
Abstract
Esophagus and trachea arise from a common origin, the anterior foregut tube. The compartmentalization process of the foregut into the esophagus and trachea is still poorly understood. Esophageal atresia/tracheoesophageal fistula (EA/TEF) is one of the most common gastrointestinal congenital defects with an incidence rate of 1 in 2,500 births. EA/TEF is linked to the disruption of the compartmentalization process of the foregut tube. In EA/TEF patients, other organ anomalies and disorders have also been reported. Over the last two decades, animal models have shown the involvement of multiple signaling pathways and transcription factors in the development of the esophagus and trachea. Use of induced pluripotent stem cells (iPSCs) to understand organogenesis has been a valuable tool for mimicking gastrointestinal and respiratory organs. This review focuses on the signaling mechanisms involved in esophageal development and the use of iPSCs to model and understand it.
Introduction
The esophagus is a muscular tube located between the pharynx and the cardia of the stomach and the only section of the gastrointestinal tract lacking any digestive or absorptive properties. The primary function of the esophagus is facilitating the passage of food from the mouth to the stomach and preventing acid refluxes. During embryonic development, the esophagus originates from an endodermal diverticulum in the anterior region of the foregut tube. Highly regulated cellular behavior and molecular events are responsible for separating the common foregut tube into the esophagus and the trachea [1], whereas epithelial biogenesis establishes proper epithelial morphogenesis of the esophagus and the trachea. Following foregut separation, the esophagus is lined with columnar cells, which then transform into a stratified squamous epithelium surrounded by two layers of smooth muscle with embedded enteric nerves and glial cells [2]. By contrast, the trachea is lined with pseudostratified epithelium and C-shaped cartilage rings [3]. Disturbances in the relevant signaling mechanisms may lead to severe esophageal congenital anomalies, such as esophageal atresia with or without tracheoesophageal fistula (EA/TEF) [4]. The same signaling pathways have been associated with the pathogenesis of esophageal diseases such as Barrett's esophagus that affect the epithelial cellular morphology [5]. Combined studies in mouse models and human-induced pluripotent stem cells (iPSCs) reveal the striking similarities of esophagus development between mouse and human [6,7].
Foregut Separation and Esophagus Development
At 4–5 weeks of human embryonic development, and mouse embryonic day 11.5 (E11.5), the anterior foregut tube is patterned down the sagittal axis, to separate dorsally into the esophagus (digestive system), and the trachea ventrally (respiratory system) (Fig. 1) [1]. Both presumptive organs are surrounded by splanchnic mesoderm. Bidirectional communication between the foregut endoderm and mesoderm orchestrate esophageal and tracheal development [8]. The esophagus develops from the dorsal side of the anterior foregut tube. The nascent esophageal epithelium is made up of a single layer of cuboidal ciliated cells, which is gradually replaced with stratified squamous epithelial cells with a basal proliferative layer. It is noteworthy that structural differences between human and mouse esophageal epithelium exist. The human esophageal epithelium is nonkeratinized consisting of cell layers folded along papillae [9], whereas murine esophageal epithelium is keratinized and contains fewer cell layers where proliferation is restricted to basal cells [10]. Unlike mice, submucosal glands are found in the human esophageal epithelium, which could serve as a source of progenitor cells responding to cell injury [11]. The trachea lined with pseudostratified columnar epithelium develops at the ventral side of the foregut tube (Fig. 2).

