
Abstract
Nerve regeneration remains a challenge. Patient-derived induced pluripotent stem cell (iPSC)-differentiated neural stem cells (NSCs) provide a promising hope. Zinc is closely involved in central nervous system development and metabolism, but its role on iPSC neural differentiation is elusive and zinc detection methods in live cells are limited. In this study, intracellular zinc was detected in real time by a zinc fluorescent chemosensor and was shown to be increased during the iPSC neural induction process. iPSC neural differentiation was promoted with the addition of zinc chloride (ZnCl2) and inhibited with the addition of zinc chelator N,N,N0,N0-tetrakis(2-pyridylmethyl)-ethylenediamine, indicated by western blot and enzyme-linked immunosorbent assay analysis of NSC marker Nestin expression and measurement of neurite-like structures. Mechanistically, the phosphorylation level of ERK1/2 and STAT3 was changed with the zinc level, suggesting that zinc may affect the neural differentiation of iPSCs through ERK-STAT signaling. In conclusion, our study shows the important role of zinc in iPSC neural differentiation and suggests a new idea for iPSC-derived NSC application in nerve regeneration.
Introduction
The regeneration of nerve tissues and treatment of neurodegeneration diseases is a huge challenge worldwide. Neural stem cells (NSCs) have the ability to differentiate into neurons, oligodendrocytes, and astrocytes and provide promising hope for neural restoration. However, the shortage of NSCs resource both in quantity and in quality is a big issue for NSC therapy [1]. The birth of induced pluripotent stem cells (iPSCs) in 2006 opened a new horizon of stem cell therapies and translational medicine [2]. Since iPSCs can be reprogrammed from autologous somatic cells, their application in cell therapy will not be limited by immunogenicity and ethical issues. Research indicates that human iPSC-derived neural stem cells (iNSCs) could be an ideal cell source for stem cell therapies because of their relatively easy access, low immunogenicity, and multipotential to differentiate into all neural cell types [3 –5]. However, due to the potential tumorigenicity of iPSCs [6], it is a better and safer method to first differentiate iPSCs into various cell types in vitro [7]. Therefore, the mechanisms of iPSC differentiation have been investigated in various studies.
Cellular zinc homeostasis plays an important role in iPSC differentiation, and zinc deficiency significantly impairs iPSC neuronal differentiation [8]. Furthermore, Moon et al. reported that zinc could increase proliferation and affect differentiation of adipose-derived mesenchymal stem cells (AD-MSC) toward a neuronal fate via ERK1/2 pathway [9]. Most researches focused on the cell proliferation or survival effect of zinc, or on the process of differentiating to mature neurons [10]. Zinc is the second most abundant trace element in human bodies, which has long been known to be an essential element for human health and physiological functions [11]. It contributes to structural component of more than 300 enzymes and participates in their functions [11]. Numerous reports have also revealed a family of DNA binding transcription factors known as zinc-finger proteins [12], which play a role in various biological processes such as DNA synthesis, cell proliferation, and differentiation [13]. It is worth noting that the brain has the highest zinc concentration in the nervous system, representing a 10-fold increase over serum zinc [14]. Disturbance of zinc metabolism is also relevant to many neurodegenerative diseases such as Alzheimer's disease, amyotrophic lateral sclerosis, and Parkinson's disease [15]. It is generally believed that zinc affects neurogenesis by regulating stem cell proliferation and differentiation. Zinc deficiency, during development or adulthood, could impair neurogenesis in the central nervous system by limiting neuronal precursor cells proliferation [16].
In addition to involving the structure of proteins or binding to DNA, about 10%–15% of zinc are in the free form, in other words, a chelatable form, distributed in the cytoplasm [17]. This free form of zinc is considered to participate in many physiological processes, but the specific mechanism remains unknown because little is known about the distribution or behavior of intracellular Zn2+. However, it is difficult to analyze the intracellular Zn2+ with conventional methods. For example, the conventional method inductively coupled plasma can only be used to detect the total concentration of Zn2+ inside and outside the cell, but it is currently not possible to detect the biological behavior of intracellular zinc. Fluorescence detection technology has the advantages of high sensitivity, real-time response, and limited damage to live cells so that it is a suitable method for real-time detection of Zn2+. A number of zinc fluorescent chemosensors (ZFCs) have been reported [18,19].
