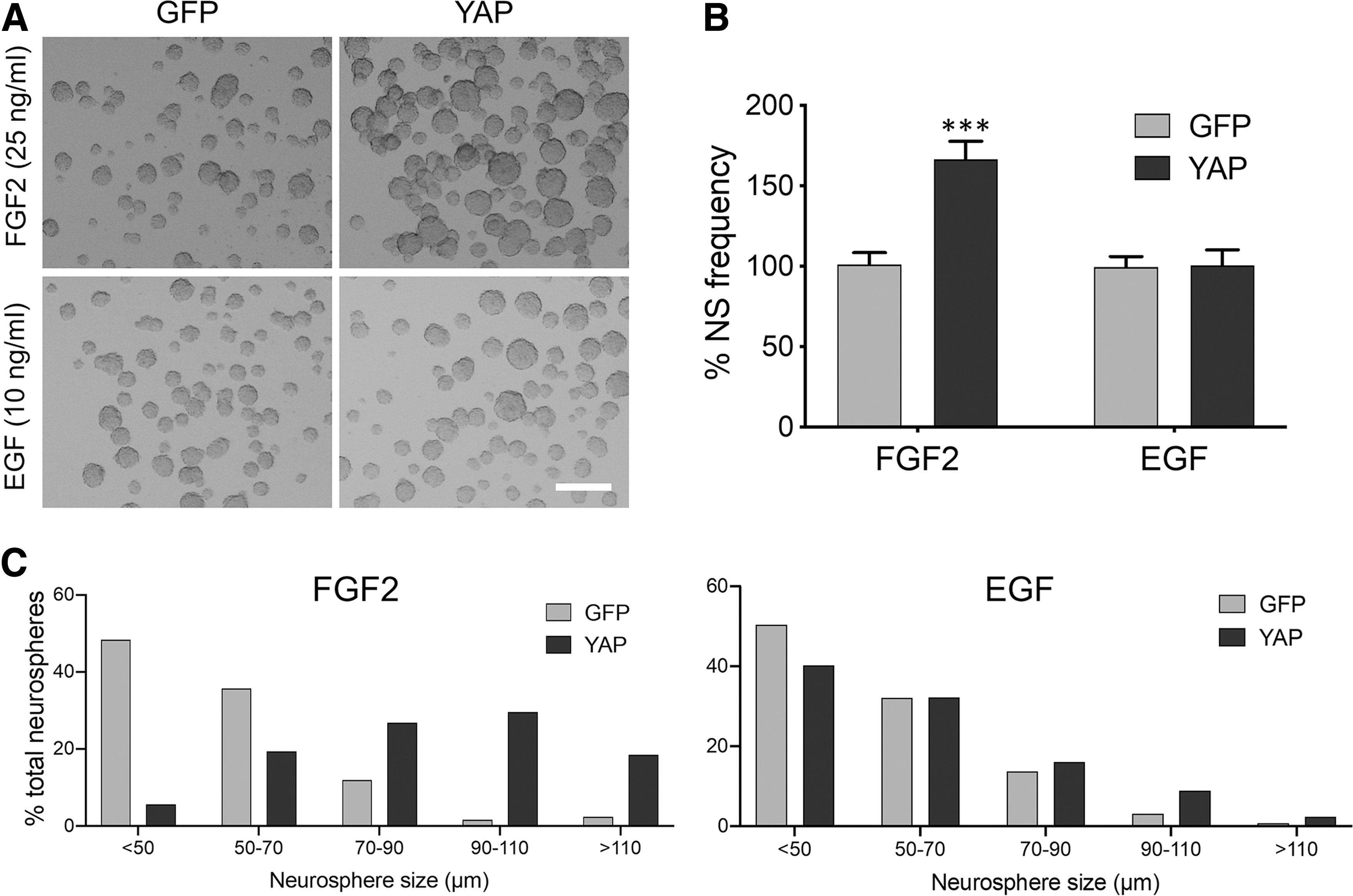
YAP-enhanced neural stem cell properties are FGF2 dependent, but not EGF dependent. Mouse E14.5 primary neural stem cells were transduced with a retroviral vector expressing YAP and then incubated in serum-free media containing 25 ng/mL FGF2 or 10 ng/mL EGF. At 7 days post-transduction, neurospheres were photographed as in
YAP enhances FGFR expression in a TEA domain transcription factor 2-independent manner followed by downstream signal activation
Next, we investigated which cellular components were responsible for the YAP-enhanced neural stem cell proliferation. First, we assessed the activation of MAPK signaling cascade, which is known to be one of the major pathways stimulated by FGF2 [5]. As shown in Fig. 3A and B, YAP expression in neural stem cells in vitro resulted in higher levels of p44/42 MAPK (ERK1/2) phosphorylation than control GFP expression upon FGF2 treatment. In addition to MAPK, AKT activation was also more efficiently induced in response to FGF2 treatment in the YAP-expressing cells (Fig. 3A, B). Enhanced neural stem cell proliferation by YAP overexpression in vitro was also confirmed by immunostaining using anti-Ki67 antibody (Fig. 