Development of esophagus and congenital anomalies:
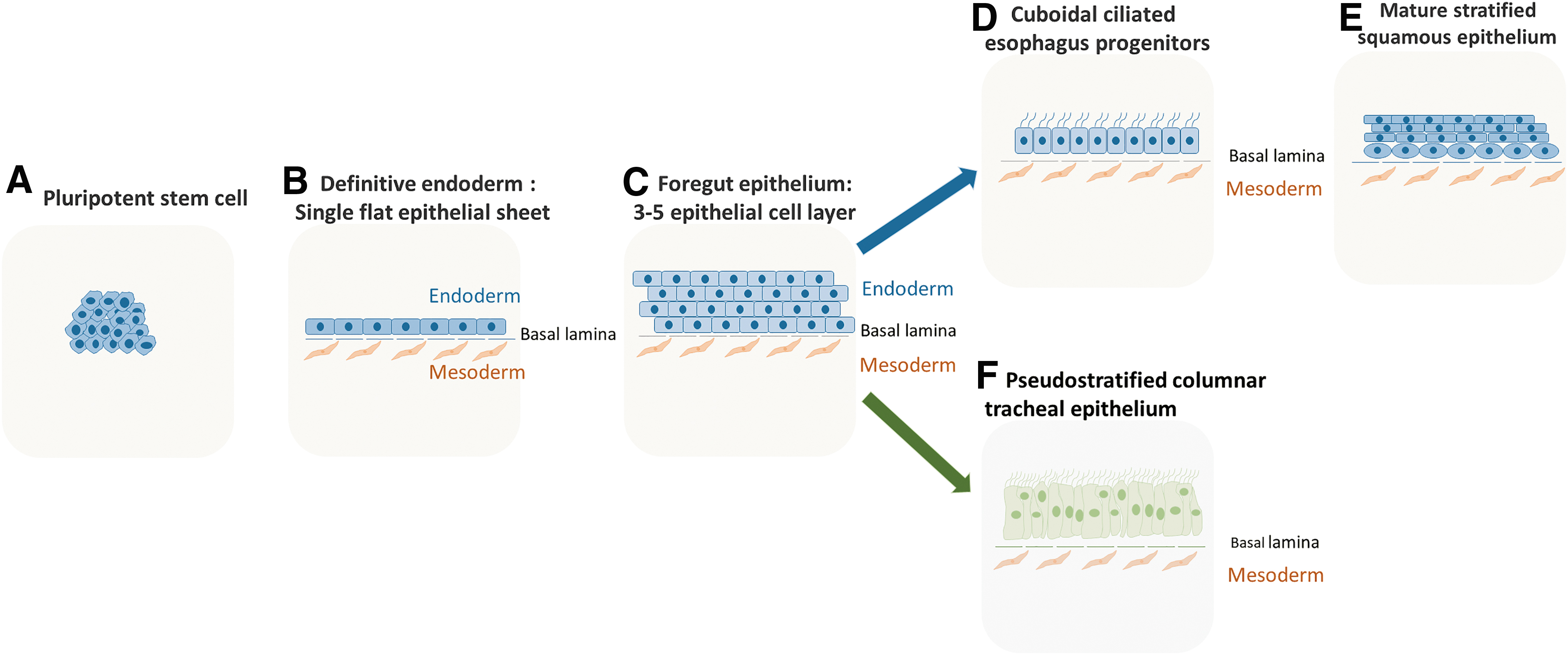
Epithelial morphogenesis of the esophagus and trachea:
Several models of foregut compartmentalization have been proposed to explain the splitting of the foregut tube into the esophagus and trachea [12]. The septation model hypothesizes that separation initially occurs at the origination of lung buds and progresses in a rostral direction along the dorsal-ventral (D/V) midline to form a septum separating the two tubes (Fig. 1B) [13]. Recent studies have shown that this model, which involves the fusion of lateral mesenchymal ridges, mirrors the development of the esophagus and the trachea. Kim et al. observed a unique epithelial population termed midline epithelial cells (MECs) at the D/V boundary, which fuses to form a septum leading to the formation of the trachea and esophagus [14]. In another study using mouse and Xenopus models, Nasr et al. suggest that epithelial remodeling requiring a small GTPase and specific extracellular matrix degradation allows the mesenchyme to invade and resolve the septum using the Hedgehog/Gli signaling pathway [14]. The septation model seems to be the most consistent with experimental evidence.
At week 4 of human gestation (E9–9.5 in the mouse), neural crest cells (NCCs) scatter on the dorsal side of the foregut and encircle the gut. Muscle differentiation begins when NCCs enter the primitive gut [15]. By the end of week 6 (E14 in the mouse), NCCs have migrated within the mesenchyme surrounding the primitive esophagus and start to form the myenteric plexus. In the Mash1−/− mouse knockout model characterized by the absence of enteric neurons in the anterior foregut, the development of the esophagus is not altered, suggesting that intrinsic neurons do not play a significant role in the separation of the foregut [16]. By week 8 (E15 in the mouse), the NCCs are located external to the presumptive esophageal internal circular smooth muscle cells [17,18]. At birth, the lower segment of the esophagus consists of smooth muscle cells, and the upper segment consists of striated muscles that originate from cranial mesoderm [19].
Esophageal Atresia/Tracheoesophageal Fistula
EA/TEF is the most frequent congenital anomaly of the esophagus affecting 1 in 2,500 to 3,000 births [4], where the upper segment is not connected to the lower part of the esophagus and the stomach. In most cases, EA is accompanied by an abnormal connection between the esophagus and the trachea, known as a fistula (Fig. 3). This malformation requires urgent surgery with subsequent associated long-term morbidity, necessitating a lifelong follow-up [20,21]. Several types of EA/TEF have been described based on the location of the malformation and the affected structures, with the most common type being type C (80%), where the upper segment of the esophagus ends in a blind pouch and a fistula connects the lower part to the trachea. Other less common subtypes include type A (8%–10%) where no fistula exists, but the esophagus is disconnected, and type H (4%) where the esophagus connects typically to the stomach, but a TEF is connecting the esophagus to the trachea. The rarest malformations are types B and D. In type B, the upper segment of the esophagus is connected to the trachea, and the lower segment ends in a blind pouch, whereas, in type D, TEF exists in the upper and lower part of the esophagus, affecting only 1% (Fig. 3). The exact cause of failure of the foregut separation is still not fully understood. EA/TEF could be due to multiple factors, including genetics, the environment, and immunology [22]. Other associated risks include fetal alcohol exposure, parental age, and maternal diabetes [23,24].