In this study, we induced iPSCs derived from human gingival fibroblasts (hGF) into NSCs using PSC neural induction medium (NIM) with different zinc concentrations. Zinc chloride (ZnCl2) or N,N,N0,N0-tetrakis(2-pyridylmethyl)-ethylenediamine (TPEN) was added to the NIM. In addition, we utilized a ZFC (designed and synthesized by State Key Laboratory of Coordination Chemistry, School of Chemistry and Chemical Engineering, Nanjing University) to detect changes of intracellular zinc level in real time. It is a ratiometric fluorescent chemosensor that can combine with free Zn2+ in cells in a ratio of 1:1. The combination will change the excitation wavelength of the ZFC so that we are able to compare the zinc concentration relative quantitatively by analyzing the fluorescence images obtained at two different excitation wavelengths. The ratiometric fluorescent chemosensor can eliminate the influence of variations in cell size, membrane permeability, background fluorescence, and concentration of chemosensor by self-calibration of two excitation bands, providing more accurate measurement of intracellular zinc [20]. Furthermore, the relationship between zinc levels and phosphorylation of ERK1/2 and STAT3 was also examined to explore the mechanism of the effect of free zinc on iPSC neural differentiation.
Materials and Methods
Culture of iPSCs
iPSCs were reprogrammed from hGF in our previous works [21,22]. iPSCs were cultured using Nuwacell™ Nova hPSC Medium (Nuwacell, Hefei, China) in six-well plates pretreated with Matrigel (Corning, NY) overnight. The culture medium was changed daily, and the cells were passaged at 70%–90% confluence. During the first 24 h after passaging, 2.5 μM Blebbistatin (Nuwacell) was added into the medium. We used iPSCs at passage numbers 30–50 for all experiments in this study.
Neural induction of iPSCs and production of iNSCs
iPSCs were induced to iNSCs using PSC Neural Induction Medium (NIM) (Thermo Fisher Scientific, MA). In brief, about 2.5–3.0 × 105 iPSCs were seeded in each well of a six-well plate. When the cells reached 15%–25% confluency, the culture medium was changed to NIM. NIM was changed every 2 days. After day 4 of neural induction, the volume of NIM was doubled. At day 7 of neural induction, the P0 NSCs were harvested and expanded.
Establishment and detection of different intracellular Zn2+ levels in iPSCs
ZnCl2 and TPEN (Sigma–Aldrich, MO) were added into the culture medium. ZFC was used to detect whether the addition or chelation of Zn2+ in culture medium could affect the intracellular level of Zn2+. About 1.5 × 105 iPSCs were seeded into 35 mm dishes, and after adherent, cells were washed twice using phosphate-buffered saline (PBS) and then treated with 5 μM ZnCl2 or 1 μM TPEN for 20 min at room temperature. After that, cells were incubated with ZFC diluted in PBS (2.5 μM) in dark at room temperature for 20 min. Images were taken by Zeiss LSM-710 confocal microscope. The excitation wavelength of free ZFC is 633 nm, which will alter to 543 nm after binding to Zn2+. Two fluorescence images were taken at excitation wavelengths of 543 nm (T1) and 643 nm (T2) separately and the band path was set at 641–741 nm. Ratiometric images were synthesized by Zen software. Six cells were circled randomly in the field of view for analysis. The ratio of fluorescence intensity in T1 and T2 (T1/T2) represents the relative zinc level.
Detection of intracellular Zn2+ during iPSC neural induction process
About 0.5–1.0 × 105 iPSCs or iNSCs were seeded into 35 mm dishes. After the cells were adherent, the medium in iPSC dishes was changed to NIM. The intracellular zinc of iPSCs at day 0, 2, 3, and 5 of neural induction and iNSCs was detected by ZFC. Cells were washed with PBS and incubated with 2.5 μM ZFC in dark at room temperature for 20 min. ZFC solution was changed to PBS subsequently, and images were taken by Zeiss LSM 710 confocal microscope. The analysis method is the same as above.
Immunofluorescence of NSC markers
The expression of NSC marker Nestin, Sox2, and Pax6 was detected by immunofluorescence. Cells were washed using PBS three times and fixed in 4% paraformaldehyde for 15 min at room temperature. Then, the cells were washed twice and incubated for 10 min with PBS containing 0.2% Triton X-100 (Sangon, Shanghai, China). Cells were washed by PBS twice and then incubated in PBS containing 5% bovine serum albumin (BSA) for 1 h at room temperature. After blocking for 1 h, the cells were incubated overnight at 4°C with following diluted primary antibodies: anti-human-Nestin (1:100), anti-human-Pax6 (1:100), and anti-human-Sox2 (1:100) (Proteintech Group, Rosemont, IL). Cells were washed with PBS three times for 5 min and then incubated with corresponding diluted secondary antibodies [fluorescein isothiocyanate (FITC) anti-rabbit immunoglobulin G (IgG), FITC anti-mouse IgG; Proteintech Group] for 1 h in dark at room temperature. After incubated with antibodies, cells were washed using PBS three times for 5 min and then incubated with 4′,6-diamidino-2-phenylindole (1 μg/mL) for 15 min at room temperature. Images were taken on a Nikon A1 confocal microscope.
