
Abstract
Mechanically stretched skeletal muscle undergoes dramatic shifts in structure, mass, and function. In vitro tensile strain models have demonstrated that myogenic progenitor cells, including satellite cells and myoblasts, are highly mechanosensitive cells, and respond to mechanical strain in a wide variety of aspects. However, the experimental results from different researchers and laboratories are not always in support of each other. Moreover, some specific molecules or signaling pathways were reported to play distinct roles in stretched myogenic cells, according to the statements of different studies. The purpose of this review is to integrate the researches conducting in vitro culture of satellite cells or myoblasts and exploring their mechanoresponses using in vitro stretching apparatus. These responses will be categorized into several groups, such as activation, proliferation, myogenic differentiation, cellular damage or apoptosis, properties of plasma membrane, transdifferentiation, reorientation, etc. In addition, detailed experimental designs like culturing conditions and straining regimens will be displayed and compared, to interpret some contradictory statements in different studies. Furthermore, the currently known interconnections among some mechanosensitive pathways will be pictured to give a better understanding about the complex regulations of myogenic cell responses to mechanical stretch. Hopefully, by summarizing the published studies about mechanoresponses of myogenic progenitor cells, future directions, and perspectives would be made clearer to researchers in this field.
Introduction
Skeletal muscle is a mechanically sensitive tissue, with many aspects (structure, mass, function, etc.) being affected by mechanical forces [1]. During the past decades, intensive in vivo studies have been conducted to investigate the effects of mechanical stimulation on skeletal muscles, using animal models like hindlimb soleus muscle stretching or contraction. Moreover, a substantial number of ex vivo studies, through isolating and stretching single muscle fiber adhering to the flexible culture plates, explored its mechanosensitive properties. Based on these valuable experimental data, it is now generally accepted that mechanical stimuli are an indispensable factor contributing to skeletal muscle homeostasis (see review refs. [2 –4]).
Even though numerous researchers reported that these mature muscular structures (myofibers, myofibrils, or even fusing myotubes) are responsive to mechanical stimuli, myogenic resident cells are also inevitably exposed to mechanical forces during daily activity of muscles, giving rise to a new field of research: what part do they play in the stretch-induced physiological and pathological changes of muscles?
Satellite cells are considered the primary stem cells in adult skeletal muscle, locating between the sarcolemma and the basal lamina of the myofiber [5]. Upon external stimuli, they are activated from the quiescent state, and experience both symmetric and asymmetric divisions, guaranteeing the maintenance of the stem cell pool and the production of committed myogenic progenitor cells, also called myoblasts [6]. These myoblasts continue to proliferate, migrate, differentiate, and fuse to form terminally differentiated myotubes, myofibrils, and myofibers. Because of their close location around muscle fibers, it is tempting to assume that both satellite cells and myoblasts are subject to mechanical stretching during physical exercise of skeletal muscle. Thus, stretch-induced behaviors of these cells presumably affect the physiological and pathological changes of skeletal muscle under mechanical stimuli.
Excitingly, with the help of in vitro cell stretching devices, accumulating evidences suggested that satellite cells and myoblasts do react in their unique fashions when they were mechanically deformed during in vitro stretching. Depending on various experimental designs, they were reported to be activated, proliferate, differentiate, or damaged, during stretching stimuli. In addition, stretch also affected their functions, arrangement, multipotency, etc. In this study, we integrated these in vitro studies and provided an overview of the influences of external mechanical stretching on satellite cells and myoblasts. Furthermore, we speculated the interconnections among different studies exploring the same pathways, and interpreted studies with contradictory results. Finally, future research directions in mechanoresponses of skeletal myoblasts will be addressed.
Stretch-Induced Satellite Cell Activation
In skeletal muscle, satellite cells are mitotically quiescent under normal condition, but are activated from quiescent state to re-enter cell cycle. In vitro culture of satellite cells has been demonstrated to be activated by mechanical stretch. A relatively clear mechanism has already been proposed regarding stretch-induced satellite cell activation, which will be discussed below.
Hepatocyte growth factor binding to c-mesenchymal–epithelial transition receptor
Hepatocyte growth factor (HGF) is present in the extracellular matrix (ECM) of uninjured skeletal muscle fibers, and is expressed and secreted by the resident myotubes and satellite cells [7]. So far, HGF is the only growth factor being able to activate quiescent satellite cells when exogenously added in culture medium [8]. Tatsumi et al. [9] first discovered that HGF was released from ECM into medium when satellite cells were stretched. The release of HGF in response to stretch is dependent on pH, with the maximum HGF being released in stretch-conditioned medium at pH 7.2 and diminished amounts at pH 7.0 and 7.4 [10]. Furthermore, stretch-released HGF-mediated activation of stretched satellite cells [9]. Interestingly, unlike other tissues, skeletal muscle possesses the active form of HGF that does not require the proteolytic activation of pro-HGF in response to stretch [11].
The specific receptor for HGF in satellite cells is c-mesenchymal–epithelial transition (c-Met) factor, which is present in quiescent satellite cells, thus available for HGF binding [9]. Notably, HGF constantly binds to c-Met over a broader pH range from 6.9 to 7.8, in contrast to the pH-sensitive release of HGF mentioned above [10]. By isolating satellite cells from different skeletal muscle tissues, Tatsumi et al. [12] found that satellite cells from lower hind limb muscles displayed the highest activation in response to stretching stimuli, which could be attributed to more HGF release and higher binding sensitivity of HGF to c-Met. Although these studies reflected the involvement of c-Met in stretch-induced satellite cell activation, the direct evidence showing this viewpoint is still lacking. Future study employing c-Met(−/−) satellite cells might fully interpret this point.
Nitric oxide synthase-mediated cleavage of matrix metalloproteinase 2
Matrix metalloproteinase (MMPs) are a large family of zinc-dependent endopeptidases that catalyze specific ECM constituent. Yamada et al. [13] had confirmed the involvement of MMP2 in stretch-induced rat satellite cell activation, as inhibition of MMP2 by tissue inhibitors of MMPs 1 completely blocked stretch-induced activation. They further demonstrated that MMP2 mediated HGF release in response to stretch [13,14]. To date, MMP2, 3, 7, and 9 have been found in skeletal muscle [8]. In addition, MMP9 activity and expression were increased in C2C12 myoblasts during cyclic stretching [15]. Thus, it needs to be clarified whether these MMPs besides MMP2 also participate in mechanical stretch-activated satellite cells.
To catalyze HGF release, proMMP2 (72 kDa) must be converted into active MMP2 (52 kDa), which relies on nitric oxide synthase (NOS)-mediated nitric oxide (NO) production in stretched satellite cell culture [14]. Skeletal muscles mainly express neuronal NOS (nNOS or NOS-I) and some endothelial NOS (NOS-III), but not inducible NOS (NOS-II) [10]. Inhibition of nNOS abrogated proMMP2 conversion, HGF release, and satellite cell activation during mechanical stretching [14,16]. Collectively, these studies established the nNOS-NO-MMP2-HGF axis that mediated stretch-induced activation of satellite cells.