Types of EA and TEF:
EA/TEF-associated anomalies are reported in 30%–50% of the cases [4,25]. These include cardiac, anal, renal, limb, or vertebral malformations, which can be associated in the VACTER/VACTERL syndrome, suggesting common development mechanisms between the different organs. Anomalies such as right aortic arch, aberrant subclavian artery, or vascular rings with double aortic arch, with or without cardiac malformations, are also frequently found in EA/TEF patients [26]. EA/TEF is frequently associated with congenital esophageal stenosis in the distal third of the esophagus [27]. Tracheobronchial remnants are often found in such stenosis, suggesting a localized abnormal D/V patterning of the foregut. Finally, EA/TEF patients also have an increased prevalence of laryngeal clefts, which may be related to the abnormal development of the trachea-esophageal septum [28].
Genes associated with developmental anomalies of the esophagus
Most cases of EA/TEF have not been associated with gene mutations. However, evidence suggests that EA/TEF may have a genetic background: (1) in twins, the concordance rate is higher in monozygotic twins with isolated EA than in dizygotic twins [29]; (2) although rare, hereditary EA/TEF has been reported [30,31]; (3) EA/TEF can be associated with single-gene disorders and karyotype anomalies (deletions or duplications of chromosomes involving genes implicated in trachea-esophagus development [22,23,32]; and (4) genetically modified mice that mimic EA/TEF have shown candidate genes in human esophageal development. Mouse knockout models have been used to mimic the genetics and morphogenetic regulations of foregut bifurcation and subsequent epithelial morphogenesis corresponding to human esophageal anomalies [1]. Mouse models of abnormal foregut development consist of two main categories depending on which organ of the esophagus or trachea is the most affected. Table 1 shows different foregut compartmentalization defects using rodent models with variant degrees of abnormal development of the esophagus and trachea. The first includes abnormal separation of the esophagus with variable hypoplasia in the esophagus leading to EA with or without TEF such as Sox2GFP/COND, endodermal deletion of SOX2, Noggin−/−, Gli2 and Gli3 mutants, Shh−/−, and Foxf1−/−. The other group encompasses impaired outgrowth of the respiratory tract with various levels of hypoplastic trachea and lungs, including NKX2.1−/−, Wnt2/2b−/−, Ctnnb1flox/flox: Shh-Cre, BMP4condKO, BMPr1a−/−, Gli2 and Gli3 mutants, and Shh−/− (Table 1).
Different Rodent Model Foregut Compartmentalization Defects with Variant Degrees of Abnormal Development of the Esophagus and Trachea
Specification refers to the specific expression of SOX2 on the dorsal side and NKX2.1 on the ventral side of the anterior foregut tube before separation. Septation refers to the physical separation of the anterior foregut into two distinct tubes, the esophagus and trachea. Esophagus and trachea refer to the presence of a visible tube with the respective identity. TEF refers to the presence of an esophageal atresia or a TEF clearly described in the study. Lungs refer to the complete or incomplete (hypoplasia) development of the lungs.
Hypoplasia of the esophagus and TEF dose dependent of sox2 expression.
Early Endodermal Deletion (E6.5) of Sox2 results in Esophageal agenesis and deletion of Sox2 after initiation of esophageal development (E9.5) results in esophageal hypoplasia.
Elevated SOX2 expression in NKX2.1 null embryo shown in Que 2007.
Abnormal notochord delamination, but septation preserved.
Not characterized.
Columnar respiratory epithelium along the ventral surface.
Normal expression of SOX2 in the dorsal foregut and abnormal expression of NKX2.1 in the ventral foregut with ectopic expression of Sox2.
The common tube is lined by columnar epithelium.
Tracheal, agenesis related to decreased cell proliferation.
NKX2.1 expression present at E9.25 and lost afterward with expansion of SOX2 expression.
Single tracheoesophageal tube that connects to the stomach.
Endodermal structures missing between the thymus and stomach levels.
NA, not applicable because of the presence of a single tracheoesophageal tube without any septation; NR, not reported; TEF, tracheoesophageal fistula.