Flow cytometry analysis
Flow cytometry analysis was performed to detect the expression of CD133, CD45, and CD34 of iNSCs. Cells were prepared to single cell suspension when reaching 80%–90% confluency. Three cell samples containing about 1 × 106 cells in 100 μL PBS were incubated with 5 μL of following antibodies: anti-human CD133 phycoerythrin (PE), anti-human CD45 allophycocyanin (APC), and anti-human CD34 FITC at 4°C for 1 h. Another sample of cells was incubated with corresponding isotypes (mouse IgG1 PE, mouse IgG1 APC, mouse IgG1 FITC; BioLegend, CA) in the same condition. The stained cells were washed twice and measured by flow cytometry using the BD FACSCalibur (Becton Dickinson, NJ) and the Flowjo V10 software.
Cell proliferation and cytotoxicity assay
WST-8 Cell Counting Kit-8 (CCK-8; APExBIO, Houston) was used to analyze the cell proliferation. Cells were seeded 2,000 per well in five 96-well plates. One day after adherent, medium was changed to NIM. There were eight groups on each plate: the control group, cells treated with 0.5, 1, or 2 μM TPEN, 2.5, 5, 10, or 20 μM ZnCl2. Each group had four duplicate wells. NIM was changed every 2 days, and one of the plates was change to NIM containing 10% CCK8 every 24 h. After CCK8 was added, the plate was incubated at 37°C for 3 h and analyzed at 450 nm using a SpectraMax M3 (Molecular Devices, CA).
Apoptosis assay
The cell apoptosis was tested by FACS analysis using ANNEXIN V-FITC/propidium iodide (PI) apoptosis kit (Solarbio, Beijing, China). About 2.5–3.0 × 105 iPSCs were seeded in six-well plates and treated with different concentrations of ZnCl2 or TPEN, the same as those in the proliferation assay (cells treated with 0.5, 1, or 2 μM TPEN, 2.5, 5, 10, or 20 μM ZnCl2, and the control group without ZnCl2 or TPEN). The medium was changed every 2 days, and at day 5, the cells reached 80%–90% confluency. Cells were washed with PBS twice and harvested using 0.25% trypsin (Beyotime, Shanghai, China) without ethylenediaminetetraacetic acid. In each sample, about 1 × 105 cells were harvested and resuspended using 100 μL binding buffer. Five microliters of Annexin V-FITC was added into each sample and cells were gently mixed and incubated at room temperature for 10 min in dark. Then, 5 μL PI was added into each sample and cells were incubated at room temperature for 5 min in dark. After that, PBS was added to the samples to 500 μL and measured by flow cytometry using the BD FACSCalibur (Becton Dickinson) and the Flowjo V10 software.
Length of neurite-like structure measurement
To evaluate the efficiency of neural differentiation in different groups, the length of neurite-like structures of iNSCs was measured. There were six groups: the control group, cells added 2.5, 5, 10 μM ZnCl2, or 1 μM TPEN, and cells added 1 μM TPEN and 5 μM ZnCl2. After 1–2 days of adherent, images were taken under an optical microscope. Using a 20 × objective, three different fields were chosen randomly. In each field, at least five different cells were measured. Neurite-like structure length was evaluated by the ImageJ software.
Enzyme-linked immunosorbent assay of Nestin expression
At day 7 of neural induction, cells were washed twice with cold PBS and lysed in RIPA containing 1% phenylmethylsulfonyl fluoride and 1% phosphatase inhibitor (Beyotime). The protein concentrations were measured by BCA Protein Assay Kit (Beyotime). Then, the concentration of Nestin in samples was detected by human Nestin enzyme-linked immunosorbent assay (ELISA) kit (Cusabio, Wuhan, China) according to the manufacturer's instructions. Plates were read at 450 nm using a SpectraMax M3 (Molecular Devices). Three replicates of each sample were prepared in this experiment.
Western blotting assay
Proteins were isolated at the day 3, 5, and 7 of neural induction. Protein isolation and concentration measurement are the same as above. Then, the samples were resolved by electrophoresis on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel. The separated proteins were transferred to 0.45 μm polyvinylidene difluoride (PVDF) membranes at 300 mA for 80 min. The membranes were blocked in 5% BSA-PBS solution at room temperature for 1 h, then washed using tris buffered saline containing 0.1% Tween (TBST) three times for 10 min, and incubated in diluted primary antibodies at 4°C overnight (Nestin, GAPDH, p-ERK1/2, ERK1/2, p-STAT3, STAT3). After incubating the primary antibodies, membranes were washed three times for 10 min using TBST and incubated in diluted secondary antibodies [anti-mouse IgG-horseradish peroxidase (HRP), anti-rabbit IgG-HRP; Proteintech Group]. The proteins on the PVDF membranes were detected with enhanced chemiluminescence detection system.
