
Abstract
Heart transplantation is the final life-saving therapeutic strategy for many end-stage heart diseases. Long-term immunosuppressive regimens are needed to prevent allograft rejection. Mesenchymal stromal cells (MSCs) have been shown as immunomodulatory therapy for organ transplantation. However, the effect of dose and timing of MSC treatment on heart transplantation has not yet been examined. In this study, we infused three doses (1 × 106, 2 × 106, or 5 × 106 cells) of human MSCs (hMSCs) to the recipient BALB/c mice before (7 days or 24 h) or after (24 h) receiving C57BL/6 cardiac transplants. We found that infusion of high dose hMSCs (5 × 106) at 24 h post-transplantation significantly prolonged the survival time of cardiac grafts. To delineate the underlying mechanism, grafts, spleens, and draining lymph nodes were harvested for analysis. Dose-dependent effect of hMSC treatment was shown in: (1) alleviation of International Society of Heart and Lung Transplantation (ISHLT) score in grafts; (2) reduction of the population of CD4+ and CD8+ T cells; (3) increase of regulatory T (Treg) cells; (4) and decrease of serum levels of inflammatory cytokines and donor-specific antibodies. Taken together, we showed timing critical and dose-dependent immunomodulatory effects of hMSC treatment against acute allograft rejection in a mouse model of heart transplantation.
Introduction
Heart failure is a collection of complex clinical symptoms characterized by the reduced ability of the heart to pump an adequate supply of blood to the body [1]. It is a global pandemic that continues to increase in both incidence and prevalence despite the advances in the pharmacologic treatment or surgical approaches [2]. Cardiac transplantation is often the final therapy for the patients of end-stage heart failure if medications or surgeries fail to improve their symptoms [3]. Although heart transplantation is capable to prolong the life span of patients, long-term use of immunosuppressive drugs is needed to avoid cardiac allograft rejection [4]. The resulting side effects, such as infection, anemia, renal toxicity, metabolic diseases, and malignancy, are not uncommon, and novel strategies are still in high demand [5].
To achieve allograft tolerance and avoid long-term immunosuppression, cell-based therapies have been proposed as innovative approaches in solid organ transplantation [6]. Administration of cells with immunoregulatory properties to transplant recipients could balance the effector and regulatory pathways to control the immune responses to the allograft. Mesenchymal stromal cells (MSCs) are multipotent cells which could be isolated from multiple tissues, such as umbilical cord, bone marrow, and adipose tissue [7]. Owing to the regenerative and immunomodulatory capacities, MSCs have been used as a potential approach against allograft rejection in solid organ transplantation [8 –10].
In rodent models, MSCs combined with immunosuppressive drugs, such as sirolimus, cyclosporine A, and mycophenolate mofetil, synergistically prolong allograft survival [11 –13]. Furthermore, human MSCs (hMSCs) have been found beneficial for immunosuppression in clinical transplant therapeutic regimens [14,15]. In clinical trials, the doses of infused MSCs have ranged from 0.4 × 106 to 20 × 106/kg body weight [16 –18]. The dose, timing, and frequency of MSC administration are suggested to be critical for the allograft survival [19]. However, little studies specifically established the dose- or time-dependent responses of MSC treatment on heart transplantation.
Mediators of innate and adaptive immunity play important roles in different forms of the rejection of the cardiac transplant, including acute rejection, cellular rejection, antibody-mediated rejection, and chronic injection [20 –22]. Multiple immunoregulatory mechanisms by which MSCs induce long-term allograft survival have been uncovered [23,24]. MSCs could inhibit (1) the proliferation, activation, and function of effector T cells; (2) induction of donor-specific regulatory T cells (Tregs); and alter the functions of natural killer cells, dendritic cells, and B cells [25 –28]. The purpose of this study is to investigate the dose- and time-dependent effects of hMSC treatment in a mouse model of heart transplantation. Moreover, the immunomodulating mechanisms by which the optimal hMSC treatment regulates allograft tolerance were examined.
Materials and Methods
Ethics Statement
This study was carried out in strict accordance with the guidelines established by the Institutional Animal Care and Use Committee of the Chinese Association for Laboratory Animal Sciences. All protocols involving animals were approved by the First Affiliated Hospital of Sun Yat-sen University Committee on the Use and Care of Animals (No. 2013-35). All surgery was performed under anesthesia using sodium pentobarbital, and all efforts were made to minimize any suffering experienced by the animals.
hMSC isolation and characterization
Human bone marrow samples were aspirated from a young healthy donor after obtaining the informed consent and ethical approval by the Institutional Clinical Ethics Committee of the First Affiliated Hospital, Sun Yat-sen University, in compliance with the provisions of the current Declaration of Helsinki Principles and Good Clinical Practice guidelines. The mononuclear cells of the bone marrow were collected and successfully cultured until the sixth passage. At the fifth passage, cells were characterized by surface marker profiling using the following markers: CD29, CD34, CD44, CD45, CD73, CD105, and CD166 (antibodies were from BD Biosciences). Cells were also assessed by osteogenic and adipocytic differentiation assays, as previously described [29,30]. Karyotyping analysis was performed for the MSCs at passage 6, and no chromosomal aberration was observed (Supplementary Fig. S1). All reagents for these assays were provided by the Key Laboratory for Stem Cells and Tissue Engineering of the Education Ministry of China, Sun Yat-sen University (Guangzhou, China).
