
Abstract
Mesenchymal stem cells (MSCs) are multipotent progenitors that have the ability to differentiate into multiple lineages, including bone, cartilage, and fat. We previously demonstrated that the least known bone morphogenetic protein (BMP)9 (also known as growth differentiation factor 2) is one of the potent osteogenic factors that can induce both osteogenic and adipogenic differentiation of MSCs. Nonetheless, the molecular mechanism underlying BMP9 action remains to be fully understood. Leptin is an adipocyte-derived hormone in direct proportion to the amount of body fat, and exerts pleiotropic functions, such as regulating energy metabolism, bone mass, and mineral density. In this study, we investigate the potential effect of leptin signaling on BMP9-induced osteogenic differentiation of MSCs. We found that exogenous leptin potentiated BMP9-induced osteogenic differentiation of MSCs both in vitro and in vivo, while inhibiting BMP9-induced adipogenic differentiation. BMP9 was shown to induce the expression of leptin and leptin receptor in MSCs, while exogenous leptin upregulated BMP9 expression in less differentiated MSCs. Mechanistically, we demonstrated that a blockade of JAK signaling effectively blunted leptin-potentiated osteogenic differentiation induced by BMP9. Taken together, our results strongly suggest that leptin may potentiate BMP9-induced osteogenesis by cross-regulating BMP9 signaling through the JAK/STAT signaling pathway in MSCs. Thus, it is conceivable that a combined use of BMP9 and leptin may be explored as a novel approach to enhancing efficacious bone regeneration and fracture healing.
Introduction
Mesenchymal stem cells (MSCs) are multipotent progenitor cells that can renew themselves and, upon appropriate stimuli, differentiate into multiple lineages such as osteoblastic, chondrogenic, adipogenic, and myogenic lineages [1 –6]. Thanks to their easy access and abundance, MSCs have received a great deal of attention in the fields of stem cell research and regenerative medicine, especially in the area of bone regeneration and tissue engineering [4,7 –10]. Osteogenic differentiation of MSCs is regulated by several major signaling pathways [3,11 –20]. Among them, bone morphogenetic proteins (BMPs) have been considered the most potent osteoinductive factors [21 –23]. BMPs are members of the transforming growth factor β (TGF-β) superfamily [3,21,22,24], and at least 15 BMPs have been identified in humans and rodents [21,22,25].
Through a comprehensive analysis of the osteogenic activity of the 14 types of human BMPs, we identified the previously least known BMP9 (also known as growth differentiation factor 2) as one of the most potent BMPs that induce osteogenic differentiation of MSCs both in vitro and in vivo [21,23,26 –29], at least in part, owing to BMP9's resistance to the naturally occurring antagonist noggin [30]. We further demonstrated that the TGF-β/BMP type I receptors activin receptor-like kinase 1 (ALK1) and ALK2 are critical to BMP9-initiated osteogenic signaling in MSCs [31]. Nonetheless, the exact molecular mechanisms through which BMP9 induces osteogenic differentiation of MSCs and maintain bone homeostasis are not fully understood.
Discovered about 25 years ago as an adipocyte-derived hormone produced in direct proportion to the amount of body fat, leptin has pleiotropic functions and regulates food intake, energy metabolism, the reproductive system, inflammation, immunity, and bone mass and mineral density [32 –37]. Leptin is also expressed in non-adipose tissues and initiates its functions by binding to its specific receptor, leptin receptor (LepR), which subsequently activates a cascade of intracellular signaling events, including JAK/STAT, insulin receptor substrate/PI3K, SH2-containing protein tyrosine phosphatase 2/MAPK, and AMPK/acetyl-CoA carboxylase, in the central nervous system and peripheral tissues [33,38]. It was reported that LepR was a marker for bone marrow MSCs, and the LepR+ cells formed bone, cartilage, and adipocytes in culture and upon transplantation in vivo [39]. Fate-mapping demonstrated LepR+ cells appeared postnatally and generated the majority of bone and adipocytes found in adult bone marrow, including bone that was regenerated postinjury. This suggests that the quiescent LepR+ cells may be the major source of bone and adipocytes in adult bone marrow [39]. Further studies revealed that leptin can act directly on LepR-expressing MSCs in adult bone marrow to influence their lineage allocation in vivo, by inhibiting bone formation and inducing marrow adipogenesis [34].