Involvement of calcium/calmodulin
Hara et al. [17] proposed that mechanosensitive (MS) cation channel and the long-lasting-type voltage-gated calcium ion (L-VGC) channel mediate the influx of extracellular calcium ions in satellite cell cultures in response to stretch. Moreover, they confirmed that stretch-induced calcium influx initiated the HGF release and satellite cell activation through calmodulin pathway [18]. The transient receptor potential canonical channel 1 represents an essential component of MS channel in C2C12 myoblasts [19]. Another potential candidate for MS channel in myoblasts is Na cation pump, which is activated by alpha2 subunit of Na-K-ATPase that translocated from endosomes to the plasma membrane (PM) in primary rat myoblasts during mechanical strain [20].
Thus, according to the model of Hara et al., the cations streamed into cells through these MS channels, and induced local depolarization of the cell membrane, which promoted gating of adjacent L-VGC channels and allowed sufficient calcium influx [17].
Inhibition of inflammatory autoimmune
Interestingly, the Calmodulin-nNOS-MMP2-HGF-cMet pathway activated by mechanical stretch in myoblasts not only stimulates their entry into cell cycle, but also concomitantly inhibits the expressions of potential autoantigens. This was demonstrated by the elevated expressions of autoantigens (Mi-2, HRS, DNA-PKcs, U1-70) and toll-like receptor 3 by inhibition of Calmodulin-nNOS-MMP2-HGF-cMet pathway in C2C12 myoblasts under 10% cyclic radial stretch [21]. Thus, the authors stated that appropriate mechanical stimuli are beneficial in preventing the inflammatory myopathies. This is of particular importance in chronic inflammatory situations, since regenerating muscle cells are considered to be the main source of autoantigens and contribute to the progression of inflammatory myopathies during inappropriate exercise training [21].
Effect of Stretch on Skeletal Myoblast Proliferation
Up to date, most studies conducting in vitro stretching of myoblast cultures agreed that mechanical stretch is a positive factor in regulating myoblast proliferation. A detailed summary of these researches is present in Table 1. In this study, we listed some frequently implicated molecules by different studies, suggesting their role in stretch-regulated myoblast proliferation (Fig. 1).
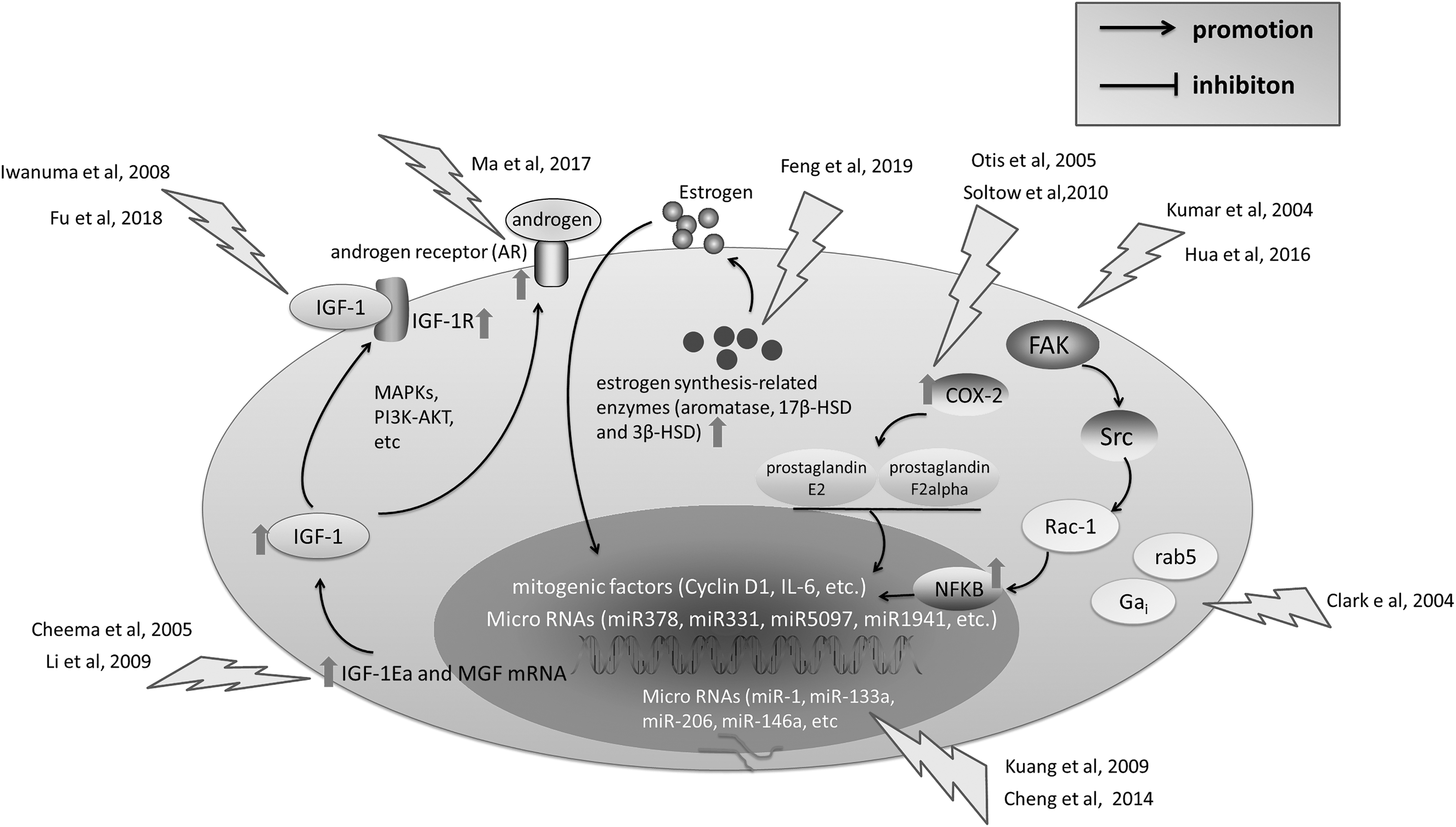
Major molecules and pathways involving in the proliferation of myoblast during stretching stimuli. Several pathways mediated the stretch-induced myoblast proliferation, such as FAK-Src-Rac-NF-κB, COX2, IGF-1, and some hormone-related pathways like androgen and estrogen. In addition to these, some micro-RNAs were also shown to positively regulate myoblast proliferation. It remains elusive whether there were more than one of these pathways being activated at the same time during stretch-mediated myoblast proliferation and what type of stretch may specifically activate one particular pathway. FAK, focal adhesion kinase; COX2, cyclooxygenase 2; IGF-1, insulin-like growth factor-1; NF-κB, nuclear factor-kappa B.