Mice
Male C57BL/6 (H-2b) and BALB/c (H-2d; 8–12-weeks old) mice were purchased from the Laboratory Animal Center of Guangdong Province (Guangzhou, China) and were used as graft donors and recipients, respectively. Care and handling of animals were carried out in accordance with the guidelines provided in the Guide for the Care and Use of Laboratory Animals published by the U.S. Department of Health and Human Services.
Heart transplantation and hMSC treatment
Vascularized heterotopic heart transplants from C57BL/6 donors to BALB/c recipients were performed with anastomosis to the vessels of the neck using a nonsuture cuff technique, as described previously [31]. The recipient mice were intravenously administered different doses of hMSCs (1 × 106, 2 × 106, and 5 × 106 cells per mouse) at different time points (7 days, 24 h before transplantation, or 24 h after transplantation). The injection volume of hMSCs was 0.5 mL. The control group received heart allograft transplantation and phosphate-buffered saline instead of hMSCs. Six to eight BALB/c mice received heart transplantation, and allograft survival was monitored twice per day. Rejection was defined as the loss of palpable cardiac contractions. Heart transplantation from BALB/c donors to BALB/c recipients was performed as the syngeneic control. For mechanism study, additional nine mice received heart transplantation in each group, among whom three mice were, respectively, euthanized at days 7, 14, and 21 after transplantation to obtain heart graft, blood, spleen, and draining lymph nodes (dLNs) of the recipient.
Histological evaluation of rejection
Heart grafts were removed on days 7, 14, and 21 post-transplantation. Paraffin-embedded transventricular tissue sections (5-μm thick) were stained with hematoxylin and eosin (H&E). The rejection was graded in a blinded manner with experienced pathologist based on the extent of lymphocytic infiltration and the anatomical localization of leukocytes according to the International Society of Heart and Lung Transplantation (ISHLT) standards [32,33]. Briefly, the heart tissue was scored as follows: 0 = no damage; 1 (mild) = evidence of interstitial edema and focal necrosis; 2 (moderate) = the graft displayed diffuse myocardial cell swelling and necrosis; 3 (severe) = necrosis with the presence of contraction bands and a neutrophil infiltrate; and 4 (highly severe) = widespread necrosis with the presence of contraction bands, neutrophil infiltrate, and hemorrhage.
Experiments detected by flow cytometry
Antibodies and kits: FITC anti-mouse CD4 (GK1.5), PerCP-Cy5 anti-mouse CD8α (53–6.7), APC anti-mouse CD25 (PC61.5), FITC anti-mouse IgG (Cat No. 11-4011-85), PE anti-mouse IgM (Cat No. 12-5790-81), PE anti-mouse IgG (Cat No. 12-4010-82), FITC anti-mouse CD3 (17A2), PE-Cy7 anti-mouse CD25 (3C7), APC anti-mouse CD4 (GK1.5), PE-Cy7 anti-mouse interferon gamma (IFN-γ) (XMG1.2), 7–AAD, and corresponding conjugated isotype control antibodies were purchased from eBioscience. Murine Treg cells were labeled using the Mouse Regulatory T cell Staining Kit from eBioscience (Cat No. 88-8111).
Splenocytes were isolated from mice and cultured with or without hMSCs in the presence of anti-CD3 mAb (1 μg/mL) and anti-CD28 mAb (1 μg/mL) for 24 or 48 h. For the 24 h cultured cells, the MSC to T cell ratio was 1:10, and the expression of activated marker CD25 on CD3+ CD4+ T cells was detected. For 48 h cultured cells, Leukocyte Activation Cocktail (2 μL/mL, BD) was added 5 h before the end of the culture period, and cells were stained to detect anti-CD3 and cytoplasmic IFN-γ.
Splenocytes were isolated from mice with or without hMSC treatment 7 days after heart transplantation and were labeled with CFSE (carboxyfluorescein diacetate succinimidyl ester, Sigma-Aldrich). CFSE-labeled splenocytes (50 × 106) were plated on a Petri dish for 45 min at 37°C to enrich lymphocytes. CFSE-labeled splenocytes (2 × 105) were then cultured with anti-CD3 mAb (1 μg/mL) and anti-CD28 mAb (1 μg/mL) at 37°C for 3, 5, and 7 days, and then proliferative cells were detected.