While leptin is well recognized to regulate bone mass and density by stimulating osteoblastic cells and promoting osteogenesis, it can indirectly inhibit bone formation by stimulating sympathetic neurons in the hypothalamus [32,35]. In fact, conflicting results of leptin on bone formation have been reported [40 –44]. While the leptin-deficient mice have reduced length and bone mineral content of long bones, but increased vertebral trabecular bone [41], a consistent bone phenotype in human leptin deficiency has not been established [32]. However, systemic leptin administration in animals and humans usually exerts a positive effect on bone mass [32]. Thus, the balance of the central and peripheral effects of leptin on bone and the skeleton system remains an area of substantial controversy.
In this study, we investigate the potential effect of leptin signaling on BMP9-induced osteogenic differentiation of MSCs. We found that exogenous leptin potentiated BMP9-induced osteogenic differentiation of MSCs in vitro and in vivo. BMP9 was shown to induce the expression of leptin and LepR in MSCs, while exogenous leptin was able to induce BMP9 expression in MSCs with higher stemness, but not in more differentiated MSCs. Mechanistically, a JAK1/2 inhibitor effectively blunted the leptin-potentiated osteogenic differentiation induced by BMP9. Collectively, our findings strongly suggest that leptin may potentiate BMP9-induced osteogenesis by cross-regulating BMP9 signaling through the JAK/STAT signaling pathway in MSCs. Thus, it is conceivable that a combined use of BMP9 and leptin may represent a novel approach to enhancing efficacious bone regeneration and fracture healing.
Materials and Methods
Cell culture and chemicals
The mouse MSC lines, imBMSCs and iMEFs, are conditionally immortalized mouse bone marrow stromal cells (imBMSCs) and immortalized mouse embryonic fibroblasts (iMEFs), respectively, as described [45 –47]. The 293pTP and RAPA cells were previously characterized HEK293 derivative cells [48,49]. All cell lines were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS; Gemini Bio-Products, West Sacramento, CA), supplemented with 100 U/mL penicillin and 100 mg/mL streptomycin, incubated at 37°C in 5% CO2 as described [50 –53]. JAK1/2 selective inhibitor Ruxolitinib was purchased from LC Laboratories (Woburn, MA). Unless indicated otherwise, other reagents were purchased from Sigma-Aldrich (St. Louis, MO) or Thermo Fisher Scientific (Waltham, MA).
Construction and amplification of recombinant adenoviruses expressing BMP9, Lep, simLep, and GFP
The AdEasy technology was used to construct recombinant adenoviruses as described [54 –56]. Briefly, the coding region of human BMP9 and mouse leptin were PCR amplified, subcloned into an adenoviral shuttle vector, and subsequently used to generate recombinant adenoviruses in HEK293, 293pTP, or RAPA cells as described [48,49,57], resulting in AdBMP9 and AdRLep.
For constructing AdRsimLep, three siRNAs targeting the coding region of mouse leptin were designed by using Invitrogen's BLOCK-IT RNAi Designer program, and simultaneously assembled to our homemade adenoviral shuttle vector as described [51,58,59]. An analogous adenovirus expressing GFP (AdGFP) or RFP only was used as a mock virus control [60 –62]. For all adenoviral infections, polybrene (4–5 μg/mL) was used to enhance infection efficiency as described [63].
RNA isolation and touchdown quantitative real-time PCR
Total RNA was extracted with TRIZOL Reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions and subjected to reverse transcription reactions using hexamer and M-MuLV Reverse Transcriptase (New England Biolabs, Ipswich, MA). The resultant cDNA products were diluted and used as PCR templates. The Primer3 Plus program was used to design touchdown quantitative real-time PCR (TqPCR) primers [64]. TqPCR analysis was performed as described [65 –68]. Briefly, SYBR Green (Bimake, Houston, TX) qPCR reactions were set up according to manufacturer's instructions using the following cycling program: 95°C × 3 min for 1 cycle; 95°C × 20 s, 66°C × 10 s per cycle, and then −3°C per cycle for 4 cycles; followed by 95°C × 10 s, 55°C × 15 s, and 70°C × 1 s for 40 cycles. GAPDH was used as a reference gene. The gene expression calculation was performed using the 2−ΔΔCt method [69 –71]. TqPCR primer sequences are listed in the Supplementary Table S1.