Summary of Mechanical Stretch on the Proliferation of Myoblasts
a-MEM, alpha modified minimum essential medium; DMEM, Dulbecco's modified Eagle's medium; NA, none available.
Nuclear factor-kappa B
Kumar et al. [22] manifested the DNA-binding activity of nuclear factor-kappa B (NF-κB) was upregulated in mechanically stimulated C2C12 myoblasts in a time-dependent manner. Furthermore, they confirmed that NF-κB was essential in stretch-induced proliferation of C2C12, since transfecting the IκB (inhibitor of NF-κB) completely restrained the elevated proliferation during 1 h stretch [22]. On the other hand, the indispensable role of NF-κB in expansion of C2C12 myoblasts during longer duration of stretch (2 h/day for 2 and 4 days) was testified by using NF-κB-specific inhibitors, Pyrrolidine dithiocarbamate and BAY11-7082 [23,24].
Regarding the upstream events contributing to stretch-activated NF-κB, Kumar et al. found that focal adhesion localized protein tyrosine kinases, such as focal adhesion kinase (FAK) and Src, led to the activation of Rac1 GTPase, which directly activated NF-κB [22]. In addition, other GTPase such as rab5 and Gai , were also activated in C2C12 myoblasts by rapid stretching stimuli, in both strain magnitude and strain rate-dependent manners, presenting themselves as potential activators of NF-κB [25]. In addition to these, Ji et al. [26] corroborated that p38 mitogen-activated protein kinase (MAPK) upregulated NF-κB transcriptional activity in C2C12 myoblasts under stretching, partially through phosphorylation of p65 subunit.
How does the activation of NF-κB in response to mechanical stretch lead to the proliferation of C2C12 myoblasts? It has been shown that cyclin D1 and interleukin-6, two well-known mitogenic factors, contain NF-κB-binding sites in their promotor regions and showed increased expressions in stretched C2C12s [22]. In addition, some MicroRNAs, such as miR378, miR331, miR5097, and miR1941, were reported to be regulated by NF-κB, being candidates in modulating stretch-induced C2C12 myoblast proliferation [24].
Cyclooxygenase 2
Cyclooxygenase 2 (COX2) is an immediate/early gene that is induced during cell injury and inflammation. Its specific inhibitor NS-398 suppressed uniaxial stretch-induced C2C12 proliferation, implying COX2 is involved in propelling C2C12 myoblast proliferation in response to stretching [23]. Notably, even though COX2 has NF-κB-binding sites in its promoter region, COX2 was not a downstream factor of NF-κB [23]. Another research reported the implication of COX2 in radial stretch-induced C2C12 proliferation by treating C2C12 cells with COX2 inhibitor SC-236 or by using COX2−/− myoblasts [27]. Moreover, they proposed prostaglandin E2 and F2alpha as downstream effectors of COX2 pathway, since supplementing stretched COX2−/− myoblasts with prostaglandin E2 or F2alpha partially rescued the inhibited proliferation [27].
Insulin-like growth factor-1
Insulin-like growth factor-1 (IGF-1) is a growth factor that functions by binding to its receptor IGF-1R. Iwanuma et al. [28] found that IGF-1 mRNA and protein expressions were elevated in C2C12 cells during the early stage (12 and 24 h) of mechanical stretching, and decreased in late stage (36 and 48 h). Furthermore, C2C12 proliferation rate was highly correlated with IGF-1 level in their study, suggesting a potential function of IGF-1 in promoting C2C12 expansion during strain. Consistently, the study by Li et al. [29] also observed that cell number of primary rat myoblasts commensurate with IGF-1 mRNA level when stretched for 0.5, 1, 2, 4, 8, and 24 h. Importantly, two isoforms of IGF-1 (IGF-IEa and mechano growth factor, also known as MGF) displayed distinct mechanical responses, and only MGF is responsible for the stretch-elevated myoblast proliferation [29,30].
On the other hand, the downstream events of IGF-1 signaling were reported to be PI3K-AKT and MAPKs (p38, ERK), the inhibition of which blocked stretch-induced C2C12 and L6 myoblast proliferation [31,32].
Hormone-related genes
The anabolic action of testosterone is mediated by intracellular androgen receptor (AR). Ma et al. [31] showed that AR expression was elevated by 15% stretch, accompanied with increased C2C12 proliferation, which was suppressed by AR inhibitor. On the other hand, estrogen level is also associated with muscle function and metabolic disorders, and C2C12 myoblast expressed raised levels of estrogen synthesis-related enzymes, including aromatase, 17β-HSD, and 3β-HSD, in response to mechanical stretching [33]. In addition, the cell proliferation was positively correlated with the level of estrogen [33]. Whether these estrogen synthesis-related enzymes play crucial roles in prompting expansion of stretched myoblasts remains to be determined by the use of their specific inhibitors.
Micro RNAs
Three muscle-specific Micro RNAs (miR-1, miR-133a, and miR-206) are essential for proliferation and differentiation of skeletal myoblasts [34,35]. Moreover, when human primary skeletal myoblasts were mechanically stimulated for 1 h, the expressions of these miRNAs were increased in 8% horse serum culture while decreased in 2% horse serum culture [36]. Another study found that miR-146a was upregulated in C2C12 cells upon 48 h cyclic stretch, accompanied by increased proliferation and repressed level of Numb that could facilitate myogenic differentiation. Inhibition of miR-146a rescued the expression of Numb at the cost of compromised proliferation [37].
In addition to these miRNAs, other miRNAs, such as miR500, miR1934, miR378, miR31, miR331, miR5097, and miR1941, were also reported to be regulated by mechanical stretching of C2C12 myoblasts, as we mentioned above to be downstream factors of NF-κB [24].
Protein translation initiation and elongation
It is well known that cell proliferation is accompanied with activation of anabolic pathways and increased protein synthesis. Thus, mechanical stimuli play a major role in the regulation of skeletal muscle mass by stimulating both myoblast proliferation and protein synthesis. Nakai et al. [38] demonstrated that static or cyclic uniaxial stretch activated p70 S6 kinase (p70S6K) and eukaryotic elongation factor 2 (eEF2), which are the makers for protein translation initiation and peptide chain elongation, respectively.
Furthermore, mammalian target of rapamycin (mTOR) was shown to act upstream of p70S6K and eEF2 activation during stretching of myoblasts. Inhibition of mTOR not only prohibited translation initiation, but also induced apoptosis in myoblasts under mechanical tension [38,39]. One mechanism considering the activation of mTOR in stretched myoblasts is the phosphorylation of some tyrosine kinases under stretching stimuli [38]. Another potential candidate regulating mTOR in myoblasts exposed to mechanical stretch is regulated in development and DNA damage responses 2 (REDD2), which is enriched in myoblasts and negatively regulated mTOR signaling by associating with 14-3-3 protein and releasing tuberous sclerosis complex 2 from the inhibition of 14-3-3 [40].