3C, D). However, YAP overexpression did not affect the stemness of neural stem cells, as assessed by immunostaining with anti-SRY (sex determining region Y)-box 2 (SOX2) and anti-Nestin antibodies (Supplementary Fig. S1). These data suggest that the cellular factor, which contributes to the YAP-enhanced neural stem cell proliferation is upstream of the branching point of the MAPK and AKT signaling pathways, proximal to the cell membrane. To test this hypothesis, we measured the levels of FGFR2 because it is expressed in the ventricular zone during brain development [24] and found that FGFR2 proteins were significantly increased in the YAP-expressing cells in vitro (Fig. 3A, B). Moreover, in an in vivo context, cells expressing a constitutively active (CA) form of YAP in the developing brain formed clusters, presumably reflecting the highly proliferative activity of CA YAP, and expressed FGFR2 (Fig. 3E). quantitative real-time polymerase chain reaction (qRT-PCR) analysis further revealed that gene expression of FGFR1, FGFR3, and FGFR4 was also increased by YAP expression in vitro (Fig. 3F).
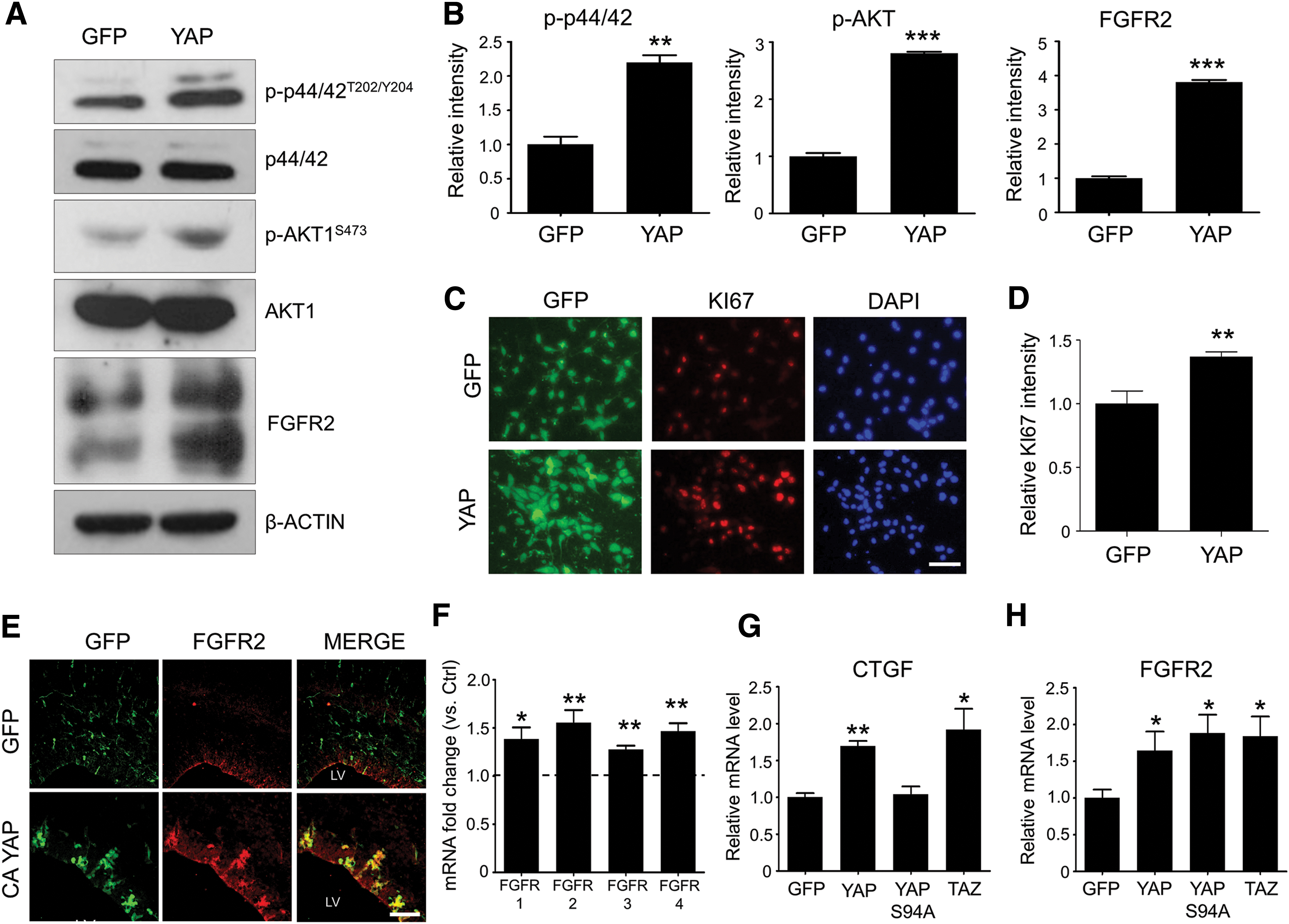
YAP expression facilitates the FGF signaling pathway by inducing FGFR expression.