Alkaline phosphatase assays
The alkaline phosphatase (ALP) activities were determined both qualitatively and quantitatively using the histochemical staining and modified Great Escape SEAP chemiluminescence assay (BD Clontech) as described previously [72 –74]. Each assay condition was conducted in triplicate.
Matrix mineralization assay (Alizarin Red S staining)
The cells were seeded in 24-well plates, infected with the adenoviruses, and cultured with 10% FBS DMEM containing ascorbic acid (50 mg/mL) and β-glycerophosphate (10 mM). At the indicated time points, Alizarin Red S staining was performed to assess mineralized matrix nodules as described [61,75 –78]. The staining of calcium mineral deposits was recorded under bright field microscopy. For quantitative assays, the Alizarin Red S stains were dissolved in 10% acetic acid. Photometric absorbance was measured at 405 nm.
Subcutaneous stem cell implantation and ectopic bone formation
The use and care of athymic nude mice were approved by the Institutional Animal Care and Use Committee (the ACUP protocol No. 71108). All experimental procedures were performed according to the approved guidelines. Subcutaneous stem cell implantation procedure was carried out as previously described [79 –85]. Briefly, subconfluent imBMSCs and iMEFs were co-infected with appropriate combinations of adenoviruses for 36 h, and then collected, resuspended in phosphate-buffered saline (PBS, ∼50 μL per injection), and injected subcutaneously into the flanks of nude mice (Envigo/Harlan Research Laboratories; n = 5/group, female, 5 weeks old; 2 × 106 cells per injection). At the indicated endpoints, nude mice were sacrificed and the masses were retrieved from injection sites for microcomputed tomographic (μCT) imaging and histologic evaluation.
μCT imaging and data analysis
The retrieved bony masses were fixed in 10% PBS-buffered formalin and subjected to μCT imaging by using the GE triumph (GE Healthcare) trimodality preclinical imaging system. The acquired imaging data were quantitatively analyzed using Amira 6.0 (Visage Imaging, Inc.) as described [45,57,61,75,84].
Hematoxylin and eosin and Masson's trichrome staining
After being μCT imaged, the fixed samples were decalcified and paraffin embedded. Serial sections of the embedded samples were subjected to hematoxylin and eosin (H&E) staining and Masson's trichrome staining as previously described [46,72,78,83].
Oil Red O staining assay
Subconfluent iMEFs were seeded in 24-well cell culture plates, and infected with indicated adenoviruses for 7 or 14 days. Oil Red O staining was carried out as described [83,86]. Briefly, the cells were fixed with 10% formalin at room temperature for 10 min and were washed with PBS. After residual PBS being aspirated, the fixed cells were stained with freshly prepared Oil Red-O solution at 37°C in an incubator for 60 min, followed by washing with distilled water. For quantitative analysis, the Oil Red O stains were dissolved with 100% 2-propanol and the photometric absorption was determined at 510 nm.
Statistical analysis
The quantitative experiments were performed in triplicate and/or repeated in three independent batches. Data were expressed as mean ± standard deviation. The one-way analysis of variance was used to analyze statistical significance. A value of P < 0.05 was considered statistically significant.
Results
BMP9 induces leptin expression, while exogenous leptin potentiates BMP9-induced early osteogenic marker ALP in MSCs
We first analyzed whether BMP9 stimulation would impact leptin expression in MSCs. Using imBMSC progenitor cells, we found that, as for the known target genes Smad6 and Id1, leptin expression was significantly upregulated by BMP9, compared with that of the GFP control group (Fig. 1A). To effectively express exogenous leptin, we constructed the recombinant adenoviral vector AdRLep, which can infect imBMSC cells with high efficiency (Fig. 1B). Furthermore, the AdRLep-infected imBMSC cells exhibited marked increase in leptin expression, when compared with that of the GFP group (Fig. 1C[a]).