Repressed myoblast proliferation by intensive stretch
Notably, the reported mitogenic effect of mechanical stretch on myoblast is largely because of the optimized experimental conditions (magnitude, frequency, duration). For example, 0.5 Hz cyclic stretch at the magnitude of 15% propelled myoblast proliferation, whereas 20% magnitude stretch impeded it [31]. Similarly, another study applied the static stretch at the magnitude of 1%, 4%, and 168%, and demonstrated that myoblast proliferation was provoked by 1%, 4%, but hindered by 168% stretch [41]. Hua et al. found that 10% cyclic stretch at 0.125 Hz rendered proliferation, but 0.5 Hz cyclic stretch obstructed it [24]. With regard to stretching duration, Feng et al. [42] showed that C2C12 proliferated most rapidly at 4 h of cyclic stretch, whereas the proliferation rate started to attenuate after longer exposure (6 and 8 h) to stretch.
Biphasic Effect of Stretch on Skeletal Myoblast Differentiation
Myoblast differentiation is initiated when myoblasts irreversibly withdraw from the cell cycle and start to express muscle-specific genes, some of which will be discussed in this study because of their mechanosensitive properties. In addition, it should be noted that mechanical strain has both positive and negative effects on myoblast differentiation, as will be displayed in Table 2 and Fig. 2.
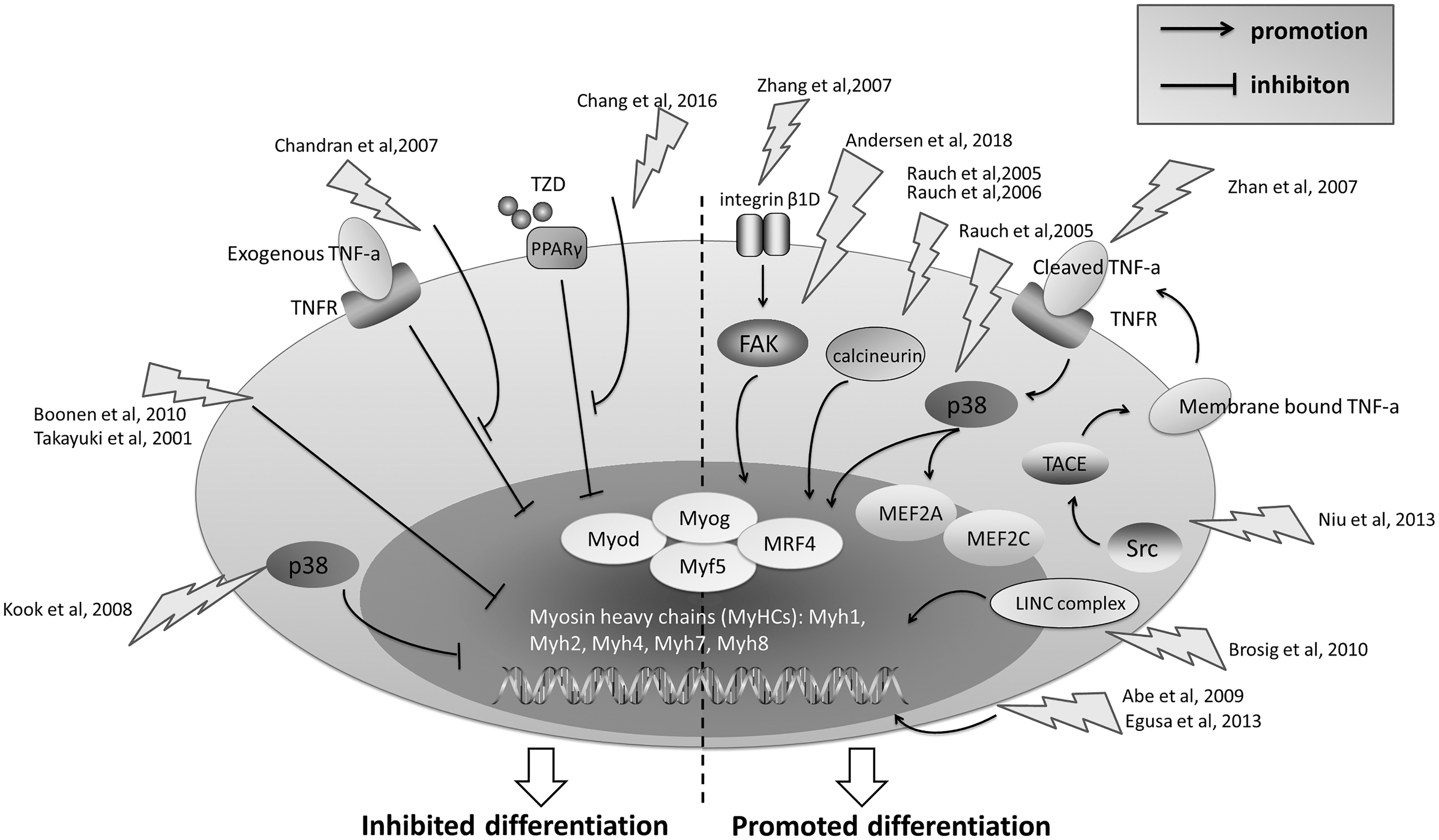
Major molecules and pathways involving in the myogenic differentiation of myoblast during stretching stimuli. Several pathways were reported to promote myoblast differentiation under various stretching stimuli, such as integrin β1D-FAK, calcineurin, p38 MAPK, Src-TACE-TNFα. The activation of MRF and MEF2 transcription factors might act downstream of these pathways and directly induce transcriptions of muscle-specific genes. On the other hand, p38 MAPK, and TNFα pathways were also reported to inhibit myoblast differentiation under some specific mechanical conditions. Moreover, mechanical stretch could antagonize the effect of inhibitors of myogenesis such as PPAR agonist TAD and exogenous TNFα. Altogether, mechanical stretch has dual effects (promotion or suppression) on myoblast differentiation. TNFα, tumor necrosis factor α; MRF, myogenic regulatory factor; MEF2, myocyte enhancer-binding factor 2; TACE, TNFα-converting enzyme; MAPK, mitogen-activated protein kinase; PPAR, peroxisome proliferator-activated receptor.
Summary of Mechanical Stretch on the Differentiation of Myoblasts
a-SMA, α-smooth muscle actin; DM, differentiation medium; GM, growth medium; MHC, myosin heavy chain; MLP, muscle LIM protein; MNF-a, myocyte muclear factor alpha; MRF, myogenic regulatory factor; MyHC, myosin heavy chain; TPM1, tropomyosin.