Mixed lymphocyte reaction
Naive mice and heart allograft recipient mice with or without (w/o) hMSCs treatment were euthanized on days 7, 14, and 21. Spleens were processed into single-cell suspensions, labeled with CFSE, and resuspended in DMEM; then 50 × 106 CFSE labeled splenocytes were plated on a Petri dish for 45 min at 37°C to enrich lymphocytes. After 45 min, nonadherent cells were collected, washed, and incubated (1 × 105 cells) with irradiated (2,000 cGy, 1 × 105 cells) naive BALB/c splenocytes in 96-well U-bottom titer plates in mixed lymphocyte reaction (MLR) medium. After 5 days, cells were stained with fluorochrome-labeled Abs against mouse CD4, CD8, and 7AAD and then analyzed by flow cytometry.
Cytometric bead array, donor-specific antibodies and antibody against hMSCs
Serum from recipient mice was isolated on days 7, 14, and 21 post-transplantation. To detect the levels of interleukin (IL)-2, IL-4, IL-6, IL-10, IL-17, IFN-γ, and tumor necrosis factor alpha (TNF-α) in the serum, cytometric bead array (CBA) was performed using commercially available kit (Shanghai Yikesai Bioproduct, Shanghai, China) according to manufacturer's instructions. To analyze the levels of donor-specific antibodies (DSA) in the serum, donor splenocytes from naive C57BL/6 mice (106 cells/100 μL) were incubated with 10 μL of recipient mouse serum at 4°C for 30 min and washed and then stained with FITC anti-mouse IgG or PE anti-mouse IgM antibodies at 4°C for 30 min. Serum from BALB/c to BALB/c sham group was used as a negative control.
Seven days after hMSC infusion, naive mice or heart allograft recipient mice were euthanized and serum was prepared. hMSCs (106 cells, 100 μL) were incubated with 100 μL of naive or recipient mouse serum at 37°C for 1 h and washed, blocked with anti-Mouse CD16/CD32 (2.4G2), and then were stained with PE anti-mouse IgG antibody at 4°C for 30 min. The incubated hMSCs were analyzed on a flow cytometer (BD FACSCalibur), and data were analyzed using FlowJo 7.5 software.
Statistical analysis
Survival curves and graphs of histological data were obtained using GraphPad Prism® (GraphPad software, La Jolla, CA). The survival curves were analyzed using the log-rank test. Data from the flow cytometry and CBA experiments were analyzed using one-way analysis of variance and expressed as mean ± standard deviation. A value of P < 0.05 was considered as statistically significant and P < 0.01 as highly statistically significant.
Results
High dose and late timing of hMSC treatment improve cardiac graft survival
To establish the optimal hMSC regimen, we monitored the survival of the cardiac graft in a mouse model of heart transplantation. After receiving C57BL/6 heart transplants for 24 h, the recipient BALB/c mice were intravenously infused with different doses of hMSCs (1 × 106, 2 × 106, and 5 × 106 cells/mouse). The mean survival time of cardiac graft in all hMSC treatment groups was increased (control group, 7.5 days; 1 × 106 MSC group, 9.5 days; 2 × 106 MSC group, 12.1 days; 5 × 106 MSC group, 20.8 days). The survival of cardiac graft among groups was compared by the Kaplan–Meier method (Fig. 1A). High dose of hMSCs (5 × 106 cells) significantly prolonged the survival time of cardiac graft compared to the control group (P < 0.001) or to the low dose of hMSC (2 × 106 cells) group (P < 0.01). Furthermore, we examined the time-dependent effect of high-dose hMSC (5 × 106 cells) treatment. The recipient BALB/c mice were intravenously infused with hMSCs (5 × 106 cells) before (7 days or 24 h) and after (24 h) receiving C57BL/6 heart transplants. The survival of cardiac graft among groups was compared by the Kaplan–Meier method (Fig. 1B). hMSCs infused at 24 h post-transplantation significantly prolonged the survival of cardiac graft compared to the other groups (P < 0.001).

Cardiac graft survival after hMSC treatment.
Dose-dependent effects of hMSC treatment on inflammatory cell infiltration in the grafts
Post-heart transplant, mediators of innate and adaptive immunity influence the possible development of rejection [34]. To evaluate inflammatory infiltration following transplantation, the ISHLT score of the grafts was assessed at days 7, 14, and 21 after heart transplantation (Fig. 2). The myocardium of the grafts from the control group (ISHLT Grade 3–4) showed extensive damage and infiltration of inflammatory cells. On the contrary, the grafts from the hMSC treatment groups showed a significant dose-dependent reduction of the ISHLT scores. At day 7, high dose (5 × 106 cells) of hMSC treatment presented the lowest level of lymphocytic infiltration and fewest changes in myocardial structural integrity among all groups (ISHLT Grade 1–2) (P < 0.05). At days 14 and 21, graft rejection in the high dose (5 × 106 cells) of hMSC treatment group had increased (ISHLT Grade 3–4). Results indicated that hMSC treatment significantly protects the cardiac grafts from acute rejection in a dose-dependent manner.