Inhibition of ERK1/2 pathway
To test whether the zinc levels affect iPSC neural differentiation through the ERK1/2 pathway, the ERK inhibitor VX-11e (MedChemExpress, NJ) was added into NIM during neural induction. The final concentration of VX-11e, ZnCl2, and TPEN was 10, 5, and 1 μM, respectively. There were eight groups in this experiment. In each group, cells were cultured with or without VX-11e, ZnCl2, or TPEN. At the day 7 of neural induction, proteins were isolated and expression of Nestin, phosphorylation of ERK1/2 and STAT3 were detected by western blot.
Statistical analysis
All measurements are expressed as mean ± standard deviation from at least three independent experiments. Data for these measurements were analyzed using Student's t-tests for paired comparisons. GraphPad Prism 7.0 (GraphPad Software, San Diego, CA) software was utilized to analyze and demonstrate the statistical significance of the assays. Difference at P < 0.05 was considered statistically significant. The significance between groups was marked on the graphs.
Results
iPSCs derived from hGF were induced efficiently into iNSCs
At passage number 30–50, iPSCs exhibited embryonic stem cell (ESC)-like morphology and were suitable for neural induction. During the process of iPSC neural induction, there were obvious changes in cell morphology (Fig. 1A–E). At day 3–5, when the cells almost reached 80%–90% confluency, neurite-like structures could be seen around the cell colonies (Fig. 1C, D). After passaging, the neurites became longer and cell morphology was similar to NSCs (Fig. 1E). We conducted immunofluorescence and flow cytometry analysis to identify iNSCs. Immunofluorescence showed positive expression of NSC markers Nestin, Sox2, and Pax6 after induction (Fig. 1G). Flow cytometry analysis showed that the proportion of CD133-positive expression of iNSCs was 78.4% compared with the isotype of 1.89%, and CD45 and CD34 were expressed negatively (Fig. 1F).
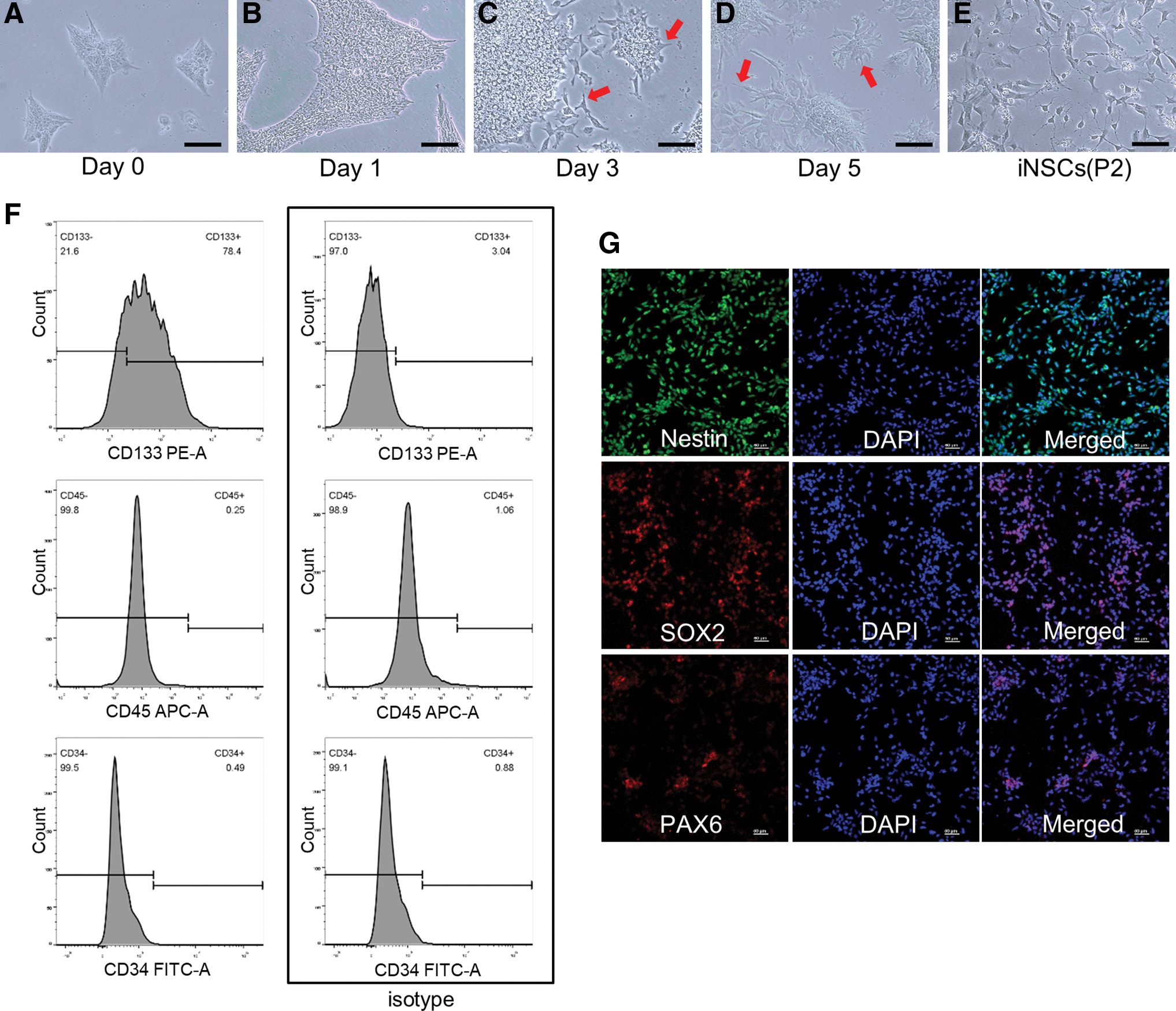
Neural induction of iPSCs using NIM and identification of iNSCs.