Myogenic regulatory factor transcription factors
Myogenic regulatory factors (MRFs), including Myod, Myog, MRF4, and Myf5, induce transcriptions of muscle-specific target genes by binding to the E box in the promoters of these genes [43]. MRFs were mechanosensitive genes in myoblasts, and some studies manifested increased levels of MRFs, such as Myog [44 –50], Myod [44,46,48,50], Myf5 [49,50], and MRF4 [50], in C2C12 cells under stretching stimulation, accompanied by accelerated myogenic differentiation. On the other hand, mechanical stretch was also reported to repress the expressions of these MRFs, such as Myog [27,37,51 –54] and Myod [51 –55], which was consistent with impaired myogenic differentiation of C2C12 cells. In addition, Dugan et al. [56] reported that both Myog and Myod were unaffected by mechanical stretch, but maturation of myotubes was perturbed.
The contradictory results of these published literatures could not simply be attributed to stretching parameters alone (static/cyclic, radial/uniaxial, continuous/intermittent, long/short duration). Instead, we insisted that the culturing status of myoblasts, such as constituent of culturing medium and stages of myogenic differentiation, have a synthetic effect on myoblast differentiation. In addition, different sources of myoblasts (rat, mouse, calf) used in these studies may also account for the various responses to mechanical stretch. In spite of these, the mechanisms of the multifaceted effects of stretch on the expressions of these MRFs remain to be elucidated.
Myocyte enhancer-binding factor 2 transcription factors
Myocyte enhancer-binding factor 2 (MEF2) proteins, such as MEF2A, 2B, 2C, and 2D, function as accessory regulators of muscle gene expression, through physically binding and cooperating with MRFs [57]. Similar to MRFs, mechanical stretch also positively regulated MEF2A [46,58] and MEF2C [45] expressions in C2C12 myoblasts. Particularly, MEF2A phosphorylation and subsequent nuclear translocation is critical for MEF2A to exert its function [58]. Still, another study found that MEF2 phosphorylation was unchanged during mechanical stretching [44]. Based on these results, it will be interesting to systematically investigate how does a specific type of stretch affect the expressions of both MRFs and MEF2 proteins, as well as their interactions in myoblasts, as this will fully interpret the myogenic differentiation by mechanical stretch.
Myosin heavy chains
Myosin heavy chains (MyHCs), encoded by Myh genes, are muscle contraction proteins that consist of many isoforms [59]. The changed expressions of these isoforms are indicative of muscle development, metabolism, and function. Some studies displayed either increased or decreased mRNA levels of Myh1 (MyHC-2D or MyHC-2X) [46,53,60,61], Myh2 (MyHC-2A) [46,50,53,60,61], Myh4 (MyHC-2B) [46,53,60,61], Myh7 (MyHC-β or MyHC-1) [60], Myh8 (MyHC-neo) [53], in myoblasts under stretching stimuli. Notably, the changed levels of these Myh genes concur with the biphasic effect of stretch on myoblast differentiation. On the other hand, other studies demonstrated stretch could also induce MyHC protein expressions, accompanied by enhanced myoblast differentiation [44,45].
The upstream pathways regulating the Myh genes expressions in myoblasts under mechanical environment were p38 kinase and calcineurin phosphatase [44,58]. Since aforementioned MRFs and MEF2s are direct transcriptional regulators of these Myh genes, it is reasonable to deduce stretching stimuli regulate Myh expressions mainly through p38 and calcineurin-dependent activation of MRFs and MEF2s.
p38 MAPK
MAPKs are well-known transducers of extracellular mechanical stimuli in various cell types [62]. In particular, one major component of MAPKs, the p38 MAPK, is crucial in myoblast biology (see review refs. [63,64]). Notably, a lot of researches explored potential roles of p38 in mediating myoblast differentiation under mechanical stretching. The upstream events of stretch-activated p38 in myoblasts was Src-TACE-TNFα pathway [45,47]. In detail, the synthesized tumor necrosis factor alpha (TNFα) is cleaved and released as a secreted form of TNFα that could directly activate p38 by binding to TNFα receptor [45]. This cleaving process is catalyzed by TNFα-converting enzyme(TACE), which is phosphorylated by tyrosine kinase Src under mechanical strain [47].
Regarding the downstream factors of p38 in mediating stretch-induced myogenic differentiation, Rauch and Loughna [58] found that MEF2A, MEF2C, Myog, p21, and Myh were all activated in a p38-dependent manner [45].
However, it is still a debate whether p38 positively or negatively regulate myogenic differentiation under mechanical environment. For example, mechanical stretch was shown to inhibit both p38 phosphorylation and bovine satellite cell myogenic differentiation, implying a positive effect of p38 on myogenic differentiation [51]. Another study by the same authors confirmed persistent p38 activation during stretching of C2C12 myoblasts, and inhibitors of p38 promoted myogenic differentiation, thus manifesting the negative role of p38 in regulating myogenic differentiation [52]. Thus, p38 has multifaceted effects on myogenic differentiation in various myoblasts.
Cytoskeleton-related proteins
Normally, cells respond to external mechanical stimuli by activation of signaling pathways that often lead to cytoskeletal contraction [65]. Integrin β1D was identified as an indispensible component of the cytoskeleton-related pathways mediating myogenic differentiation in mechanical microenvironment [66]. Increased integrin β1D expression in C2C12 cells by mechanical stretch activated downstream cytoskeletal signaling proteins FAK and RhoA, and knocking down integrin β1D inhibited C2C12 differentiation [66]. On the other hand, Andersen et al. found that activation of FAK is necessary for the myoblast differentiation, since FAK inhibition compromised the stretch-prompted myogenic differentiation [49].
In addition to these proteins located on cell surface or cytoplasm, other family of proteins anchoring in the outer nuclear membrane connected actin cytoskeleton to the nuclear lamina, providing a continuous structure between ECM and nuclear lamina. For example, the integrality of LINC complex (the bridge across the nuclear membranes
The antagonizing effects of stretch on inhibitors of myogenesis
Mechanical stretching of myoblasts has been shown to antagonize the inhibitory effect of some drugs on myoblast differentiation. For example, thiazolidinedione (TZD), a specific agonist for peroxisome proliferator-activated receptor γ, suppressed the C2C12 myogenic differentiation, whereas application of cyclic stretch rescued the inhibitory effects of TZD [67]. Another case is reported by Chandran et al., who verified that exogenous added TNFα impeded the myoblast differentiation, while mechanical stretch acted as potent inhibitors of the intracellular actions of TNFα by inducing muscle cell differentiation in a proinflammatory environment [46].
Stretching-induced myoblast differentiation in muscle tissue engineering
Restoration of muscle mass and function is critical in muscle tissue engineering. Moon du et al. [68] developed a mechanical strain device to cyclically stretch human skeletal muscle cell-seeded three-dimensional scaffolds before implantation onto the latissimus dorsi muscle of mice. They found that prestretching myoblast scaffolds not only produced viable muscle tissue constructs with maturely developed multinucleated muscle fibers, but also generated tetanic and twitch contractile responses [68]. This demonstrated the importance of stretching preconditioning in vitro to accelerated skeletal myoblast differentiation, maturation, and function in vivo.