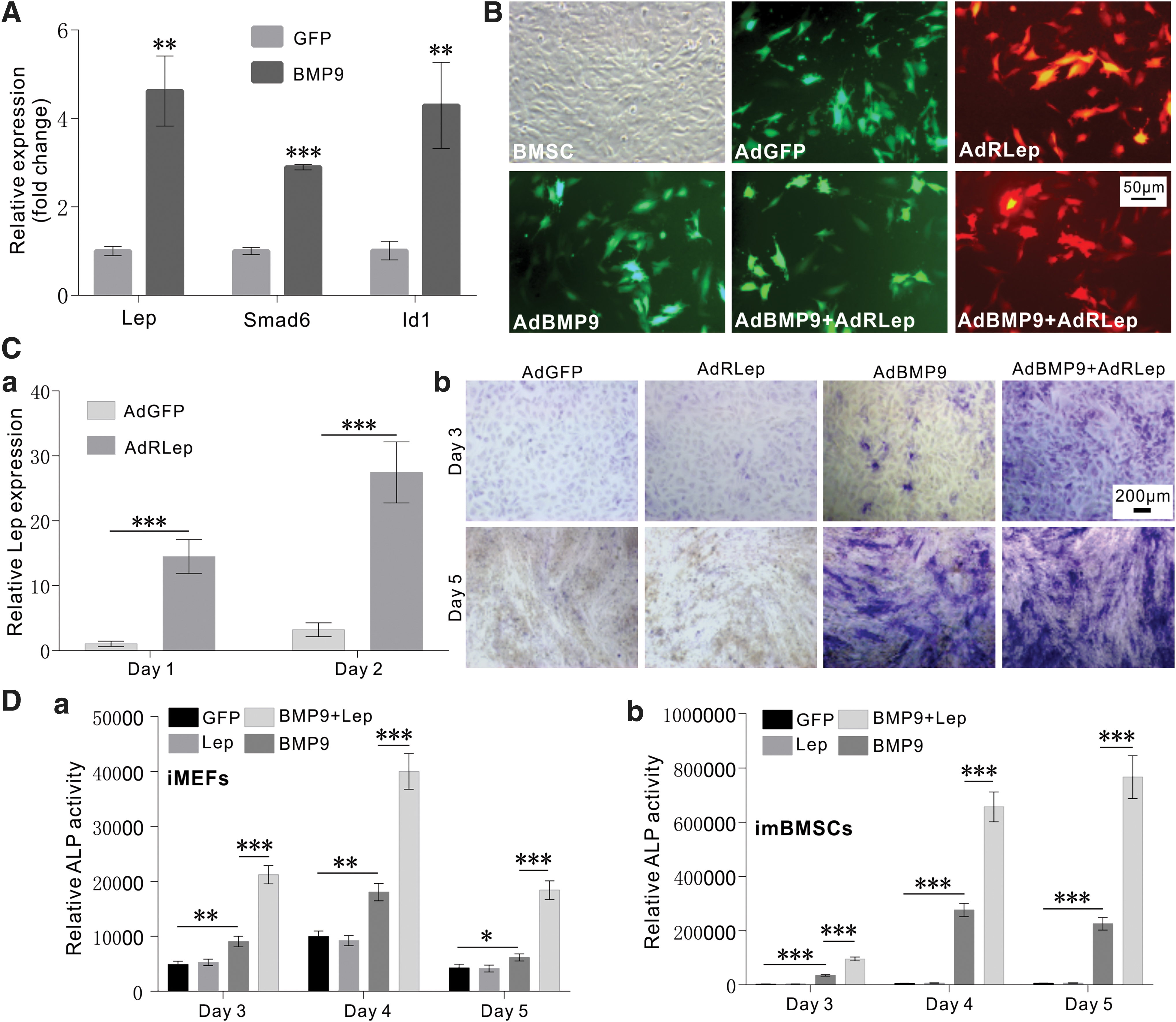
Exogenous Lep potentiates BMP9-induced early osteogenic marker ALP in MSCs.
We further analyzed the effect of exogenous leptin on BMP9-induced activity of the early osteogenic marker ALP (ALK) in MSCs. When iMEF cells were infected with AdGFP, AdRLep, and/or AdBMP9 alone or in combination, histochemical staining indicated that BMP9-induced ALP activity was significantly potentiated in the presence of exogenous leptin on both day 3 and 5 after stimulation, although leptin itself did not induce detectable ALP activity (Fig. 1C[b]). Quantitative ALP assay further confirmed that exogenous leptin significantly enhanced BMP9-induced ALP activity at the three time points examined in both iMEF cells (Fig. 1D[a]) and imBMSC cells (Fig. 1D[b]).
Conversely, we tested whether silencing endogenous leptin would impact BMP9-induced osteogenic differentiation of MSCs. We constructed a recombinant adenovirus AdRsimLep and demonstrated that it transduced iMEFs with high efficiency (Fig. 2A[a]) and effectively knocked down endogenous leptin in MSCs (Fig. 2A[b]). When imBMSC cells were infected with AdRsimLep, we found that BMP9-induced ALP activity was not significantly affected (Fig. 2B[a, b]). Thus, these results indicate that silencing endogenous leptin expression does not significantly impact BMP9-induced osteogenic differentiation in MSCs.