Possible mechanisms leading to EA/TEF
Defects in notochord resolution
After gastrulation, a narrow band of cells delaminates from the endoderm to form the notochord, a major regulator of embryonic patterning [33]. Little is known about the cellular dynamics of notochord separation from the endoderm. It acts as a signaling center for proper patterning of the foregut, the neural tube, and surrounding tissues [34]. Notochord resolution from the dorsal foregut endoderm allows the endoderm to fold ventrally and form the gut tube. Once the notochord is resolved, lung buds emerge on the ventral side of the anterior foregut tube, commencing the bifurcation into the esophagus and trachea [35]. Defects in notochord resolution result in EA/TEF, potentially linked to the excess of dorsal foregut endodermal tissue attached to the notochord, leaving insufficient dorsal foregut endoderm to form the esophagus [36]. BMP signaling attenuation is required in early notochord resolution, which seems to be a prerequisite for an intact foregut separation [37]. Noggin knockout mouse models show both notochord abnormalities and EA/TEF [37].
Abnormal D/V patterning
D/V patterning defects also lead to the failure of foregut separation. The lack of differentiation of either the esophagus or the trachea may result in an abnormal foregut separation. Multiple signaling pathways (described below) are responsible for establishing proper D/V patterning during separation. The precise temporal and spatial distribution of signaling molecules within the foregut tube are critical in establishing D/V patterning. It allows foregut separation and subsequent differentiation of epithelial progenitors into the esophageal or tracheal epithelium. For instance, NKX2.1−/−, SOX2GFP/Cond, Asciz−/−, and Wnt2/2bcond are examples of genetic mouse models where the loss of D/V patterning results in EA/TEF along with tracheal and esophageal malformations [38 –41]. Table 1 shows different foregut compartmentalization defects using rodent models with variant degrees of abnormal development of the esophagus and trachea.
Abnormal septation
Foregut septation is a critical event in gut-respiratory development. Our knowledge of foregut compartmentalization comes mainly from mouse models. Mouse knockout models such as Noggin−/−, BMP4condKO, and ephrin-B2lacZ/lacZ fail to compartmentalize the foregut leading to EA/TEF and have distinctive foregut phenotypes [42 –44] (Table 1). Intact septation of the foregut may not always be linked to proper D/V patterning. BMP antagonist Noggin is normally expressed on the dorsal side of the endoderm [42]. Noggin−/− mice have EA/TEF, but the foregut has normal D/V patterning, suggesting that inhibition of BMP is not necessary for establishment or maintenance of D/V patterning. Noggin is essential for notochord resolution before compartmentalization [36]. BMP4 is normally expressed in the mesenchyme surrounding the ventral foregut where early development of the lungs occurs. BMP4condKO mice have hypoplastic lungs and absent trachea, indicating the critical role of BMP in trachea and lung development and a fistula between the esophagus and trachea. Any manipulation in BMP signaling results in esophageal and tracheal malformations. The temporal and spatial expression of BMP agonists and antagonist play critical roles at different developmental cues before, during, and after foregut separation.
Another less studied signaling pathway is the Eph/ephrin pathway. Ongoing research is revealing the role of Eph/ephrin in patterning, morphogenesis, and its self-organizing properties [45,46]. It is implicated in multiple compartmentalization events during development [44]. Eph/ephrinB2LacZ/LacZ embryos have foregut defects [44]. Loss of signaling results in loss of compartmentalization. Dravis and Henkemeyer found that Eph/ephrin signaling is active specifically at the D/V midline of the foregut [44]. It is suggested that Eph/ephrin regulates cellular behavior at the midline during compartmentalization. D/V patterning in ephrinB2 mutants is unknown. Eph/ephrin mutants could be a good model to explore the potential cellular behavior of cells at the D/V boundary.
Another example of mutants with failed foregut separation is Barx1−/− knockout mouse models. Barx1 is normally expressed in stomach mesenchyme [47]. It is required for the expression of Wnt antagonists such as secreted frizzled-related proteins 1 and 2, which suppress Wnt activity and specify stomach fate [47]. The deletion of Barx1 gene results in EA/TEF with a significantly truncated esophagus [48]. Barx1 establishes esophageal identity by suppressing Wnt signaling in the surrounding mesenchyme [49]. Interestingly, Barx1 is also detected in specific cells in the dorsal foregut mesenchyme between the nascent esophagus and trachea, where the original site of separation is located [49]. It would be interesting to identify the potential role of Barx1 in modeling cellular behavior during the initiation of separation, and the rostral movement of the septum during separation.