Zinc addition or chelation in medium affected iPSC intracellular zinc level
To establish different intracellular zinc levels, we added ZnCl2 or zinc chelator TPEN into the cell culture medium and detected intracellular zinc level using the ZFC. Images taken by confocal microscope are shown in Fig. 2A. Statistical analysis showed that the intracellular zinc level significantly changed (Fig. 2B). It means that direct addition or chelation of zinc in medium could affect intracellular zinc level. The ZFC showed rapid response and high sensitivity.
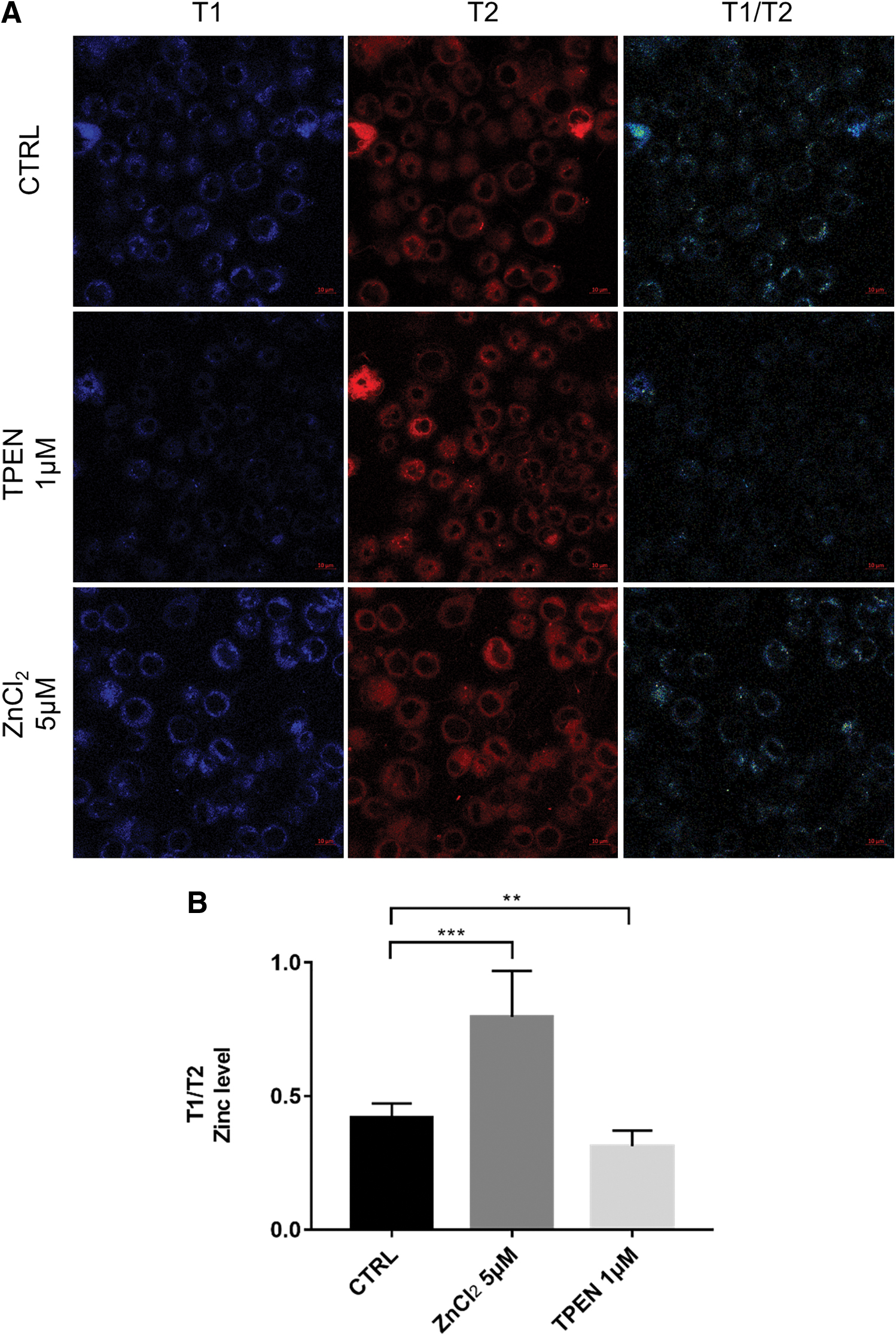
Addition or chelation of zinc in medium affected intracellular zinc level.