Silencing endogenous Lep expression does not significantly impact BMP9-induced osteogenic differentiation in MSCs.
Exogenous leptin potentiates BMP9-induced late stage of osteogenic differentiation, but inhibits adipogenic differentiation in MSCs
We further analyzed the effect of leptin on BMP9-induced matrix mineralization in MSCs. When iMEF cells were infected with AdBMP9, or AdGFP, AdR-Lep, and/or AdR-simLep, and cultured in mineralization medium, we found that overexpression of leptin significantly enhanced BMP9-induced matrix mineralization on day 14, as assessed by Alizarin Red S staining, although silencing endogenous leptin marginally decreased BMP9-induced matrix mineralization (Fig. 3A[a, b]).

Exogenous Lep potentiates BMP9-induced late stage of osteogenic differentiation, but inhibits adipogenic differentiation in MSCs.
As BMP9 is able to induce adipogenic differentiation of MSCs [22,23,29,86], we examined the effect of leptin on BMP9-induced adipogenic differentiation. When subconfluent iMEFs were infected with AdBMP9 or AdGFP, AdR-Lep, and/or AdR-simLep, and subjected to Oil Red O staining, we found overexpression of leptin significantly diminished BMP9-induced adipogenic differentiation (Fig. 3B[a, b]). Collectively, these in vitro results indicate that exogenous leptin potentiates osteogenic differentiation, but inhibits adipogenic differentiation of MSCs induced by BMP9.
Exogenous leptin augments BMP9-induced ectopic bone formation of MSCs in vivo
We further analyzed the in vivo effect of leptin on BMP9-induced ectopic bone formation from MSCs. When iMEFs were co-infected with AdBMP9 or AdGFP, AdRLep, and/or AdRsimLep and injected subcutaneously into the flanks of athymic nude mice, we found bony masses in AdBMP9, AdBMP9 + AdRLep, and AdBMP9 + AdRsimLep groups. Gross assessment indicated that co-expression of BMP9 and leptin yielded larger bony masses, compared with BMP9 alone (Fig. 4A[a]), which was confirmed by μCT imaging (Fig. 4A[b]). Quantitative analysis of the μCT imaging data revealed that leptin expression significantly augmented the average bone volume (Fig. 4A[c]) and increased the mean bone density (Fig. 4A[d]) induced by BMP9. The H&E histologic analysis revealed that thickest trabecular bone and highest overall bone matrix mineralization were found in the BMP9 + Lep group, while silencing Lep expression seemingly weakened the trabecular bone thickness and overall matrix mineralization (Fig. 4B[a]). These results were further confirmed by trichrome staining. The samples from the BMP9 + Lep group exhibited highly mineralized and mature bone matrix, compared with that of the BMP9-only group or BMP9 + simLep group (Fig. 4B[a, b]). These results suggest that that leptin overexpression may enhance the BMP9-induced trabecular bone density and maturity. Collectively, the above in vitro and in vivo results demonstrate that leptin effectively potentiates BMP9-induced osteogenic differentiation of MSCs.