Abnormal vascularization development
Abnormal vessel development could lead to vascular insufficiency and physical compression in the developing foregut, resulting in growth anomalies and atresia of the dorsal part of the anterior foregut. Anomalies such as right aortic arch, aberrant subclavian artery, or vascular rings with double aortic arch, with or without cardiac malformations, are frequently found in EA/TEF patients [26]. Aorta development occurs at gestation week 3 before the bifurcation of the foregut occurs at week 4 and is surrounded by vascular structures with a bilateral distribution. Presumptive trachea and esophagus are in contact with developing heart and aortic arches that ultimately obliterate to form the aorta and great vessels [50].
Most recently, Isl1, a transcription factor involved in the multipotent and proliferative properties of the cardiac mesoderm, has been identified in the bifurcation area of the foregut tube into the esophagus and trachea [51]. This suggests that common mechanisms may involve abnormal separation of the foregut and vascular and cardiac malformations.
Abnormal muscle cell differentiation
Besides healthy vessel development, failure in normal esophageal development could be due to the developmental failure of cells/organs simultaneously developing with the esophageal epithelium. In type A EA/TEF, foregut separation is successful, but failure in the development of a continuous tube connecting the pharynx to the stomach can be observed. One hypothesis is that a localized abnormal smooth muscle malformation or failure of the enteric nerve cells to reach maturation in the esophagus could explain this phenomenon.
Signaling Networks Regulating Foregut Separation and Subsequent Epithelial Morphogenesis
During embryonic development, the intrinsic potency and the surrounding environment determine the epithelial cell fate. Cells are organized by a process known as pattern formation, which depends on regional specification caused by morphogen gradients of diffusing molecules. These molecules are involved in specific signaling pathways leading to the activation or inhibition of different transcription factors, thus determining cell fate and commitment. Mouse esophageal development studies have shown that cells within the common foregut tube are patterned along the D/V axis to give rise to the esophagus and trachea on the dorsal and ventral side, respectively. Normal foregut separation depends on transcription factors such as SOX2 and NKX2.1. Signaling molecules and pathways also involved in the bifurcation and subsequent epithelial morphogenesis of the esophagus and trachea include bone morphogenetic proteins (BMPs), Wnt/ß-catenin, hedgehog, fibroblast growth factors (FGF), and retinoic acid (RA) (Fig. 4).
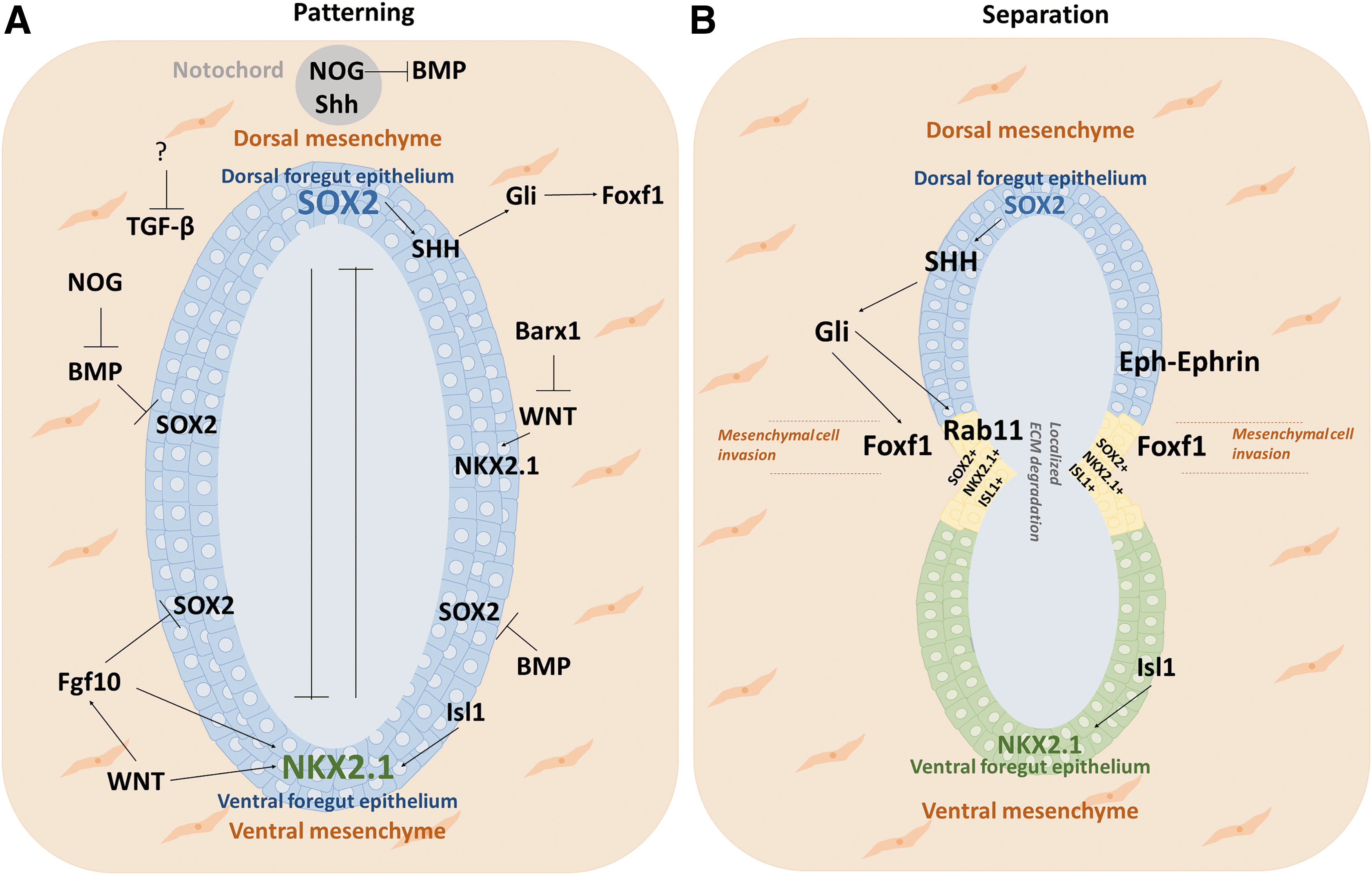
Signaling networks responsible for D/V patterning and subsequent separation of the anterior foregut into either esophageal or respiratory fate:
Transcription factors SOX2 and NKX2.1: the key players in foregut specification
SOX2, a member of the SOX family, is a conserved transcription factor essential in maintaining stem cell pluripotency [52,53]. SOX2 expression levels vary in foregut-derived organs. It is expressed on the dorsal side of the endodermal foregut tube, where the esophagus will develop, whereas NKX2.1 is expressed on the ventral side, where the trachea arises (Fig. 4A). The temporal and spatial regulation of expression of SOX2 and NKX2.1 in the dorsal and ventral regions of the foregut promote proper foregut separation into the esophagus and trachea [39]. Defects in the expression levels of SOX2 or NKX2.1 fail foregut compartmentalization. In Sox2 hypomorphic mouse models, a significant decrease in the expression of SOX2 leads to EA/TEF [39]. The critical role of SOX2 in esophageal development in mice models with anterior foregut-specific SOX2 deficiency [6,54]. In these models, the abnormal NKX2.1 expression in the dorsal region affects the normal foregut specification and separation, which results in the absence of the esophagus [6,54]. In NKX2.1 null mice, the foregut fails to separate, and the resulting tube is of esophageal identity with SOX2 expressed throughout the foregut tube [38,39]. SOX2 and NKX2.1 hypothetically repress each other; however, mechanisms involved in this repression remain unclear [1]. Kim et al. observed that Isl1 regulates the transcription of NKX2.1 [51]. In summary, studies suggest that proper D/V patterning and the expression of these transcription factors are essential to initiate separation of the foregut tube [6,14,51,54].
Following foregut separation, the esophagus is lined with squamous epithelium and the trachea with columnar epithelium [12]. The differentiation of the epithelium toward a columnar or a squamous fate is regulated by the expression of SOX2, tumor protein 63 (p63), and NKX2.1 [39,42]. SOX2 and p63 are responsible for squamous epithelial differentiation [55], while NKX2.1 is for columnar epithelium [55,56]. During esophageal development, high levels of SOX2 stimulate the formation of stratified squamous epithelium and prevent pseudostratified columnar epithelium, lining the trachea. SOX2 and p63 are expressed in the basal proliferative layer of the stratified squamous epithelium, with lower expression levels in the suprabasal layers. Decreased SOX2 expression has been linked to the downregulation of p63 [39]. p63 is involved in the proliferation of the basal layer and the stratification of the epithelium. p63 null mice have an esophagus lined with simple columnar epithelium lacking basal cells [57]. Bailey et al. recently showed the critical role of the yes-associated protein in self-renewal and stratification of the epithelium to a multilayered stratified squamous epithelium [58].