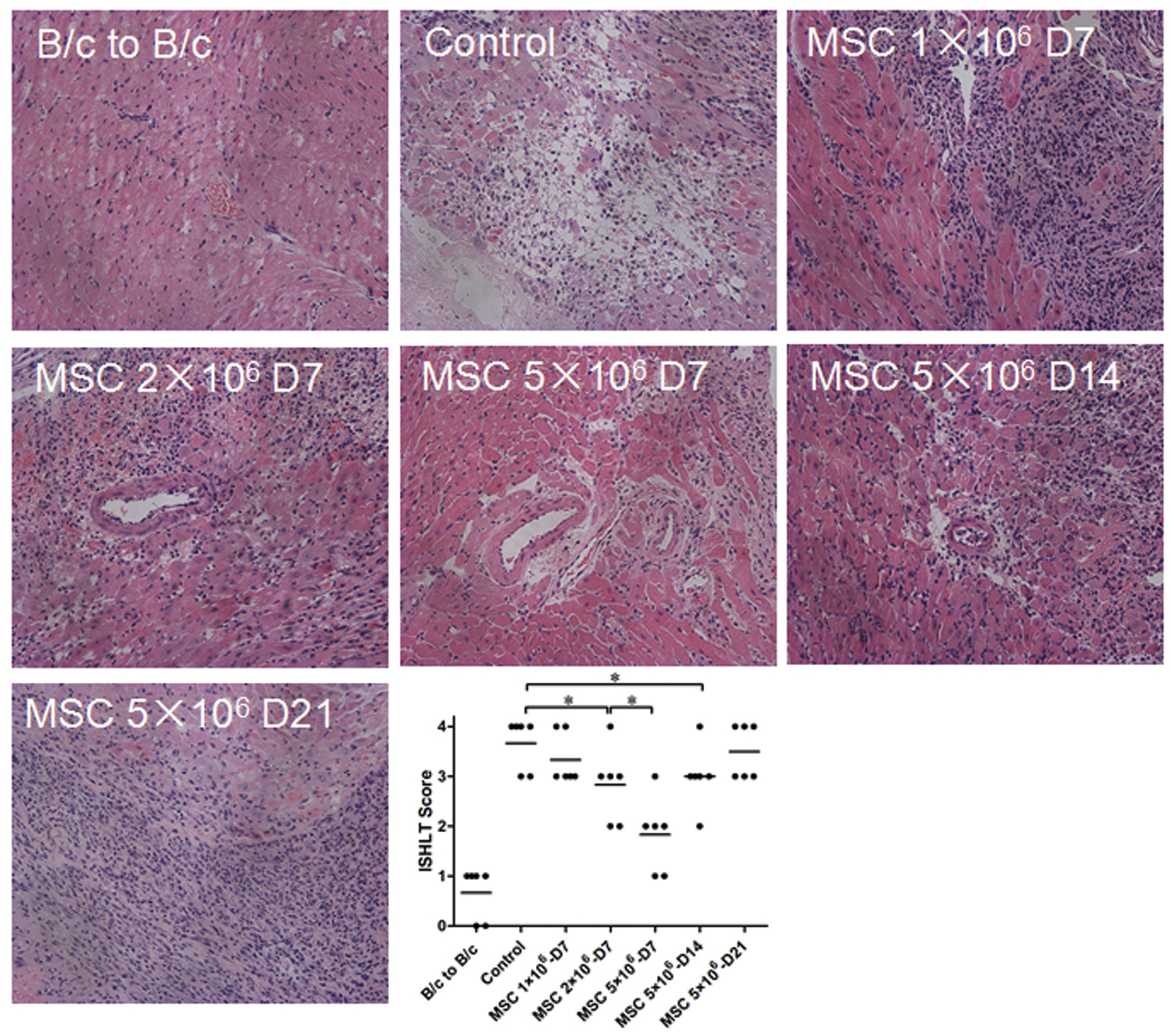
Pathological changes and the ISHLT scores of the cardiac grafts. The recipient BALB/c (H-2d) mice were intravenously infused with hMSCs at a dose of 1, 2, or 5 × 106 cells after 24 h of C57BL/6 (H-2b) heart transplantation. Grafts were harvested at days 7, 14, and 21 after heart transplantation and assessed using H&E staining (100 × ). Representative photographs of each group were shown. Following pathological observations, the ISHLT score was calculated and shown in the scatter plot (*P < 0.05; n = 6 for each group). Bars indicate mean of values. H&E, hematoxylin and eosin; ISHLT, International Society of Heart and Lung Transplantation. Color images are available online.