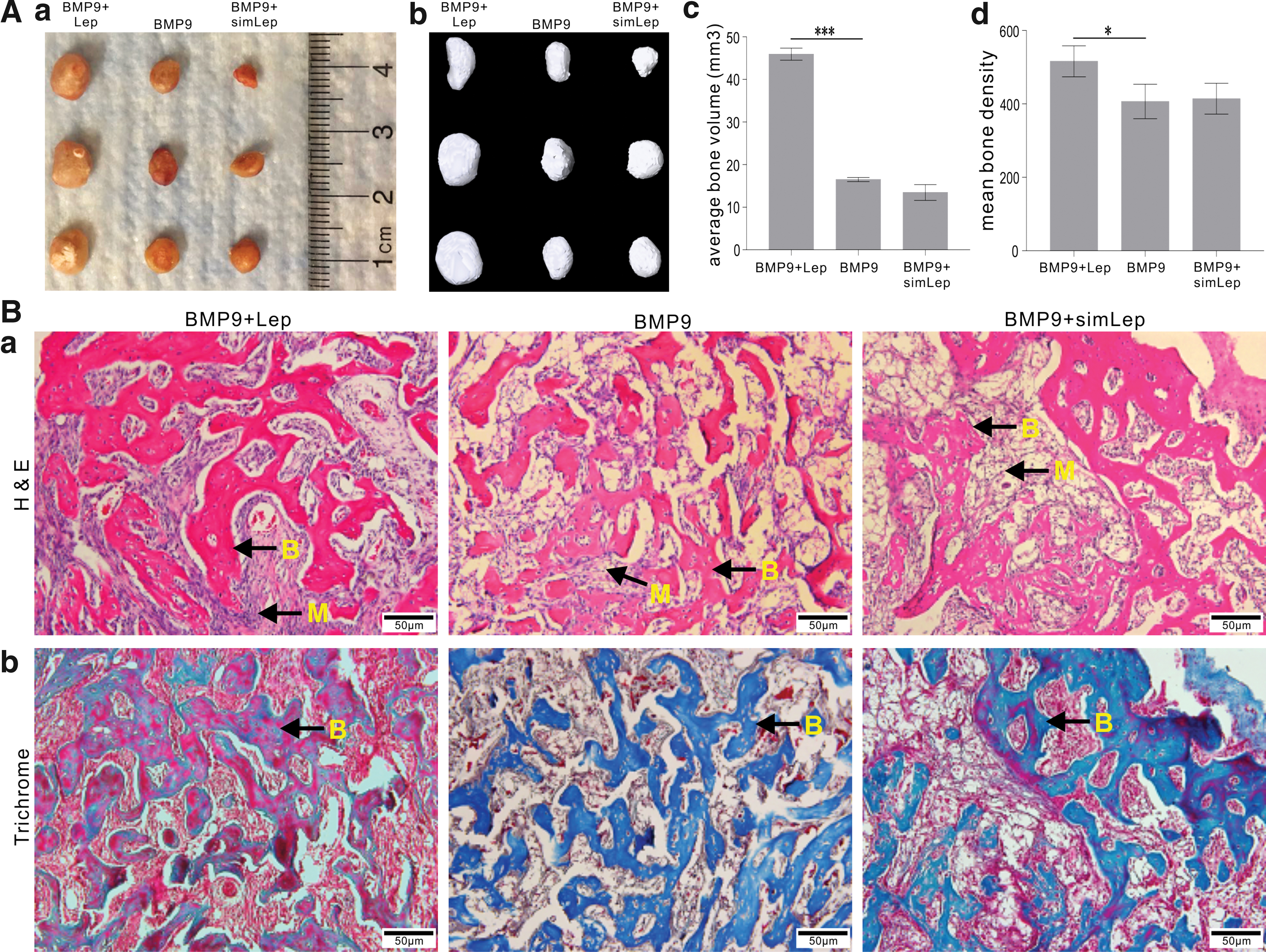
Exogenous Lep augments BMP9-induced ectopic bone formation of MSCs in vivo.
Leptin cross-regulates BMP9 signaling through the JAK2 signaling pathway
To explore the possible molecular mechanism underlying the potentiation effect of leptin on BMP9-induced osteogenesis, we analyzed whether BMP9 regulated leptin expression in MSCs, or vice versa. When iMEFs and imBMSCs were stimulated with BMP9, we found that leptin expression was significantly upregulated at 48 h after AdBMP9 infection in both lines, while leptin was noticeably induced in iMEFs at as early as 24h (Fig. 5A[a] vs. Fig. 5A[b]). Similarly, leptin receptor was also upregulated by BMP9 in both iMEFs and imBMSCs at 48 h after AdBMP9 infection (Fig. 5A[c] vs. Fig. 5A[d]). Conversely, we also analyzed if leptin affected BMP9 expression and found that exogenous leptin effectively induced BMP9 expression in the iMEFs, but not in the imBMSCs (Fig. 5B[a] vs. Fig. 5B[b]). These results indicate that BMP9 and leptin can cross-regulate each other in MSCs.