The BMP pathway promotes and spatially restricts respiratory fate in ventral anterior foregut epithelium
BMPs belong to the tumor growth factor-β (TGF-β) superfamily and are involved in stem cell proliferation and differentiation [59 –61]. Dual inhibition of BMP/TGF-β promotes the specification of anterior foregut endoderm (AFE) to dorsal foregut endoderm, which further differentiates to esophagus [7]. BMP4 is mostly expressed in the ventral mesenchyme of the foregut tube right before separation, whereas Noggin, a BMP antagonist, is highly expressed in the dorsal endoderm [42] (Fig. 4). Conditional ablation of BMP4 or BMP type 1 receptors [36,62] results in tracheal agenesis without alteration of NKX2.1 expression [36], but an expansion of SOX2 expression in the ventral endoderm [62,63]. This suggests the role of BMP4 in inhibiting digestive fate and promoting respiratory fate in the ventral anterior foregut epithelium (AFE) [62]. Noggin misexpression results in EA/TEF [64]. Noggin−/− mutants closely resemble type C of human EA/TEF [36]. Ablation of BMP7 [36] or of BMP4 function [42] in Noggin−/− mice rescued EA/TEF. Noggin is required for early notochord formation and delamination [37]. Dorsally, the action of Noggin is essential to attenuate BMP signaling in the resolving notochord, a critical event for proper separation of the foregut tube [37]. In summary, BMP4 inhibits digestive fate and restricts the site of primary bud formation in the ventral AFE, whereas Noggin plays a vital role dorsally in proper notochord delamination [37,62].
After the esophagus is established from the foregut, inhibition of BMP by Noggin is crucial to allow the stratification of the squamous epithelium (Fig. 2E). Noggin levels in the squamous epithelium decrease once esophageal fate is determined. This results in the activation of the BMP pathway in the suprabasal differentiated layer, decreasing SOX2 and p63 expression levels [65] with increasing levels of mature markers such as loricrin and involucrin. Inhibition of BMP receptor1a results in a multilayered undifferentiated epithelium, lacking suprabasal markers' expressions [66]. This suggests the dynamic role of the BMP pathway during esophageal epithelium development and maturation. First, its inhibition is necessary for the stratification of the esophageal epithelium, and then its activation is necessary for the differentiation of the proliferative basal cells into a suprabasal differentiated layer. Guyot and Maguer-Satta also showed that the inhibition of the TGF-β pathway along with the BMP pathway further promotes esophageal fate and basal esophageal cell expansion in vitro [67].
The Wnt pathway
Wnt pathway is a conserved pathway that comprises a large family of 19 proteins in humans, which include the canonical (Wnt/β-catenin dependent) or the noncanonical (β-catenin independent) pathways. Canonical Wnt pathway is necessary for respiratory fate in the anterior foregut. The Wnt ligands, WNT2 and WNT2b, are expressed in the mesenchyme surrounding the ventral foregut (Fig. 4). Suppressed Wnt signaling leads to lung and tracheal agenesis affecting NKX2.1 expression in the ventral AFE [41,68]. Wnt5a has been shown to regulate Fgf10 expression during lung formation and was shown to be upstream of Sox2, negatively regulating its expression [69]. Continuous activation of the WNT pathway leads to the loss of SOX2 expression needed for dorsal patterning and an increase in NKX2.1 expression [41]. However, the attenuation of the WNT pathway ensures tissue segregation and cellular fate during esophageal development. Esophageal fate is directed by suppression of the WNT pathway by the mesenchymal transcription factor Barx1 [49]. Wnt/Fgf crosstalk is required for the specification of the basal cells of the trachea. Wnt proteins in the respiratory epithelium regulate basal cell and tracheal cartilage development [70].
Hedgehog signaling pathway
Sonic hedgehog (Shh) signaling pathway regulates important events during development and is essential for embryonic development, adult tissue maintenance, and regeneration. Shh proteins act in a concentration- and time-dependent manner binding to canonical receptor Patched and its co-receptors, mediating downstream signal transduction, including dissociation of the transcription factors Gli2 and Gli3 proteins [71]. In the absence of an Shh signal, Gli2 is degraded and Gli3 is cleaved into a transcriptional repressor. Shh ligands are expressed in the endoderm and mediate a paracrine communication to activate Gli2 and Gli3 in the surrounding mesoderm. The role of Shh signaling in foregut development was first observed in Shh-mutant mice. In these mice, the foregut fails to separate correctly, and the resulting esophagus is hypoplastic [72]. In mice with inactivation of Gli2 and low dosage of Gli3 (Gli2−/− and Gli3+/− mutants), a hypoplastic foregut was similarly observed, with a small esophagus and hypoplastic trachea and lungs [73]. Recently, Nasr et al. demonstrated that Shh/Gli pathway plays a critical role in the tracheoesophageal separation. Shh/Gli pathway initiates medial constriction by condensation of Foxf1 mesodermal cells at the Sox2-Nkx2.1 boundary [14] (Fig. 4B). Mahlapuu et al. also suggest that SOX2/Shh signaling acts through Foxf1 to support the development of an intact esophagus during foregut separation. Loss of Foxf1 has been linked to TEF and the development of small lungs [74].