Intracellular zinc level increased during iPSC neural induction
During the process of iPSC neural induction, we used ZFC to detect the change of intracellular zinc level. Fluorescence images are shown in Fig. 3A. Compared with iPSCs, intracellular zinc level of cells at day 5 of neural induction showed significant difference. The difference between iNSCs and iPSCs was more significant. It is worth noting that at day 2 and 3 of neural induction, although there is no statistical difference compared with iPSCs, there was an upward trend of intracellular zinc level (Fig. 3B).

Intracellular zinc level increased during the process of iPSC neural induction.
Low doses of ZnCl2 and TPEN showed little impact on cell apoptosis and proliferation
Since our aim was to explore the role of Zn2+ in iPSC differentiation, we first examined the effect of zinc concentration on cell apoptosis and proliferation. We found that little difference was observed among groups with additional ZnCl2 concentration ranging from 0 to 20 μM and TPEN concentration ranging from 0.5 to 1 μM at day 5 of neural induction. However, when the concentration of TPEN increased to 2 μM, the cell apoptosis rate increased (Fig. 4A). In the cell proliferation assay, when the concentration of ZnCl2 increased to 20 μM and the concentration of TPEN increased to 2 μM, cell proliferation appeared to be inhibited (Fig. 4B). Therefore, to chelate Zn2+ as much as possible without affecting cell proliferation, 1 μM TPEN was used in our subsequent experiments.
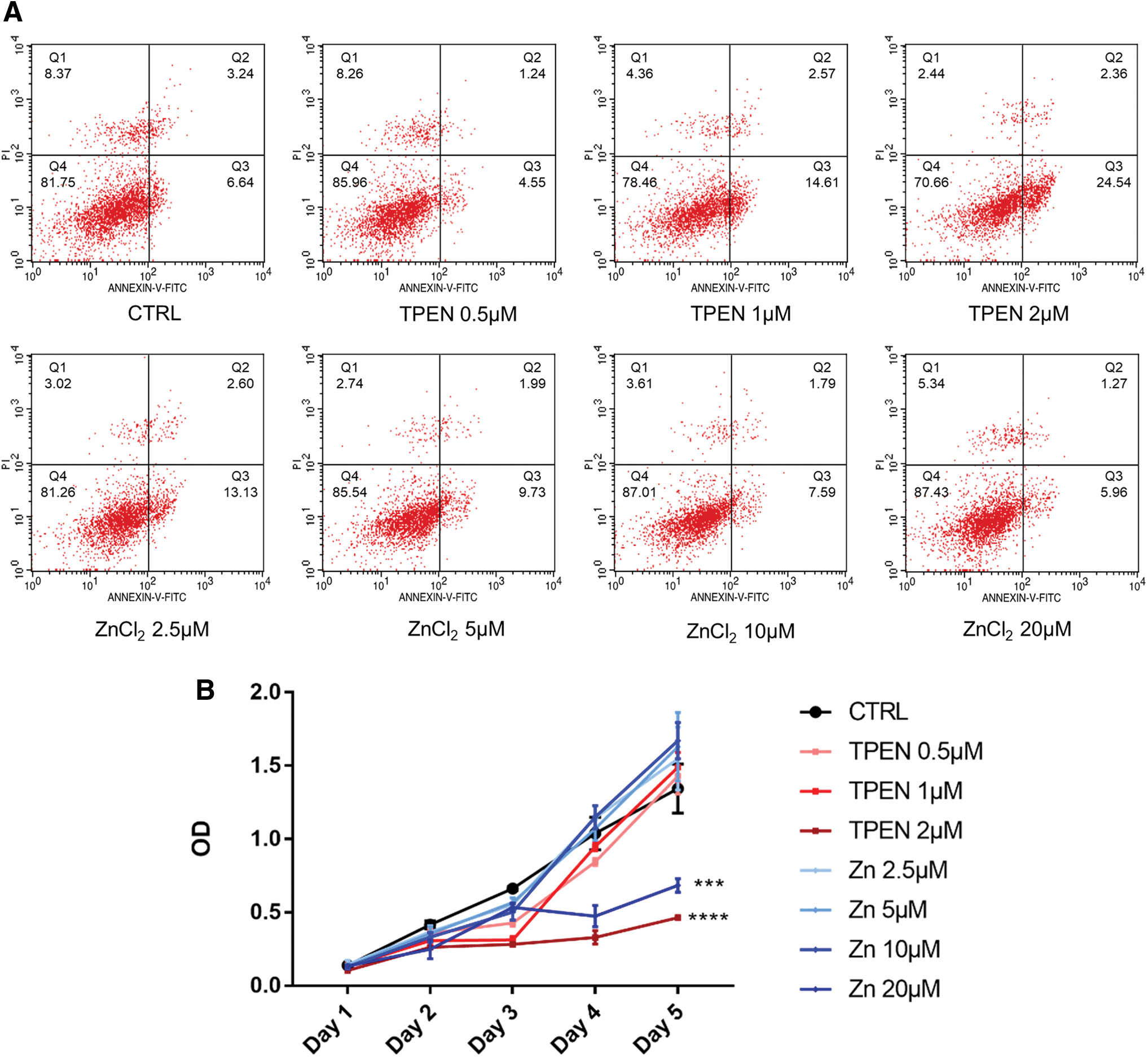
Impact of different concentrations of ZnCl2 and TPEN on cell proliferation and apoptosis.
