
Abstract
Donor mesenchymal stem cells (MSCs) have been documented in fetal and maternal circulations after plain intra-amniotic injection, with diverse therapeutic effects. We sought to determine the pathway of this unique cell kinetic route. Rat fetuses (n = 226) were divided into two groups based on the content of intra-amniotic injections performed on gestational day 17 (E17): either a concentrated suspension of luciferase-labeled syngeneic amniotic fluid-derived MSCs (afMSCs; n = 111), or acellular luciferase (n = 115). Samples from placenta, chorion, amnion, amniotic fluid, stomach fluid, peripheral blood, and umbilical cord were procured at five daily time points thereafter until term (E18–22) for luminometry. In addition, 53 sets of fresh gestational membranes (chorion/amnion combined) from nonmanipulated term fetuses were secured to transwell inserts for in vitro analysis of MSC migration using luciferase-labeled afMSCs. Statistical analyses included the Mann-Whitney U-test, Wald test, nonlinear regression modeling, and Fisher's exact test. In vivo, luciferase activity was observed in the amnion, chorion, and placenta of fetuses receiving cells, but not in those receiving acellular luciferase (P < 0.001). There was a consistent nonlinear age-dependent relationship of luciferase activity between the amnion, chorion, and placenta following a parabolic bimodal pattern characterized by significantly higher early preterm (E18) and late-term (E22) activities (P < 0.001), with no differences between E21 and E22 (P = 0.12). In vitro, the presence of cells was documented by luminometry in 21/53 (39.6%) of the assays, in suspension and/or attached to the plastic substrate, and within all screened gestational membrane sets, irrespective of stimuli with collagen coating or fetal bovine serum. We conclude that, after intra-amniotic injection, donor MSCs undergo controlled cell routing, as opposed to passive clearance. Transgestational membrane transport appears to constitute the path for donor cells to reach the placenta, a known gateway to the fetal circulation, significantly expanding the potential applications of transamniotic stem cell therapy.
Introduction
Mesenchymal stem cells (MSCs) have long been isolated from the mammalian placenta, chorion, and amnion [1 –4]. Yet, to this day, the spatiotemporal mapping and kinetic interchange between these and other stem cell pools throughout gestation, along with the mechanisms that govern such movements, remain incompletely understood. The emerging concept of transamniotic stem cell therapy (TRASCET), consisting of concentrated intra-amniotic injections of select fetal-derived MSCs (typically from amniotic fluid or placenta), is based on harnessing some of these movements so as to magnify the biological role of these cells in fetal tissue repair, targeting therapeutic benefit through a minimally invasive approach to targeted cell delivery to the fetus [5 –14]. Previous experimental work in both healthy animals and disease models has shown that donor MSCs quickly reach the placenta after plain intra-amniotic delivery, which then appear to serve as the gateway for hematogenous trafficking of these cells through routes that include the umbilical cord and fetal bone marrow, explaining many of the intriguing effects of TRASCET documented so far [15 –17]. In this study, we sought to test our hypothesis that the gestational membranes constitute the path through which donor MSCs can reach the placenta after plain intra-amniotic injection.
Materials and Methods
This study was approved by the Boston Children's Hospital's Institutional Animal Care and Use Committee under protocol number 15-07-2951R.
Donor MSC processing and labeling
Donor cells consisted of previously banked amniotic fluid-derived MSCs (afMSCs) procured from healthy syngeneic Lewis rat dams on gestational day 21 (E21; term = 21–22 days), isolated and expanded based on methods as previously described by our group [18]. Fluorescence-activated cell sorting analysis was used to confirm their mesenchymal progenitor identity with unconjugated primary mouse monoclonal antibodies previously validated for use in rats, namely CD29; CD44; CD45; CD73; and CD90 (all from Becton Dickinson, Franklin Lakes, NJ), using the Vantage SE cell sorter (Becton Dickinson). Nonspecific cell staining was excluded using mouse isotype immunoglobulin controls.