Dose-dependent effects of hMSC treatment on the frequencies of CD4+ and CD8+ T cells in blood, spleen, and dLNs of the recipient mice
Since T cells have been widely shown to contribute rejection in organ transplantation [35], we isolated lymphocytes in the blood, spleen, and dLNs of the recipient mice at days 7, 14, and 21 after heart transplantation. Mononuclear cells were gated and separated from debris. T cell populations (CD4+ helper and CD8+ cytotoxic T cells) were further analyzed by flow cytometry (Fig. 3). At day 7 post-transplantation, compared with the B/c to B/c sham group, the heart transplantation control group showed remarkable increase in the frequency of CD8+ T cells (2.3-fold in peripheral blood mononuclear cell [PBMC], 1.8-fold in spleen, and 1.5-fold in dLNs) and modest increase in CD4+ T cells (1.3-fold in PBMC, 1.4-fold in spleen, and 1.4-fold in dLNs). Meanwhile, the treatment of hMSCs reduced the frequencies in CD4+ and CD8+ T cells in a dose-dependent manner. Notably, hMSCs at high dose (5 × 106 cells) were most capable in the suppressing effect than low dose groups. It was of note that the percentages of CD8+ T cells in PBMC, spleen, and dLNs remained lower than those of control group on day 21 in the high-dose hMSC group (5 × 106 cells). While the percentage of CD4+ T cells in PBMC in the high-dose hMSC group rebound to the control group level on day 14, similar phenomena were observed on day 21 in the spleen and dLNs. Collectively, the data suggested that hMSCs at high dose (5 × 106 cells) are superior to low dose groups in reducing T cell frequencies in circulation and lymphoid organs.
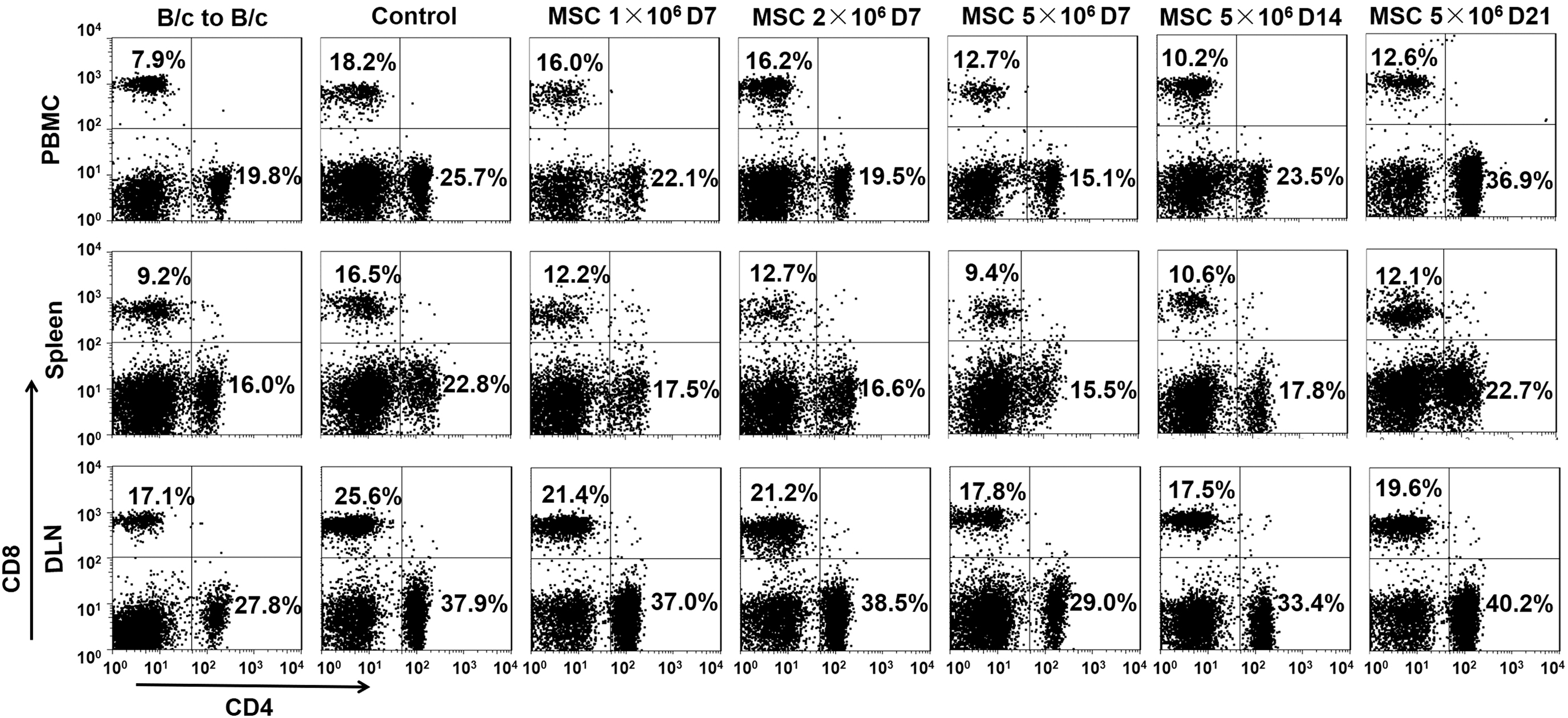
Distribution of CD4+ and CD8+ T cell in the blood, spleen, and dLN of the recipient mice receiving hMSC treatment and the control mice. The recipient BALB/c (H-2d) mice were intravenously infused with hMSCs at a dose of 1, 2, or 5 × 106 cells after 24 h of C57BL/6 (H-2b) heart transplantation. Blood, spleen, and dLN were harvested at days 7, 14, and 21 after heart transplantation and assessed for the distribution of CD4+ and CD8+ T cells using flow cytometry. Representative dot plots of three independent experiments were shown. dLN, draining lymph node.