Among the YAP-interacting cotranscriptional factors, we suspected that YAP-increased FGFR expression was TEA domain transcription factor dependent because, like YAP, TEAD2 is abundantly expressed in the developing brain (
YAP-increased neural stem cell proliferation reverted to control levels upon inhibition of the FGFR signaling pathway
Finally, to assess the level of contribution of increased FGFR expression to YAP-enhanced neural stem cell proliferation, we utilized the FGFR pathway inhibitor, AZD4547. Since strong inhibition of FGFR signaling would completely inhibit normal neural stem cell growth, we tried to reduce only as much FGFR signaling activity as YAP expression additionally increased. For this, we treated YAP-transduced neural stem cells with several concentrations of AZD4547 for 48 h, and determined that 10 nM was the concentration at which p44/42 MAPK phosphorylation was lowered to control levels by western blot analysis (Fig. 4A, B). As expected, YAP-transduced neural stem cells incubated with 10 nM AZD4547 grew similarly to control cells until 72 h after transduction (Fig. 4C). Taken together, these data indicate that the increased proliferative effect of YAP expression on the neural stem cells is mainly attributable to YAP-enhanced FGFR signaling pathway activation.
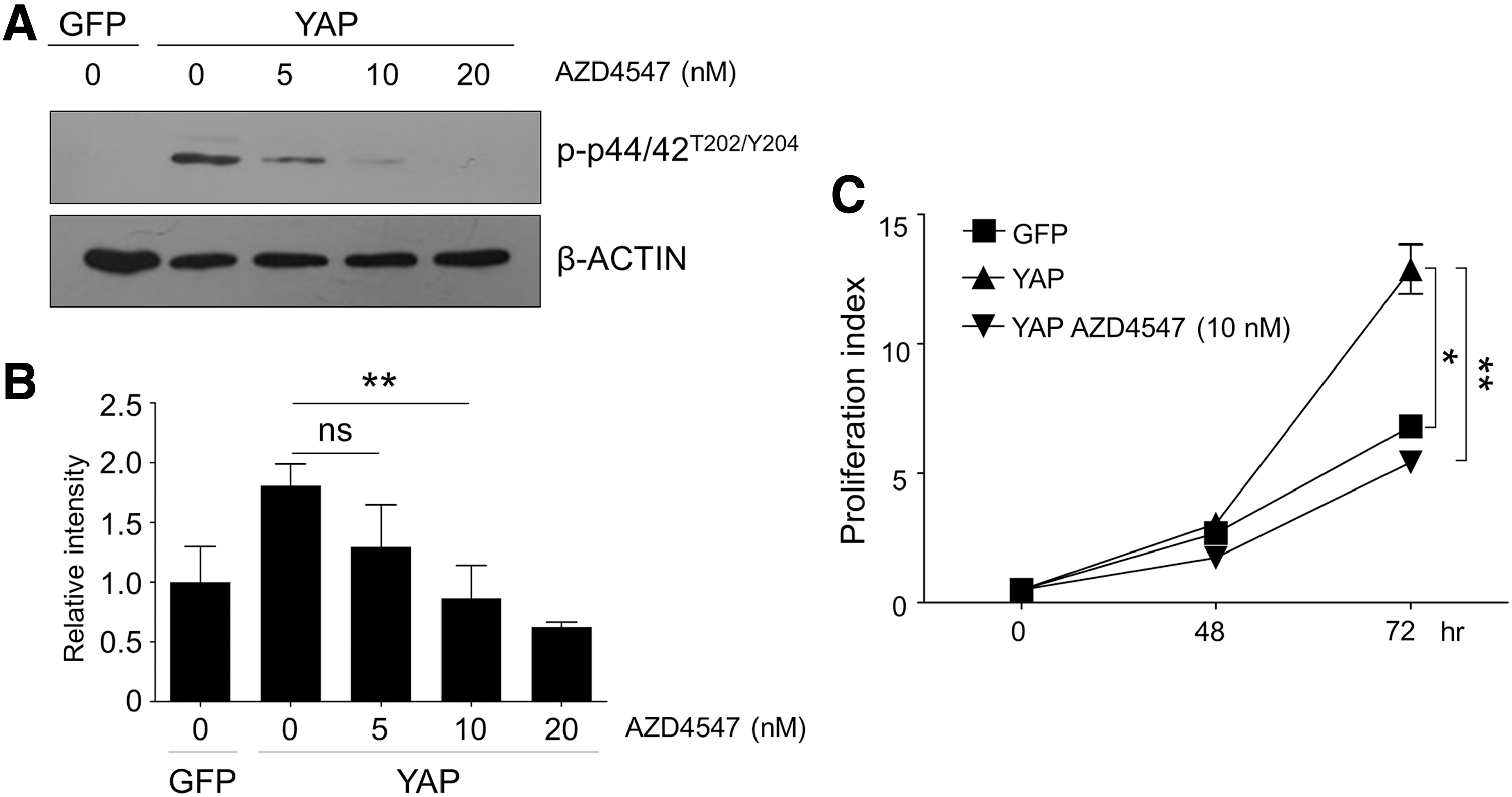
YAP-increased neural stem cell proliferation was offset by the inhibition of FGFR signaling.
Discussion
There has been increasing interest in the roles of YAP in neural stem cell regulation and brain development. In this study, we show that YAP expression benefits neural stem cell proliferation when the cells are stimulated by FGF2, but not EGF, and that elevated FGFR levels contribute to the YAP-upregulated response to FGF2. Our group previously reported that YAP expression enhances neural stem cell properties based on observed phenotypic changes [17]. Here, we provide deeper insight into the mechanistic roles of YAP by establishing a functional link between YAP and the FGFR signaling pathways in the regulation of neural stem cells at the embryonic stages.