Proliferating donor afMSCs were labeled with the Cignal™ Lenti Reporter firefly luciferase reporter gene (Qiagen, Germantown, MD) based on methods as previously described [19,20]. Briefly, cells were transduced with puromycin-resistant Cignal Lenti Reporter lentivirus (multiplicity of infection = 50; Qiagen) in high-glucose Dulbecco's modified Eagle medium (DMEM) with
Intra-amniotic injections
Nineteen time-dated pregnant Sprague–Dawley dams (Charles River Laboratories, Inc., Wilmington, MA) were fed a normal diet ad libitum and housed individually under standard dark/light cycling conditions. All dams underwent intra-amniotic injections through surgery performed on E17. General anesthesia was induced and maintained with isoflurane (Abbott Laboratories, North Chicago, IL), chamber inhaled at 1.5%–3% in 100% oxygen, and dams were kept on a warmer throughout the procedure. A midline laparotomy was performed and the bicornuate uterus was eviscerated to allow for controlled, individual intra-amniotic injections under direct vision. A 33G non-coring needle (Hamilton Company, Reno, NV) on a 100 μL syringe (Hamilton Company) was introduced into each amniotic cavity with a viable fetus (n = 226) by the ventral aspect of the conceptus, with care to avoid injection into it, the placenta, or the umbilical cord (Fig. 1). Volume-matched injections of 50 μL of one of two different suspensions were performed. In one group of dams (n = 10; n = 111 fetuses), the suspension consisted of the luciferase-transduced syngeneic afMSCs at a concentration of 2 × 106 cells/mL in PBS. In the other group (n = 9; n = 115 fetuses), the suspension consisted of 30 ng of the recombinant luciferase protein (Promega) in PBS. In both groups, after the injections, the uterus was returned to the abdomen and the incision closed in two layers with 3-0 Prolene (Ethicon, Somerville, NJ) and 5-0 Vicryl (Ethicon) simple running sutures. Animals were allowed to recover with no additional manipulation other than powdered flagyl (Unichem Pharmaceuticals, Hasbrouck Heights, NJ) on the wound and analgesia with buprenorphine (Reckitt and Colman Pharmaceuticals, Richmond, VA) and Meloxicam (Norbrook, Inc., Lenexa, KS), as needed.

Intraoperative view of an intra-amniotic injection in a dam, using a 33G non-coring needle on a 100 μL syringe.
Specimen procurement and preparation
Dams were euthanized with chamber-inhaled carbon dioxide at five daily time points between the intra-amniotic injections and term (ie, at E18, E19, E20, E21, and E22) for specimen procurement, with 15–23 samples of each specimen per group per time point. A midline incision was made and the uterus was eviscerated. The myometrium was carefully incised over each amniotic cavity, keeping the gestational membranes intact, followed by removal of the fetuses “en-caul.” The amniotic fluid was procured by puncture of the gestational sac with a 31G needle. The gestational membranes were cut sharply from the placenta and umbilical cord, followed by mechanical separation of the amnion and chorion. Umbilical cords were preserved whole, including the vessels and Wharton's jelly. Placental samples were normalized to 200 ± 10 mg. Fetal peripheral blood was obtained either by aspiration from the external jugular vein with a 31G needle or decapitation and direct blood collection. At E20–22 only (due to size constraints at the other time points), fetal stomach fluid was procured by aspiration with a 31G needle and syringe under direct vision. All tissue and fluid samples were placed in 1.5 mL Eppendorf tubes, initially frozen on dry ice, and then stored at −80°C.
Days later, all samples were homogenized with a solution of 1 × lysis buffer (Promega). Amniotic fluid and stomach fluid were vortexed for 5 s, incubated for 20 min at room temperature, and hand spun briefly to remove debris. Sterile stainless-steel beads (Next Advance, Averill Park, NY) were added to placenta, umbilical cord, peripheral blood (clotted), amnion, and chorion samples to aid with cell lysis. Beaded samples were placed in an automated tissue homogenizer (Next Advance) for 5 min and then submitted to three freeze/thaw cycles, alternating exposure to liquid nitrogen and 50°C water bath to enhance cell lysis. Finally, all samples were centrifuged at 12,000 rpm for 5 min and the supernatant was isolated for luminescence analysis.