Zinc addition or chelation affected efficiency of iPSC neural differentiation
The intracellular zinc level could be affected by changing the concentration of zinc in the culture medium, so in the following experiments, we explore the effect of different intracellular zinc levels on iPSC differentiation by zinc addition or chelation in NIM (Fig. 8A). Cell morphology of different groups after passaging is shown in Fig. 5A–F. When treated with TPEN during neural induction, iNSCs appeared more iPSC-like after passaging (Fig. 5E). We measured neurite-like structure length of single cells, and the statistical analysis is shown in Fig. 5G. To evaluate the efficiency of neural induction, NSC marker Nestin was also detected by ELISA and western blot. At day 7, the results of ELISA showed that expression level of Nestin was higher in groups that added ZnCl2 compared with the control group. In the zinc chelation group, the expression level of Nestin was significantly lower than the control group (Fig. 5H). Consistent with the results of ELISA, western blot assay showed that Nestin expression level was significantly higher in zinc addition groups compared with the control group, whereas Nestin expression level in the TPEN group was significantly lower. No significant difference of Nestin expression was observed between the group with both ZnCl2 and TPEN and the control group (Fig. 5I, J).

Different zinc levels affected the efficiency of iPSC neural differentiation.
Zinc addition accelerated iPSC neural differentiation
To further investigate the role of Zn2+, the neural induction was suspended at day 3, 5, and 7 separately and cells were passaged. Western blot analysis of Nestin expression shown in Fig. 6A, B revealed that in the zinc addition group at day 3, the Nestin expression level showed no significant difference compared with the control group at day 7. In the zinc addition group at day 5, the Nestin expression level was significantly higher than the control group at day 7. At day 3, cells exhibited iPSC-like colonies while the zinc addition group showed more neurite-like structures (Fig. 6C–F). Neurite-like structures were more obvious at day 5 (Fig. 6G–J). Immunofluorescence showed that Nestin expression in the zinc addition group at day 5 showed no significant difference compared with the control group at day 7, indicated that Nestin was expressed 2 days earlier. The results also showed decreased expression of Nestin in the TPEN group, which is consistent with western blot analysis (Fig. 6R).

Zinc addition accelerated iPSC neural differentiation.
Phosphorylation of ERK1/2 and STAT3 was associated with zinc levels
We also preliminarily explored the possible mechanism of how intracellular zinc affected iPSC neural differentiation. p-ERK1/2, ERK1/2, p-STAT3, and STAT3 levels at day 7 of iPSC neural induction were detected by western blot (Fig. 7A). Phosphorylation of ERK1/2 and STAT3 was promoted with the addition of ZnCl2 while inhibited with the addition of TPEN. Phosphorylation levels showed no significant differences among groups of different ZnCl2 levels (Fig. 7B, C). Since there might be a close relationship between zinc and ERK1/2 phosphorylation, we blocked the ERK pathway to observe the effects on Nestin expression and STAT3 phosphorylation. After treated with 10 μM ERK inhibitor VX-11e during the induction process, cell morphology of iNSCs was more iPSC-like (Fig. 7D–G). Western blot showed that Nestin and p-STAT3 expression was also downregulated when the phosphorylation of ERK1/2 was inhibited (Fig. 7H). iPSC neural differentiation and STAT3 phosphorylation were significantly inhibited after treated with ERK inhibitor (Fig. 8B). In groups with VX-11e, either zinc was chelated or added, no significant difference was observed.

Zinc level regulates phosphorylation of ERK1/2 and STAT3 and inhibition of the ERK-STAT pathway impaired neural differentiation of iPSCs.

Overview of the study.