Uniaxial versus equibiaxial stretch on myoblast differentiation
Almost all the studies above explored the effects of one specific type of stretch on myoblast differentiation, using either uniaxial or equibiaxial stretching. Intriguingly, one study by Pennisi et al. compared the distinct impacts of uniaxial and equibiaxial 15% cyclic tensile stretch (CTS, 0.5 Hz) on myogenic differentiation [69]. According to their conclusion, uniaxial CTS drives C2C12 differentiation while equibiaxial CTS does not favor the differentiation process. However, even though some of the studies above corroborated this statement [48,49,51,52,54,55,67], other literatures seemed to display opposed results. In detail, some authors revealed the accelerated myoblast differentiation in response to equibiaxial stretch [44 –47,58,60,66], while others uncovered dampened or unaffected myoblast differentiation by uniaxial stretch [53,56].
Collectively, it should be concluded that these contradictory results actually reflected that mechanoresponses of myoblasts vary with the subtle changes of cellular microenvironment, such as types of stretch (cyclic/static, uniaxial/equibiaxial, intermittent/constant), as well as culturing conditions and sources of cell, etc.
Effect of Stretch on Skeletal Myoblast Mechanical Resistance, Survival, and Death
Genes regulating mechanical properties of myoblasts
When cells were stretched, the morphological extending of cell body is always accompanied by reorganization of cytoskeleton. In this regard, the accurate balance between stiffness and plasticity of cytoskeleton is critical for myoblasts to withstand and survive during physical stretching, as well as to adapt to mechanical microenvironment. One important gene in mechanosensing and making cytoskeletal adaptations is the LMNA gene (which encodes lamin A/C protein) [70]. LMNA (−/−) myoblasts are unable to resist mechanical stretching, leading to severe intracellular damage to the cytoskeleton, which was due to abnormal activation of yes-associated protein signaling [70]. This suggest that cytoskeletal plasticity and adaptability to ECM mechanical cues requires a functional lamin A/C network.
Other reported gene candidates in mediating cytoskeletal elasticity and rigidity in myoblasts are focal contact proteins talin and vinculin [71]. Zhang et al. found that static stretch enhanced the elastic modulus (E app) of C2C12 cells, through NO-induced increase in talin and vinculin expressions and decrease in calpain-mediated talin proteolysis [71].
In addition, integrin β1D was also involved in stretch-induced E app elevation in C2C12 cells, since integrin β1D knockdown prohibited the increased E app [66]. Another intriguing study by Mitsui et al. [72] displayed a time lapse sequence of stiffness distributions of living myoblasts by scanning probe microscopy when cells were stretched intermittently. They proposed that myoblasts have tensional memory effect to maintain cytoskeletal tensional homeostasis, based on the observation that cell stiffness was increased instantly after stretching but gradually decrease and return to its original level. In addition, this memory effect, which largely depends on the duration, rather than the magnitude of stretching, might be related with phosphorylation level of myosin II regulatory light chain [72].
Relationship between cytoskeletal stiffness and myoblast death under stretching
On the other hand, we could easily assume that cellular damage occurs when mechanical stretch is too severe for myoblast cytoskeleton to resist and adapt, which is often seen in muscle fiber degeneration in a number of myopathies due to higher mechanical vulnerability of skeletal muscle fibers during muscle contraction [73]. Accordingly, a study by Wang et al. [74] investigated the role of F-actin in the survival ratio of C2C12 myoblasts under 30% uniaxial stretch for 30 min. They found that some C2C12 cells died during stretching, while lowering the tension of cytoskeleton or disassembling F-actin with drugs (Blebbistatin and cytochalasin D) increased the survival rate of myoblasts. Furthermore, myoblasts with thinner morphology had more stable peripheral F-actin than wider ones, thus possessing more viability under stretching than wider ones [74].
Goldmann and colleagues also explored the involvement of cytoskeleton of human and mouse myoblasts in resisting mechanical stretching-induced cellular damage [75,76]. Desmin is a muscle-specific intermediate filament (IF) protein, the mutation of which is associated with desminopathies. By transfecting human myoblasts with a genetically heterozygous R350P desmin mutation (DesR350P), Bonakdar et al. [75] found that DesR350P myoblasts exhibit increased cell stiffness, and thus reduced cell viability in response to mechanical stretch. Consistently, other studies also demonstrated the crucial position of desmin in regulating the cytoskeletal rigidity through overexpressing mutant desmin E413K and R349P in mouse myoblasts [77,78].
In addition, plectin, a giant (4,500 kDa) cytolinker protein, anchors IFs to ECM/adhesion complexes. Interestingly, plectin-deficient mouse myoblasts had lower mechanical vulnerability upon mechanical stretch compared with wild-type cells, which was attributed to lower cellular stiffness [76]. Altogether, these studies demonstrated that cytoskeletal filament systems are sensitive to physical stretching of cells, the disruption of which directly affect the survival and death of myoblasts under mechanical stretching.
Molecular mechanisms of stretch-induced myoblast death
During the last decade, our team had thoroughly investigated how does mechanical stretch lead to myoblast apoptosis and which signaling pathways are involved. We used two skeletal myoblast cell lines, L6 and C2C12, to study the effect of cyclic stretch with different loading magnitudes and durations on apoptosis of these cells.
First, caspase-3 was the direct motivator of myoblast apoptosis in both stretching magnitude- and stretching duration-dependent manners [79,80]. Subsequently, two major organelles, endoplasmic reticulum (ER) and mitochondria, were reported to mediate stretch-activated caspase-3 pathway. Caspase-12, a specific ER-residual caspase, was cleaved at Asp94 during ER stress, which was a prerequisite to the caspase-3 activation and myoblast apoptosis [81]. Inhibition of caspase-12 by either nonspecific inhibitors or caspase-12-specific siRNA could partially rescue the myoblast death under stretching stimuli [81,82]. Apart from these, miR-147 was also found to be related with stretch-induced ER stress and myoblast apoptosis by targeting breast cancer metastasis suppressor 1 during mechanical stretching of L6 myoblasts. Stretching stimuli suppressed miR-147 expression, thus favoring ER stress and apoptosis of myoblasts [83].