Transwell invasion and migration assay
Fresh, intact gestational membrane sets (chorion/amnion combined; n = 53) were procured from nonmanipulated healthy Sprague-Dawley fetuses at E21 and hermetically secured to transwell inserts with the amnion facing the inner well, after removal of a manufacturer-provided polycarbonate membrane (Corning, Inc.). A water-tight seal was ensured and rat luciferase-labeled afMSCs at aliquots of 4 × 105 cells in 100 μL DMEM were added to each insert (Fig. 2). Receiving wells were either uncoated and filled with DMEM only (n = 27) or precoated with rat tail collagen-I (Sigma-Aldrich) and filled with DMEM supplemented with 10% FBS (Life Technologies; n = 26) to asses for eventual chemotaxis impact on cell invasion and migration. After a 3-day incubation period, inserts and membrane sets were removed, and a water-tight seal was again confirmed. Receiving well contents were screened for the presence of suspended and/or attached cells by microplate luminometry. Attached cells were trypsinized from receiving wells after removal of receiving well media. In a subset of inserts (n = 5), the gestational membrane sets were also screened for the presence of cells.

Transwell insert fitted with a set of hermetically sealed gestational membranes (chorion and amnion).
Luminescence analysis
Specimens were screened for luciferase activity by microplate luminometry using a commercially available luciferase assay kit according to the manufacturer's instructions (Promega). Briefly, 20 μL of each sample lysate was loaded at room temperature into each well of a 96-well opaque white microplate (Corning, Inc.) and placed in an automated high-throughput FLUOstar Omega microplate reader (BMG Labtech, Cary, NC). For each well, 100 μL of an assay solution consisting of luciferin and ATP was injected and luminescence was measured in relative light units (RLUs). A known quantity of luciferase protein diluted with sterile water, cell culture lysis buffer (Promega), and 1 mg/mL bovine serum albumin (Thermo Fisher Scientific, Inc., Waltham, MA) was used as a positive control for each plate. Triplicate wells with 20 μL of 1 × cell culture lysis buffer were utilized as blanks for adjustment of background luminescence, also for each plate. The mean and standard deviation of these three wells were calculated and a value composed of the mean plus two standard deviations above the mean was subtracted from the RLUs of the specimens on the same plate. To create binary data, specimens demonstrating RLUs above this threshold were deemed positive, while those below this threshold were deemed negative.
Statistical analysis
Statistical analysis of the in vivo data was by the Mann-Whitney U-test, Fisher's exact test, and nonlinear regression modeling aimed at describing the time-based pattern of donor cell homing. In addition, logistic regression analysis was applied with a generalized estimated equations approach and the Wald test for comparing the rate of positive activity with term (day 22) as the reference group, considering sites in which there was significant activity detected. RLU data are presented as medians and interquartile ranges. Statistical analysis of the in vitro data was by the Fisher's exact test. In all analyses, statistical significance was set at two-tailed P < 0.05.
Sample sizes provided 80% statistical power for detecting differences in luminescence activity based on RLUs compared to controls and in relation to activity at term, and 80% statistical power for detecting differences in luminescence activity in receiving wells with versus without chemoattractant. Data analysis was performed using SPSS statistical software version 24.0 (IBM Corporation, Armonk, NY).
Results
Overall fetal survival after intra-amniotic injection was 92% (209/226), with no significant difference between the cell-injected (91%; 101/111) and the acellular luciferase-injected (94%; 108/115) groups (Fisher's exact test, P = 0.457). Luminescence activity was documented in the amnion, chorion, and placenta samples of fetuses receiving cells, but not in those receiving plain (acellular) luciferase (P < 0.001). There was a nonlinear gestational age-dependent variation in luciferase activity in the placenta and amnion, following a bimodal U-shape pattern, or parabola, characterized by significantly higher early preterm (E18) and late-term (E21/E22) activities (Fig. 3). In placenta samples, there was lower activity at E19 (P < 0.05), E20 (P < 0.02), and E21 (P < 0.005) compared to E22, but no significant difference in activity between E18 and E22 (P = 0.96). For both placenta and amnion, the vertex or bottom of the U-shape pattern was observed at E19. In chorion samples, there was significantly lower activity at E18, E19, and E20 compared with E22 (all P < 0.001), but no significant difference between E21 and E22 (P = 0.08). In amnion samples, there were no significant differences at the different time points compared with E22 (P > 0.10), except at E21, when luminescence was higher (P < 0.001). Overall, there was significantly higher activity in the amnion compared with both chorion and placenta (Wald test, P < 0.01). No significant differences were observed in median luminescence activity of stomach fluid (P > 0.20), amniotic fluid (P > 0.30), umbilical cord (P > 0.13), and peripheral blood (P > 0.19) samples between cell-injected and acellular luciferase-injected fetuses.