Time-dependent effects of high dose hMSC treatment on the Tregs frequency in spleen and dLNs of the recipient mice
T regulatory cells (Tregs) play important roles in maintaining self-tolerance and immune homeostasis by suppressing subset of T cells with self-antigen reactivity [36]. MSCs have been shown to expand the circulating population of Tregs in a mouse model heart transplantation [37]. Thus we examined the effect of hMSC treatment on the frequency of Tregs in spleen and dLNs of the recipient mice at days 7, 14, and 21 after heart transplantation. Mononuclear cells were gated and separated from debris. Tregs were identified as CD4+ and Foxp3+ cells using flow cytometry (Fig. 4). There was no difference in the Tregs frequency between B/c to B/c group and heart transplantation control group at day 7. The dose–response in the effect of hMSC treatment on modulating Tregs frequency was observed at day 7. Tregs frequency of high dose group (5 × 106 cells) was higher than the low dose group (1 × 106 cells) in spleen at day 7, and at day 14, the percentage also increased in the dLNs. The data suggested that hMSCs at high dose (5 × 106 cells) might exert the immunosuppressive effect by inducing long-term and time-dependent expansion of Tregs in the lymphoid organs, spleen, and dLNs.

The population of CD4+ Foxp3+ T (Treg) cells in the spleen and dLNs of the recipient mice receiving hMSC treatment and the control mice. The recipient BALB/c (H-2d) mice were intravenously infused with hMSCs at a dose of 1, 2, or 5 × 106 cells after 24 h of C57BL/6 (H-2b) heart transplantation. Spleen and dLN were harvested at days 7, 14, and 21 after heart transplantation and assessed for the population of CD4+ Foxp3+ Treg cells using flow cytometry.
Dose-dependent effects of hMSC treatment on the serum levels of cytokines of the recipient mice
To evaluate the immune status systematically, we collected the serum from recipient mice at days 7, 14, and 21 after heart transplantation and performed cytokine profiling using CBA system and flow cytometry (Fig. 5). At day 7, compared with the B/c to B/c sham group, the heart transplantation control group showed significantly high levels of lymphocyte pro-proliferating cytokine IL-2 and pro-inflammatory cytokine IFN-γ, which could be dose dependently reduced in hMSC treatment. On the contrary, the levels of anti-inflammatory cytokine IL-10 was dose dependently induced in hMSC treatment groups. At days 14 and 21, the levels of IL-2 and IFN-γ were increased, whereas the levels of IL-10 were reduced over time. The data were consistent with the effect of hMSC treatment on the T cell subsets (Figs. 3 and 4). In addition, we found no significant changes in the serum levels of IL-4, IL-6, IL-17, or TNF-α among the control and hMSC treatment groups (data not shown).
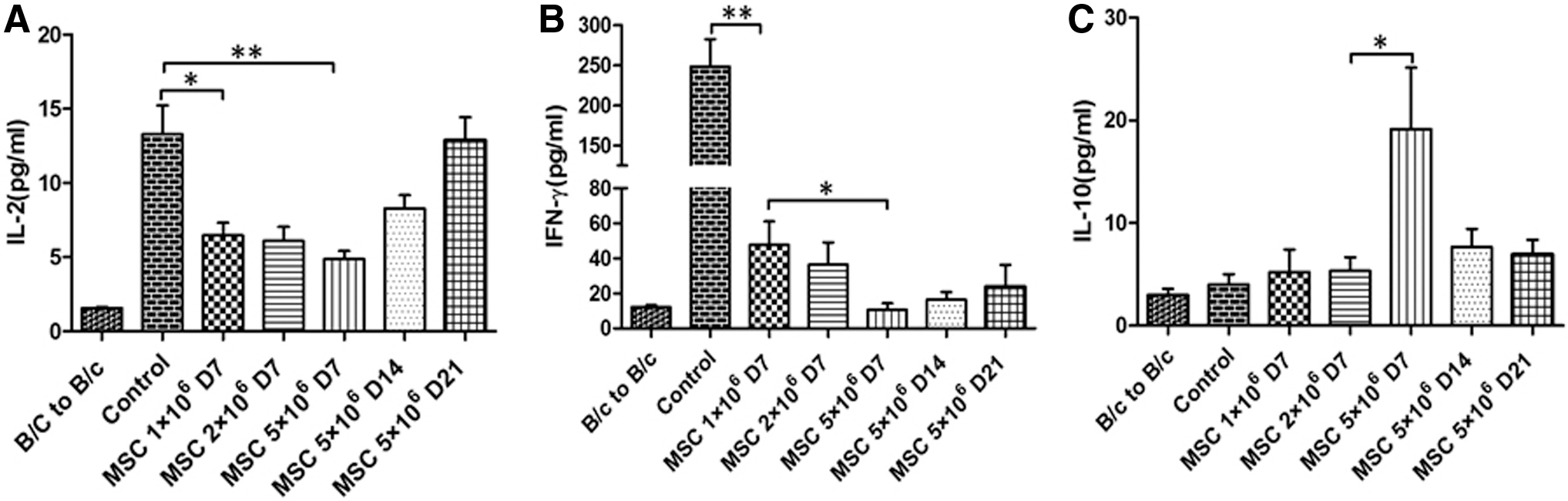
Serum levels of cytokines in the recipient mice receiving hMSC treatment and the control mice. The recipient BALB/c (H-2d) mice were intravenously infused with hMSCs at a dose of 1, 2, or 5 × 106 cells after 24 h of C57BL/6 (H-2b) heart transplantation. Blood was collected at days 7, 14, and 21 after heart transplantation, and serum was separated and assessed using cytometric bead array for the levels of IL-2
Dose-dependent effects of hMSC treatment on the serum levels of DSA of the recipient mice
Along with the T cell mediated cardiac allograft rejection, antibody-mediated rejection is also detrimental and mainly contributed to the generation of DSA [38]. We therefore collected the serum from recipient mice at days 7, 14, and 21 after heart transplantation and incubated with donor splenocytes isolated from naive C57BL/6 donor mice. DSA were then measured using anti-IgG or -IgM antibodies and flow cytometry (Fig. 6). At day 7, compared to the B/c to B/c sham mice, DSA were remarkably elicited in the heart transplantation control group. Treatment with hMSCs showed dose-dependent effect in reducing DSA. At days 14 and 21, the reduced serum levels of DSA were reversed over time.