Discussion
There have been several methods to differentiate PSCs into NSCs including embryoid body or neurosphere formation [23,24]. Chambers et al. found that dual inhibition of SMAD signal can induce neural differentiation of PSC efficiently in adherent culture conditions [25]. In this study, we used PSC NIM to induce iPSCs into NSCs, which also does not require the intermediary step of embryoid body formation. In a previous study, iNSCs induced by NIM showed multipotential to differentiate into neuron, astrocyte, and oligodendrocyte [7]. During the induction process, the intracellular zinc level gradually increased by real-time detection of ZFC. Difference between iPSCs and iNSCs after passaging was more significant. One possible explanation might be that zinc is essential for neural differentiation of iPSCs and iNSC functions. It can be speculated that zinc plays an important role in the differentiation of iPSCs into NSCs. The change of intracellular zinc level might also be related to the alternate expression of zinc homeostasis genes during the neural differentiation process [8].
When added ZnCl2 reached to 20 μM, cell proliferation occurred to be inhibited while zinc level showed little impact on apoptosis. It means that these concentrations of ZnCl2 do not lead to cell apoptosis, only affect cell proliferation. Different cell lines seem to have different tolerance to zinc levels. In AD-MSC, 30–100 μM ZnCl2 promoted proliferation while cytotoxicity was observed until the concentration reached 300 μM [9]. However, in our study, low doses of zinc did not promote cell proliferation. It is also possible that there has been sufficient Zn2+ in the NIM so that only additional 20 μM ZnCl2 can lead to inhibition of proliferation. This result indicates that high concentration of Zn2+ showed toxicity to the cells, which is consistent with the fact that high concentration of Zn2+ has been proved to be neurotoxic [26,27].
Zinc has always been considered vital to cell functions including cell proliferation and differentiation. Wang et al. reported that maternal mice with zinc deficiency diets impair brain Nestin expression of offspring mice, which suggests that zinc deficiency suppresses the development of NSCs [28]. Rather than the intracellular free zinc, role of zinc finger proteins in iPSC differentiation or maintenance of pluripotency has raised more attention [29 –31]. Few researches focused on the role of free Zn2+ in iPSC neural differentiation. The results of this study indicate that the chelatable Zn2+ distributed in the cytoplasm are also indispensable for neural differentiation. The change of zinc level in medium causes the corresponding change of intracellular zinc level, which possibly regulates the neural differentiation of iPSCs. Notably, we preliminarily proved that the addition of ZnCl2 could accelerate the neural induction process, and chelation of Zn2+ significantly impaired neural differentiation.
Zinc is proved to be an important modulator of ERK1/2 signaling [32]. It is confirmed by our results that phosphorylation of ERK1/2 was promoted by zinc addition while inhibited by zinc chelation during the process of iPSC neural differentiation. In addition to ERK1/2, STAT3 was also shown to be regulated by zinc. It is also evident that ERK1/2 and STAT3 contribute to the neural differentiation of pluripotent stem cells. When STAT3-dominant negative ESCs were induced to neural differentiation, the Nestin expression decreased and fewer neural progenitor cells were produced [33]. Inhibition of p-ERK1/2 also significantly reduced the number of NeuN-positive cells after neural induction using retinoic acid [34]. After blocking the ERK pathway using VX-11e, the downstream STAT3 phosphorylation is also inhibited. Among groups treated with VX-11e, no significant difference of Nestin expression was observed. That is to say, block of ERK-STAT pathway inhibited neural differentiation regardless of the zinc level changes. Therefore, we hypothesized that zinc regulates neural differentiation by regulating ERK1/2 phosphorylation, STAT3 phosphorylation, and Nestin expression changes subsequently. During the process of iPSC neural differentiation, zinc might regulate ERK1/2 phosphorylation by directly inhibiting phosphatases that are involved in ERK1/2 dephosphorylation or indirectly activating ERK phosphokinase through activating upstream from the ERK1/2 cascade such as GRP39 and MMP [32].
In conclusion, we demonstrate that low doses of ZnCl2 or TPEN addition present little impact on cell proliferation but significantly promote iPSC differentiation through ERK-STAT signaling. During the iPSC neural differentiation process, intracellular zinc level increased detected by fluorescence detection technology, an effective method for detection of intracellular metal ions such as Zn2+. However, it should be noted that this study only focused on changes in cell morphology and Nestin expression and examined a few molecules that were regulated by zinc. To the best of our knowledge, there are many pathways that can lead to neural differentiation of iPSCs or maintenance of stemness of NSCs in vitro, such as Wnt/β-catenin, TGF/BMP, Notch, and JNK/MAPK [35]. Further research on the detailed mechanisms of iNSC function in vivo is needed.
Footnotes
Acknowledgment
The authors thank State Key Laboratory of Coordination Chemistry, School of Chemistry and Chemical Engineering, Nanjing University, for the provision of zinc fluorescent chemosensor.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (grant number 81200770) and the Nanjing Medical Science and technique Development Foundation (grant number YKK18122).