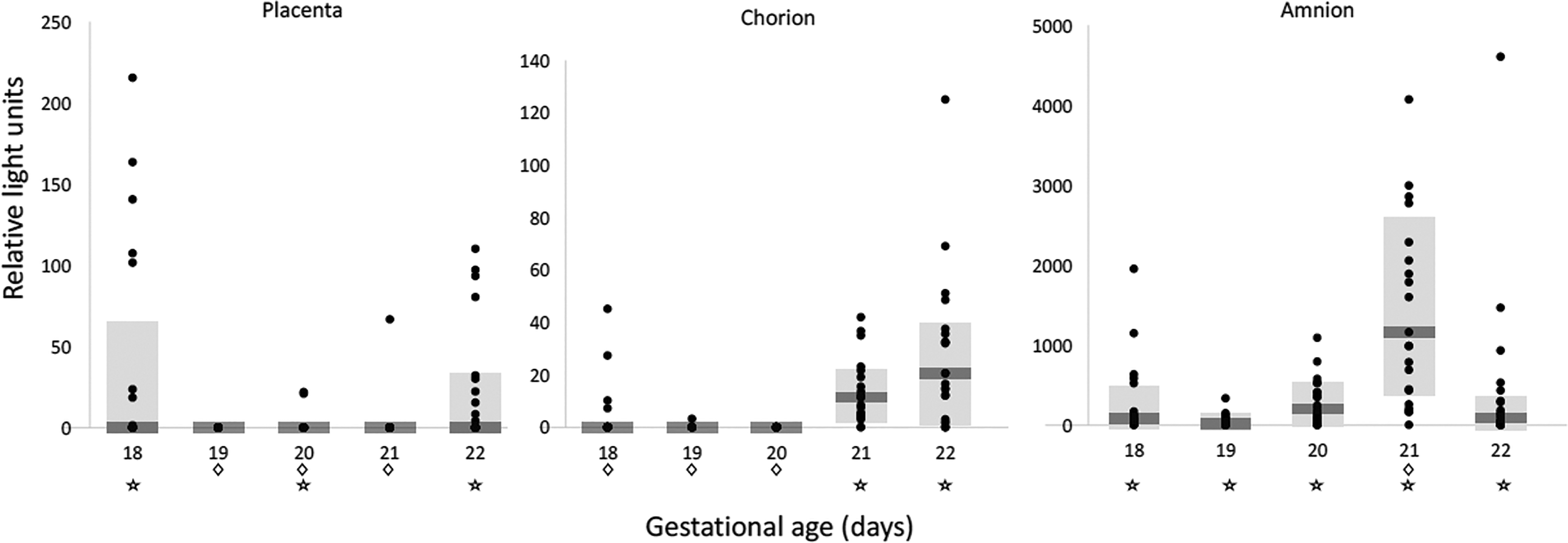
Time-dependent homing of donor afMSCs to fetal annexes demonstrated a consistent nonlinear gestational age-dependent relationship of luciferase activity following a parabolic bimodal pattern, characterized by higher early preterm (E18) and late-term (E22) RLU levels. Shaded area = median ± interquartile range; ⋆, difference compared with acellular luciferase control fetuses at the same gestational age (P < 0.05, Mann-Whitney U-test); ◊, difference compared with term fetuses (P < 0.05, Wald test). afMSC, amniotic fluid-derived mesenchymal stem cell; E, gestational day; RLU, relative light unit.
In the transwell invasion and migration assay, the presence of cells was documented by luminometry in 21/53 (39.6%) of the receiving wells, in suspension and/or attached to the plastic substrate, and in all of the screened gestational membrane sets. Neither collagen coating nor the presence of FBS had an impact on enhancing cell invasion and migration (P = 0.39).