Serum levels of donor-specific antibodies in the recipient mice receiving hMSC treatment and the control mice. The recipient BALB/c (H-2d) mice were intravenously infused with hMSCs at a dose of 1, 2, or 5 × 106 cells after 24 h of C57BL/6 (H-2b) heart transplantation. Blood was collected at days 7, 14, and 21 after heart transplantation, and serum was separated and assessed using flow cytometry for the levels of IgG
Discussion
As the field of stem cell therapy continues to expand, clinical trials of various approaches for heart disease continue to indicate the dose-dependent effect of MSCs in clinical responses [39 –41]. However, the comparison of optimal timing and doses of MSC treatment in heart transplantation in either animal models or clinical trials have not yet been reported. In this study, we sought to examine whether there are dose- and time-dependent effects of MSC treatment on graft rejection and immune activation in a mouse model of heart transplantation. We found the best efficacy in prolonging allograft survival in the group of intravenous infusion of high dose (5 × 106 cells/mouse) of hMSCs at 24 h after heart transplantation. The underlying mechanism might be through suppression of the frequencies of effector T cells, serum levels of pro-inflammatory cytokines, and DSA. In reciprocal, the frequencies of Tregs and serum levels of anti-inflammatory cytokine were upregulated.
In agreement with studies using animal models of stroke and lung injury [42 –44], we also demonstrated that the timing and dose of MSC administration are critical for their beneficial effect. Compared to 24 h before or 7 days before heart transplantation, infusion of hMSCs at 24 h after heart transplantation was more efficient in the protection of cardiac allograft survival. However, some studies showed that administration of MSCs at 4 and 7 days before transplantation could achieve longer graft survival time [13,37]. The discrepancy among these results might be related to the differences in transplant models or the sources of MSCs. Regarding the dose of MSCs, comparative analysis of preclinical murine data suggested intravenous administration of 50 million cells/kg (106 cells per mouse weighing 20 g) typically [19,45]. Therefore, in an attempt to address the maximum no adverse effect dose of MSC treatment, we compared the effects of three escalating doses (106, 2 × 106, and 5 × 106 cells for each mouse) on cardiac allograft survival. The findings indicated that, without any obvious side effects, the dose of MSCs is critical on the survival of allografts, and high dose of MSCs was more effective in inhibition of acute rejection. A higher effective dose of MSCs could be further determined in the future with the concern that life-threatening side effects, such as embolism, might occur.
Despite the remarkable efficacy of high dose (5 × 106 cells) MSC treatment in preventing cardiac allograft from acute rejection and lymphocyte infiltration at day 7 after heart transplantation, the ISHLT score continued to increase over time and reach to a similar score of the control group at day 21 post-transplantation. This finding indicated that single dosing of MSCs might not be sufficient for long-term prevention of chronic rejection. The beneficial effect of multiple dosing of MSCs has been shown in animal models of diabetes and myocardial infarction [46,47]. Further investigation would be needed to compare the effect of dosing frequency on the efficacy of MSC treatment in long-term protection of allograft survival.