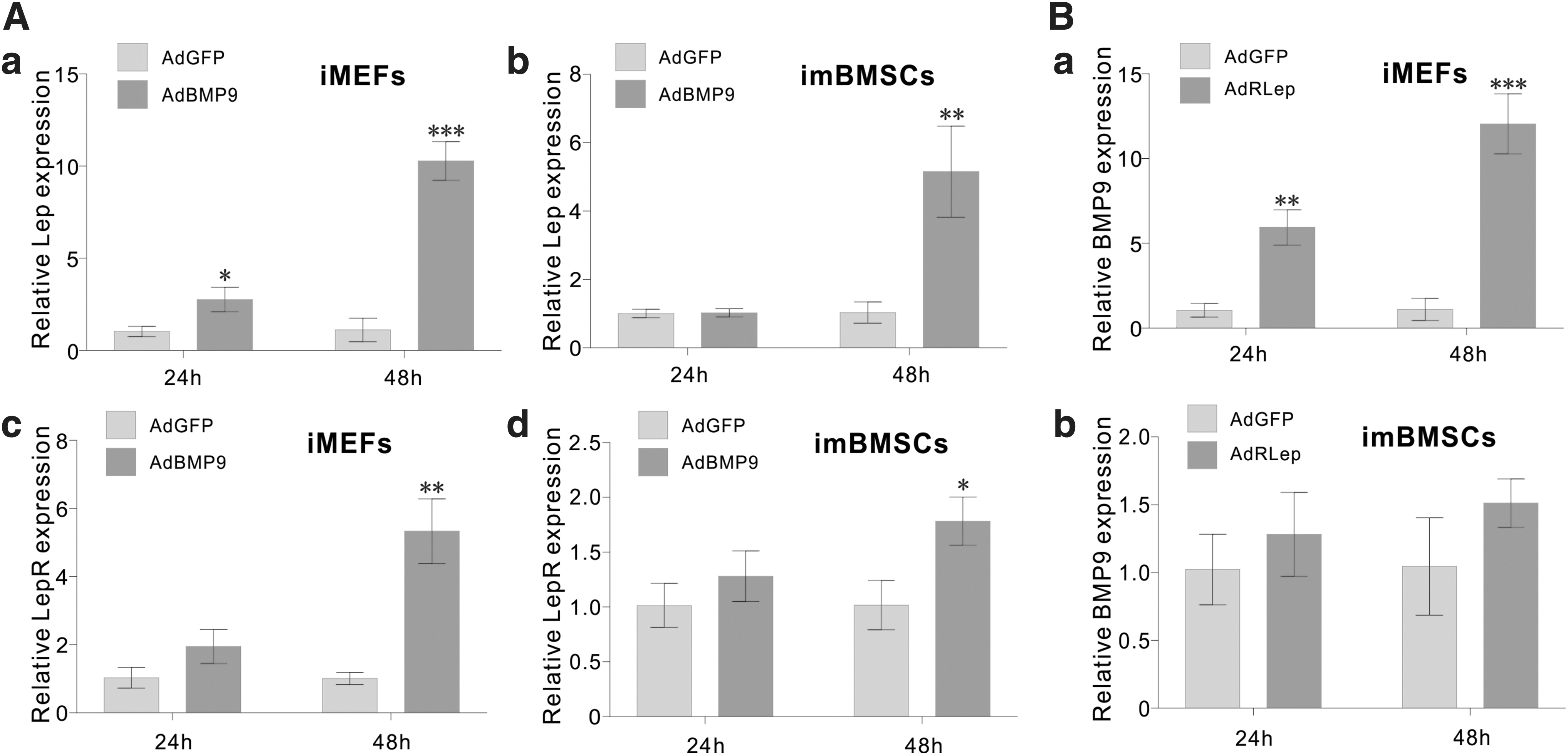
Cross-regulation of BMP9 and Lep signaling in MSCs.
As leptin activates several signaling pathways, including the JAK/STAT signaling pathway [33,38], we investigated if a blockade of JAK signaling affected the potentiation effect of leptin on BMP9-induced osteogenic differentiation of MSCs. When iMEFs were infected with AdBMP9, AdGFP, and/or AdRLep, and treated with JAK1/2 inhibitor ruxolitinib or dimethyl sulfoxide solvent, we found that leptin-enhanced BMP9-induced ALP activity was effectively blunted by ruxolitinib on both day 3 and 4 time points, although the ALP activity induced by BMP9 alone was also inhibited to certain extent (Fig. 6A[a]). These results were also confirmed by qPCR analysis, which revealed that leptin-enhanced BMP9-induced ALP expression was significantly diminished by ruxolitinib in the iMEFs (Fig. 6A[b]). We further analyzed the effect of ruxolitinib on the expression of downstream target genes of BMP9 and leptin signaling pathways, and found that leptin-induced interleukin-6 expression was significantly blunted by ruxolitinib (Fig. 6B[a]). Accordingly, leptin-enhanced BMP9-induced expression of Col1a1 and OPG was effectively inhibited by ruxolitinib (Fig. 6B[b, c]). Interestingly, BMP9-induced expression of PPARγ2 and LPL was not affected by ruxolitinib treatment while leptin-mediated inhibition of BMP9-induced expression of PPARγ2 and LPL was alleviated by ruxolitinib in MSCs (Fig. 6C[a, b]). Collectively, these results demonstrated that leptin may cross-regulate BMP9 signaling through the JAK/STAT signaling pathway during BMP9-induced osteogenic differentiation of MSCs.