On the other hand, mitochondrion was another important organelle involving in stretching-induced myoblast death, with mitochondrial maker protein apoptosis-inducing factor (AIF) level being elevated during mechanical stretch [84]. Furthermore, we found a stretching magnitude-dependent increment of reactive oxygen species (ROS) in C2C12 myoblasts, which activated two pathways that contribute to myoblast death: JNK and p53 [85,86]. When ROS was inhibited, stretch-activated JNK was also attenuated; inhibition of both ROS and JNK could alleviate myoblast apoptosis under stretching stimuli [85]. Furthermore, ROS-induced p53 mitochondrial translocation only occurred when ROS was overaccumulated by stretching stimuli, which was a critical event leading to myoblast apoptosis, possibly depending upon classical mitochondrial apoptotic pathways [86].
Effect of Stretch on Properties of Plasma Membrane of Skeletal Myoblasts
It could be easily accepted that when myoblasts are mechanically stretched, the PM should be the first structure being affected, and therefore, the mechanoresponse of PM is critical in initiating the following cellular changes (behaviors, structures, functions, gene expressions, etc.) during mechanical stretching. Caveolae are flask-like invaginations of the PM that is organized by caveolins. By briefly stretching myoblasts (1 min), Gervasio et al. proposed that stretch induced reversible folding and unfolding of caveolae and activation of Src kinase. Furthermore, overexpression of caveolin-3 potentiated the stretch-evoked activation of Src kinase in myoblasts. These results were attributed to the redistribution of caveolin-3 from intracellular pools to cell surface [87]. Thus, the dynamical changes of these microstructures on PM might reflect the initial responses of myoblasts to mechanical stretch.
On the other hand, Leopold and Gefen explored the permeability of the PM of myoblasts being radially or uniaxially stretched. Using a wide range of stretching magnitude from 3% to 12%, they suggested that mechanical deformation of PM induced some nonspecific pores/tears in PM, the number and size of which increased in a magnitude-dependent manner [88,89]. This will allow for the build-up of intracellular concentrations of biomolecules (ions, metabolites, free radicals, hormones, etc.) that are present at the extracellular space, leading to metabolic changes to cells. Specifically, O2 transport in C2C12 myoblasts (either O2 influx into cells or loss of intracellular O2) became faster with the increasing levels of stretching magnitude, due to either increased PM permeability or cell surface area [90].
Effect of Stretch on Multipotency of Transdifferentiation of Skeletal Myoblasts
It has been reported that skeletal myoblasts possess pluripotent differentiation potential in vitro and in vivo [91]. Mechanical stretch, as a unique external stimulus, has been proposed to affect the pluripotency of skeletal myoblasts. For example, coculture of skeletal myoblasts with cardiomyocytes promoted skeletal myoblasts to express cardiac-specific proteins such as cardiac troponin T, atrial natriuretic peptide, Nkx2E, and GATA4, which was inhibited by addition of nifedipine or culture in Ca2+-free media. Stretching the skeletal myoblasts cocultured with cardiomyocytes completely restored this inhibitory effect and promoted transdifferentiation into cardiomyocytes [92].
Another research confirmed that C2C12 cells differentiate into adipocytes in adipogenic induction medium, while mechanical stretch prohibited this process through Wnt signaling [93]. In addition, Kim et al. found that bone morphogenetic protein 2 (BMP-2) treatment enhanced the gene expression of osteogenic markers (ALP, type I collagen, osteopontin, osteocalcin, cbfa1, osterix, and dlx5) in C2C12 myoblasts. BMP-2 combined with mechanical stretching of C2C12 further potentiated the expressions of these genes, demonstrating the positive effect of stretch on osteodifferentiation of myoblasts [41].
Collectively, these studies corroborated two points in common: (1) skeletal myoblasts possess the multipotency of transdifferentiation into other types of cells when cultured in special conditions, and (2) mechanical stretching either positively or negatively affected the progressing of myoblast transdifferentiation.
Effect of Stretch on Reorientation and Alignment of Skeletal Myoblasts
It has been demonstrated that skeletal myoblast orientation and internal cell structure displayed dynamical changes in response to mechanical stretch, largely depending on the direction of the stretch. To realign themselves from a randomly aligned state, skeletal myoblasts first need to be able to migrate. One study conducted by Marom et al. found that radial stretch combined with exogenous sodium pyruvate (NaPy) supplements accelerated C2C12 migration rate, in contrast to control group only supplemented with NaPy [94]. Subsequently, Ahmed et al. explored the adaptive responses of C2C12 skeletal myoblasts seeded on fibronectin lines with different orientations relative to the stretching direction. After application of uniaxial stretch (amplitude of 7%, frequency of 0.5 Hz), they found that actin fiber organization is predominantly controlled by stretch [95].
Another interesting study by Leccia et al. [96] recorded the dynamics of reorientation of C2C12 myoblasts after 1, 3, and 6 hours of uniaxial stretch (amplitude of 20%, frequency of 0.3 Hz). Cells retracted after the first stretching hour and started to spread again after 3 h of stretch. At 6 h of stretching, cells realigned themselves at the mean orientation of 66.5° relative to the direction of stretch. Desmin plays a crucial part in this process, since C2C12 myoblasts expressing mutant desmin D399Y failed to display the same mechanoresponses as control cells during stretch, leading to less retracting, spreading, and mean orientation angle [96].
Similarly, there were other two literatures reporting the reorientation of myoblasts being extended by uniaxial stretch [48,49]. However, both studies showed that myoblasts aligned orthogonal to the direction of stretch, in contrast to the orientation of 66.5° in Leccia et al.'s [96] research. There is no clear illumination addressing this deviation among these studies. However, we suspected the stretching duration might partially account for this problem, since the study by Leccia et al. [96] applied stretching for 6 h, while the studies showing perpendicular orientation stretched myoblasts for 24 h and 4 days, respectively.
Notably, all the researches above manifested uniaxial stretch-induced reorientation of myoblasts. By comparing uniaxial and equibiaxial stretches, Pennisi et al. further demonstrated that while myoblasts aligned vertically to the direction of stretch when extended uniaxially, they had no clear uniform orientation and displayed signs of membrane damage when extended equibiaxially [69].
Contradictory Responses of Myoblasts to Mechanical Stretching
The cellular responses of myoblasts to mechanical stimuli (proliferation, differentiation, apoptosis, etc.) seemed to be opposed to each other. One illumination is apparently because of different experimental designs, including stretching parameters and culturing conditions, which distinctly affect the intracellular signaling and gene expression profile, thus leading to substantially different reactions of myoblasts.
However, cell heterogeneity is also an ignored problem considering the multifaceted effect of stretching on myoblasts [97,98]. For example, in our previous studies, we had demonstrated that even though myoblast apoptotic rate increased in response to stretching of longer duration and higher magnitude, there were still some cells surviving after exposure to these intensive stretches. We suspected these cells might possess some “mechanical resistant” properties that are absent from the apoptotic cells, and possibly undergo proliferation or differentiation under these proapoptotic stretching conditions. Single-cell analysis will provide better insights into the complicated phenotypes of myoblasts during mechanical stimulation, considering cell heterogeneity as a factor contributing to multiple responses of cells to mechanical stimuli.