T cells are recognized as critical cellular mediators in solid organ transplantation [35]. The inhibitory effects of MSCs on CD4+, CD8+, and even memory antigen-specific T cells have been reported [48 –51]. In the context of heart transplantation, we herein showed consistent results that the frequencies of CD4+ and CD8+ T cells were increased in circulation and lymphoid organs, including spleen and dLNs, which were further reduced by MSC treatment in a dose-dependent manner. Not only by inhibiting the frequencies, but MSC treatment also suppressed the function of CD4+ and CD8+ T cells by reducing systemic levels of IL-2 and IFN-γ. Since we found that MSC treatment did not affect the levels of IL-4, IL-6, IL-17, or TNF-α (data not shown), it is somewhat suggested that MSC treatment might suppress Th1 responses better than Th2 in our model. However, according to other report showing that MSCs could also suppress Th2 cells [52], the specific effects of MSC treatment on Th1 and Th2 cells in heart transplantation need to be dissected in more detail. In this study, when continuously cultured with T cells in vitro, hMSCs significantly inhibited CD4+ T cell activation (Supplementary Fig. S2A) and IFN-γ secretion of CD3+ T cells (Supplementary Fig. S2B). However, this therapeutic advantage cannot be sustained for a long time when hMSCs were intravenously transfused in vivo as shown in Supplementary Fig. S3, and also we did not find statistically significant suppression of donor specific immune reaction by MLR (Supplementary Fig. S4). One of the reasons might be that most intravenous MSCs were trapped and eliminated in the lung, and too few MSCs reach to lymphoid organ or tissues to induce strong and persistent inhibitory effect. This leads to one of the important issues regarding translational research of MSCs, that is, improvement of administration methods. Second, we also found that the percentages of Tregs increased in the dLNs more significantly and maintained for a longer time. Local immune microenvironment and immunosuppression may play a critical role in the hMSC treatment protocol. Further studies are needed to enhance therapeutic effect of hMSCs.
Opposite to the detrimental roles of effector T cells, Tregs are beneficial for the immune tolerance of allograft, and the transfer of Tregs even becomes an attractive cell-based therapy to improve the outcome of transplantation in either preclinical models or clinical trials [53]. Therefore, it is critical to see whether Tregs are involved in the immunosuppressive abilities of MSCs in our model. We found that hMSC treatment increased the frequencies of Tregs over time but the dose–response was not statistically significant. Further investigation will be needed to establish the role of Tregs. Nevertheless, it has been shown that MSC infusion could generate CD4+CD25+Foxp3+ Tregs in a semiallogeneic heart transplant model [37]. In addition, the levels of anti-inflammatory cytokine IL-10 were transiently upregulated in the high dose (5 × 106 cells) of MSC treatment group, which were somewhat in agreement with previously published data that suggested the importance of IL-10 in MSC-mediated immunosuppression [54,55]. MSCs could also increase CD5+ regulatory B cells that produced IL-10 [56]. The function of B cells was suppressed by MSC treatment since we observed a dose–response in DSA generation. Besides, recent research indicated that macrophages and monocytes play a critical role in the immunomodulatory effect of MSC, mainly through indoleamine 2,3-dioxygenase (IDO) production and Foxp3+ Tregs induction [57,58]. Taken together, we, therefore, speculated that higher dose or repeated dosing at optimal timing of MSC treatment might be able to induce Tregs and IL-10 more efficiently to promote long-term graft protection.
HMSCs were used in this study, since xenogeneic mouse models may provide referable preclinical data of hMSC efficacy, which is of importance for clinical trial design. Accumulating evidence showed that hMSCs could effectively suppress immune responses in immunocompetent mice, but hMSCs also generated immune response in mice. As shown in Supplementary Fig. S5, anti-hMSC antibody was significantly induced 7 days after hMSC infusion in mice. This phenomenon maybe can explain the lack of effect of hMSC when given pretransplant, due to their possible reduction by the anti-hMSC antibodies the mice developed during time. The consequence of anti-hMSC antibody needs to be further investigated.
In summary, our data provide a parallel comparison of the effects of different timing and dose of hMSC treatment on cardiac allograft rejection in a mouse model of heart transplantation. The timing and dose reported here might not be readily translated to equivalency in human studies. Nevertheless, our results provide further evidence in heart transplantation that the efficacy of hMSC treatment on protecting allograft rejection would be largely determined by the dose and timing of administration.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by the National Natural Science Foundation of China (81870511, 81670680, 81700655, 81970109, 81701582), Science and Technology Planning Project of Guangdong Province, China (2015B020226002, 2017A020215012), Guangdong Natural Science Foundation (2018A030313016), Key Scientific and Technological Program of Guangzhou City (201803040011), Pearl River S&T Nova Program of Guangzhou (201906010095), and Guangdong Provincial Key Laboratory on Organ Donation and Transplant Immunology (2013A061401007, 2017B030314018).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.