The potentiation effect of Lep on BMP9 signaling can be effectively blunted by inhibiting the JAK2 signaling pathway.
Discussion
While numerous studies have examined the physiological and pathological effects of leptin on bone metabolism and skeletal homeostasis, we primarily focused on the potential synergistic effect between leptin and osteogenic factors, in this case BMP9, on osteogenic differentiation of MSCs. Our in vitro and in vivo studies indicate that exogenous leptin effectively potentiates BMP9-induced osteogenic differentiation of MSCs, while exerting inhibitory effect on BMP9-induced adipogenesis of MSCs. Furthermore, our results demonstrate that exogenous leptin alone exerts no detectable osteogenic effect on MSCs under our experimental conditions.
Mechanistically, we demonstrate that leptin may exert its synergistic effect on BMP9-induced osteogenic signaling by cross-regulating each other and activating JAK/STAT signaling. Interestingly, we previously showed that BMP9 can induce the expression of growth hormone and growth hormone-regulated JAK/STAT signaling also plays an important role in BMP9-induced osteogenic differentiation of MSCs [23,73].
We and others have shown that BMP9 exerts its pleiotropic functions through cross talk with several major signaling pathways, such as WNT/β-catenin, NOTCH, IGFs, EGF, TGF-β, Hedgehog, HIF1α, NELL1, and retinoic acids [3,11 –19,23,50,72,76,82 –85]. Interestingly, BMP9 has also been shown to play an important role in regulating glucose metabolism and insulin resistance [87,88]. It was reported that, in patients with metabolic syndrome, circulating BMP9 levels were significantly lower in patients with metabolic syndrome patients, or type 2 diabetes mellitus, compared to those of healthy controls [89,90]. We previously demonstrated that BMP9 and several osteogenic BMPs effectively induce adipogenic differentiation of MSCs [22,23,86]. Thus, it would be interesting to investigate whether the cross talk between BMP9 and leptin plays any role in regulating adiposity and energy metabolism, in addition to its role in regulating osteogenesis.
In summary, we analyzed the potential effect of leptin signaling on BMP9-induced osteogenic differentiation of MSCs. Our results showed that exogenous leptin potentiated BMP9-induced osteogenic differentiation of MSCs, while inhibiting BMP9-induced adipogenic differentiation. We further found that BMP9 induced the expression of leptin and LepR in MSCs, while exogenous leptin reciprocally induced BMP9 expression in less differentiated MSCs. Mechanistically, we demonstrated that a JAK1/2 inhibitor effectively blunted the leptin-potentiated osteogenic differentiation induced by BMP9. Taken together, these findings strongly suggest that leptin may potentiate BMP9-induced osteogenesis by cross-regulating BMP9 signaling through the JAK/STAT signaling pathway in MSCs. Therefore, a combined use of BMP9 and leptin may be explored as a novel approach to enhancing efficacious bone regeneration and fracture healing.
Disclaimer
Funding sources were not involved in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; and in the decision to submit the article for publication.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The reported work was supported, in part, by research grants from the National Institutes of Health (CA226303 to T.-C.H.), the United States Department of Defense (OR130096 to J.M.W.), and the Scoliosis Research Society (T.-C.H. and M.J.L.). W.W. was supported by the Medical Scientist Training Program of the National Institutes of Health (T32 GM007281). This project was also supported, in part, by The University of Chicago Cancer Center Support Grant (P30CA014599) and the National Center for Advancing Translational Sciences of the National Institutes of Health through grant number UL1 TR000430. T.-C.H. was also supported by the Mabel Green Myers Research Endowment Fund and The University of Chicago Orthopedics Alumni Fund.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.