Recently, Rizvi et al. [29] reported an association between FGFR and YAP expression. However, several differences exist between that study and the present one. First, they measured FGFR expression in cholangiocarcinoma cells, not neural stem cells. Second, although TBX5 was identified as a key YAP-binding cotranscriptional factor responsible for the YAP-dependent FGFR expression in the cholangiocarcinoma cells, it is not likely that TBX5 also plays a similar role in the neural stem cells because it is not expressed in the central nervous system during the embryonic mid-neurogenic period (
TEAD2 is strongly expressed in the ventricular zone (VZ) in the developing brain, and consistently, we previously reported that YAP-increased self-renewal ability of embryonic neural stem cells is TEAD dependent. Therefore, we suspected that TEAD would be the transcriptional binding partner of YAP for FGFR expression. But surprisingly, the YAP S94A mutant lacking TEAD binding ability still exerted FGFR expression-inducing effects at wild-type levels. These results suggest that another transcriptional partner(s) besides TEAD2 and TBX5, as mentioned above, plays a role in YAP-induced FGFR expression. More importantly, the present study also implies that YAP-enhanced self-renewal activity and YAP-increased neural stem cell proliferation by augmenting the FGFR signaling pathway appear to be distinctly regulated by different YAP cotranscriptional factors.
Because protein structure and functions of TAZ are closely related to YAP [27,28], we expected that it would also exhibit the FGFR-increasing activity. As expected, TAZ expression enhanced FGFR expression in the neural stem cells. Thus, given the YAP-increased FGFR expression in cholangiocarcinoma cells [26], TAZ is also highly likely to induce FGFR expression in cancer cells. Further studies using cancer cell models in which TAZ expression is detected should address this hypothesis.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This research was supported by the National Research Foundation of Korea funded by the Ministry of Science and ICT (grant nos. 2018R1A2B2001076 and 2019M3A9H1103702).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.