Discussion
In light of the juxtaposition of the placenta with the gestational membranes and amniotic fluid, it is plausible that cell populations would migrate between them. Surprisingly, however, this had yet to be demonstrated. Our results are strongly suggestive of that being the case. The in vivo data after intra-amniotic injection show a close parallel in the bimodal temporal distribution of donor afMSCs in the gestational membranes and the placenta, which would be incompatible with passive cell clearance, while being highly indicative of transmembrane migration as the mechanism of MSC interchange between these sites. Although the data from the chorion were somewhat different from the amnion, they were both congruent with such a process. Importantly, the second peak in luminescence observed in both gestational membranes near term, combined with the virtual absence in the amniotic fluid at that time, would indicate a “two-way street,” that is, the direction of migration could conceivably be both from the amnion to the chorion to the placenta, as well as vice-versa. Such a bidirectional pattern of movement would also suggest that MSC traffic could be a component of normal (and thus possibly also abnormal/preterm) labor. At the same time, the in vitro data showed that these cells are indeed capable of migrating through gestational membranes, independent of chemotactic and nutritional gradient stimuli, further corroborating the in vivo observations.
Before elucidating exactly how donor MSCs can cross through gestational membranes, we needed to first determine whether there was any evidence at all of such a path. A more detailed mechanistic analysis of how that process happens, and what drives it, would be beyond the scope of this study. It is known, however, that MSCs express cell adhesion molecules that can facilitate transmigration, for example, across endothelial barriers [21,22]. In fact, these molecules seem to be central to the ability of MSCs to migrate between the fetal and maternal circulations [22,23]. Nonetheless, MSCs have not yet been shown to migrate through whole tissue layers such as gestational membranes, and previous studies have analyzed MSC homing only after intravenous, rather than intra-amniotic injection. It is reasonable to speculate that, if MSCs can migrate through endothelial layers, comparable mechanisms could also be at play in their apparent ability to migrate through the gestational membranes and placenta. Our results warrant further scrutiny into that. Incidentally, we procured stomach fluid as a means to examine whether fetal swallowing could also contribute to donor cell routing, but our data would suggest that this did not occur. Still, the absence of the cells at that location does not rule out the possibility that these cells may possibly reach the lumen of the gastrointestinal tract.
Of note, significantly greater activity was observed in amnion samples when compared with chorion and placenta. This difference cannot be explained by contamination of the amnion with free-floating donor cells, in that, besides the methodology to avoid such contamination as described above, there were no statistically significant differences in amniotic fluid luminescence from cell-injected animals at each time point, when compared with the acellular controls. On the other hand, one possible contributing factor is the fact that the amnion is avascular, and the hemoglobin content of amnion lysates was likely much lower compared with chorion and placenta, if at all present. When present in lysates, hemoglobin can absorb some proportion of emitted light (a recognized limitation of the luciferase assay) and this could have contributed to the observed differences in measured RLUs between amnion samples and samples from the vascularized sites, that is, chorion and placenta [24].
Such hemoglobin interference, along with small sample volumes, methodological dilution factors, and the described low concentrations of circulating MSCs in the fetus may also have limited our ability to detect donor MSCs in the peripheral blood samples [24,25]. Possibly, other methods such as separation of cells into a low-density fraction before isolation followed by culture of MSCs may improve the sensitivity to detect them from that particular source in future experiments. Still, the robust presences of donor cells in the placenta, as well as in the umbilical cord, fetal bone marrow, spleen, and other sites, as demonstrated in our previous studies, do indicate hematogenous trafficking [16,17]. This is further corroborated by the previous identification of these cells at select sites in the mother [15].
Another possible explanation for the higher luciferase activity in amnion samples is the well-described affinity of MSCs to home to areas of inflammation and tissue damage [26,27]. After intra-amniotic injections, donor MSCs could possibly have been directed to the injection site where the gestational membranes were disrupted, to contribute to their repair. Furthermore, if the injection solution was in any way immunogenic, it could have caused some degree of subclinical chorioamnionitis, which in turn could attract more donor cells to the membranes. In addition, specific chemoattractants such as the complement activation products C3a and C5a, interferon gamma, and others have been shown to impact fetal MSC migration in vitro and could be at play in our model as well [28].
Regardless of these limitations and conjectures, this study lends further support to the notion that the amniotic cavity can be an easy access point for at least select cells to reach the placenta, which is a known gateway to the fetal circulation. Indeed, as alluded to above, donors MSCs have been previously documented in the fetal circulation after TRASCET [16,17]. This perspective significantly expands foreseeable therapeutic applications of TRASCET. It also provides provocative insight into a previously unrecognized pattern of prenatal MSC traffic.
Footnotes
Author Disclosure Statement
No competing financial interests exist from any of the authors.
Funding Information
This work was supported by the Kevin and Kate McCarey Fund for Surgical Research at Boston Children's Hospital.