Interconnections Among the Mechanosensitive Pathways in Myoblasts
Notably, some pathways were frequently mentioned in different studies, such as Src [22,38,47,87], NF-κB [22 –24,26,54,85], p38 [26,31,32,38,45,47,51,52,54,58], FAK [22,49,66], IGF-1 [28 –32], NOS [16,21,23,46,66,71], AKT [31,32,40,45,86], and mTOR [38 –40], in spite of various stretching parameters being used in these studies. To some extent, this suggests these pathways are more mechanosensitive, and are more easily activated during distinct type of mechanical strain. Furthermore, we could assume that under one specific mechanical stimulus, more than one of these pathways might be involved in transducing mechanical signal and in mediating cellular adaptations to external straining environment. Thus, understanding about the interconnections among these pathways should shed some light on the clear molecular mechanisms regulating the responses of myoblasts to mechanical stretching (Fig. 3).
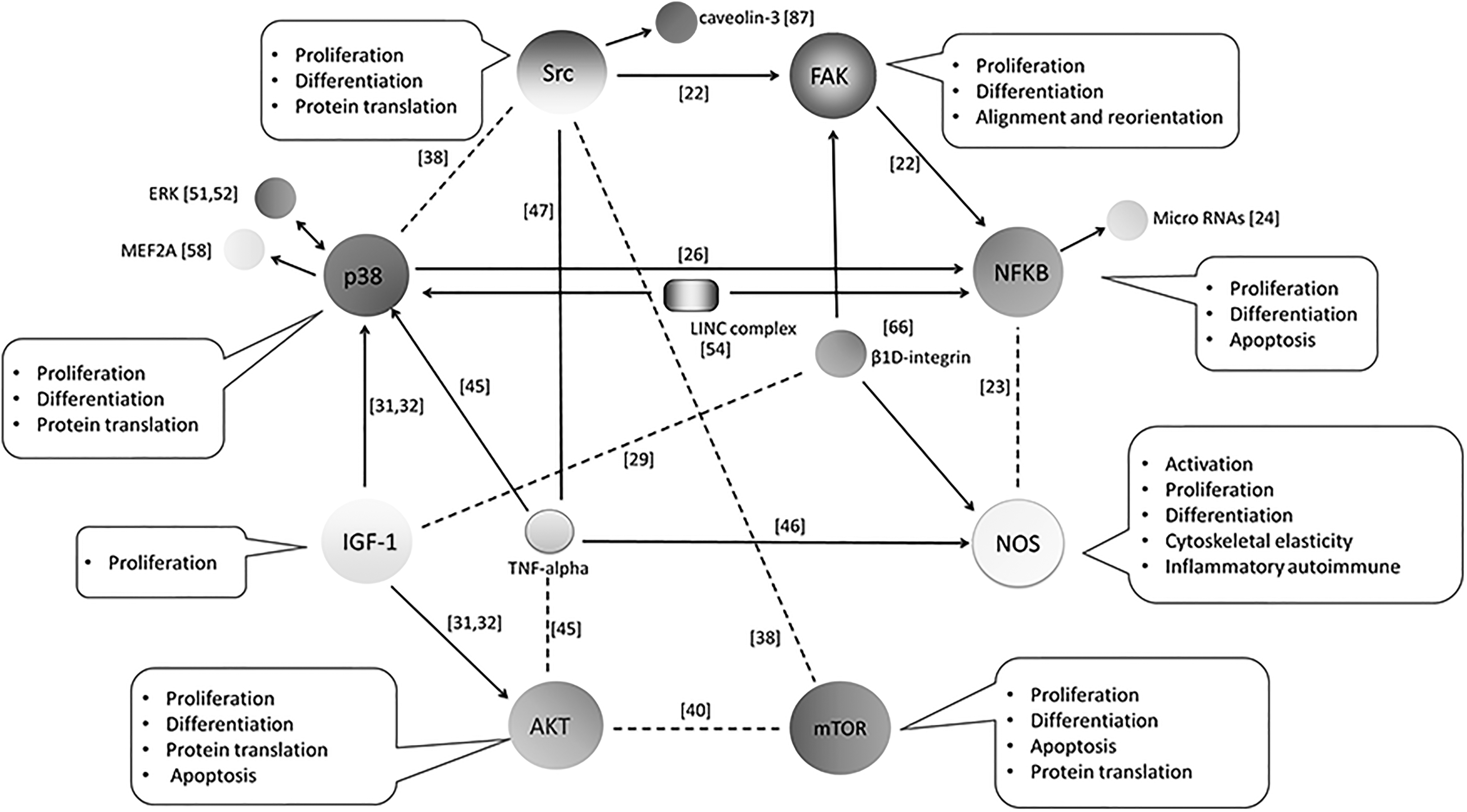
Interconnections among the mechanosensitive pathways in skeletal myoblasts. In this review, several pathways were reported to be sensitive to various regimens of mechanical stretch, as displayed in different references. In addition, they were demonstrated to play distinct roles in mechanoresponses of myoblasts, which are summarized in the annotations beside each one of them. Notably, there were interconnections among these pathways in stretched myoblasts, with arrow line indicating direct regulation between two pathways, and dotted line suggesting possible indirect relation between two pathways.
Perspectives
Identification of molecules and pathways that integrate mechanoresponses of skeletal myoblasts will be an important key for unlocking a better understanding of the complex physiological and pathological changes of skeletal muscle under mechanical stimuli. During the past decades, substantial works have been done in progressing the knowledge of multiple responses of skeletal myoblasts to mechanical stretching, which will enhance our comprehension about the myoblast contributions to macroscopical muscular changes (structure, mass, function, etc.) during physical exercise, loading bearing, or medical treatment.
However, what remains elusive is the mechanisms by which all these genes and signaling pathways work together to give a unified response resulting in altered phenotypes and behaviors of skeletal myoblasts due to mechanical loading, as we have partially overviewed in the last part of this review. A more global view of integration of intracellular molecules and pathways is necessary for development of specific and effective countermeasures for maximizing the beneficial skeletal muscle changes while minimizing or ablating deleterious side effects during external mechanical loading. New technologies such as microarray analysis and single-cell analysis should pave the way for the study of skeletal myoblast mechanoresponses, rather than the examination of separate individual components, as had been done before. On the other hand, strain regimens comprising amplitude, duration, orientation, frequency, and rest period, should be optimized in search of a more advantageous mechanical environment that is conducive to skeletal muscle homeostasis.
Overall, a better understanding of myoblast biology during external stretching, appropriate countermeasures for the side effects of mechanical loading, and optimal straining environment will promote the developments of muscular biomechanics, regeneration and reconstruction of muscle, medicine, kinesiology, etc.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (no.11702154), the China Postdoctoral Science Foundation (no. 2018M642621), and the Qingdao Outstanding Health Professional Development Fund.