Retinoic acid and fibroblast growth factor 10
RA, an active derivative of Vitamin A, is known as a master regulator of patterning and differentiation of many organs. RA has been linked to posteriorizing the foregut and disrupting of RA results in abnormal development of posterior foregut organs [75,76]. In RARα1−/− β−/− mutants, the midline septum in the foregut tube was absent, and the resulting lungs were hypoplastic [77,78]. RA signaling has been shown to induce the expression of FGF10, an essential morphogen in tracheal differentiation [79]. FGF10 is first detected as early as E9.5 during primary lung bud formation in the anterior mesenchyme surrounding the future trachea and is highly expressed in budding sites during lung development [79,80] (Fig. 4A). FGF10 has been shown to upregulate the expression of NKX2.1 and downregulate SOX2 [39]. FGF10 has also been shown to influence the expression of other FGFs, BMPs, Wnts, and Shh during development, while regulating polarity, adhesion, and directed cell migration indicating its critical role during foregut patterning [81]. In fgf10-null mutant mice, normal separation of trachea and esophagus occurs, despite the absence of primary bud lung formation, which could mean that separation of the foregut is not initiated by primary bud induction [82]. RA has been shown to regulate the expression of Shh in the anterior side of the foregut tube before lung bud formation [83]. Crosstalks between factors such as RA, Shh, and NKX2.1 are essential for foregut separation and subsequent trachea/lung development.
The mechanisms that regulate these signaling pathways, downstream transcription factors, and modulators responsible for foregut development and generation of a mature functional esophagus remain poorly understood.
iPSCs as a Model for Understanding Organ Development
In 2006 in mice and 2007 in humans, Takahashi et al. have shown that somatic cells could be reprogrammed to generate iPSCs, which demonstrated a similar developmental capacity as embryonic stem cells (ESCs) [84,85]. Since then, iPSCs have been extensively used for research in stem cell biology and regenerative medicine, especially in gaining insights on human development [86]. Differentiation of iPSCs and their genetic manipulation provides a unique opportunity to study human embryonic development. iPSCs, like ESCs, could be specifically directed to differentiate into a specific cell fate, which involves the addition of growth factors, small molecules (activators or inhibitors) under specific culture conditions mimicking human development. Several types of cells have been derived from iPSCs [87 –92]. The first critical step in generating esophageal epithelial cells is to direct the differentiation of iPSCs to definitive endoderm (DE). Definitive endodermal cells are then guided into an anterior foregut (AF) fate from which the esophageal progenitors, as well as tracheal progenitors, arise when exposed to specific growth factors. By mimicking signaling pathways that regulate D/V patterning during development, AF cell populations can be guided into a dorsal esophageal fate or a ventral tracheal fate. The inhibition of BMP and TGF-β pathways promotes an esophageal fate, whereas the activation of BMP, Wnt, and RA pathways favors a ventral tracheal fate (Fig. 5).

Scheme representing protocols for differentiation of iPSCs/hESCs into mature esophageal and tracheal epithelium: the first critical step is the formation of the DE from pluripotent stem cells expressing SOX2, OCT3/4, and NANOG. DE is generated by activating the Nodal and BMP pathways and the expression of SOX17, CXCR4, and FOXA2. DE cell population is then directed into an AF fate by the inhibition of BMP, TGF-β, and WNT pathways. Trachea and esophagus derive from the anterior side of the foregut. Patterning of the anterior foregut tube along sagittal axis results in the formation of the esophagus on the dorsal side and the trachea on the ventral side. By mimicking the main signaling pathways involved in foregut patterning, AF can generate esophageal progenitors by the dual inhibition of BMP and TGF-β pathways and the tracheal progenitors by activation of WNT, BMP, and RA signaling pathways. Esophageal progenitors can be further matured by activating the BMP and Notch signaling pathways to generate a stratified squamous epithelium expression KRT4, KRT13, INV, and LOR. On the other hand, tracheal progenitors generate mature tracheal epithelium expressing NKX2.1 and SFTC by activating the Wnt pathway. AF, anterior foregut; hESCs, human embryonic stem cells; iPSCs, induced pluripotent stem cells; TGF-β, tumor growth factor-β. Color images are available online.
iPSC-derived esophageal organoids
It is essential to mimic organ structure and functionality to understand its development accurately. iPSC-derived cells seem to follow their intrinsic developmental queues and self-organize into “organoids” that could mimic various histological and functional aspects of real tissues and organs [93 –95]. Organoids could mimic the complexity of epithelial tissues composed of layers of interacting cells shaped into invaginations, compact folds, cysts, and tubes [96]. Several researchers have developed protocols generating organoid versions of multiple organs such as the pancreas, intestine, liver, retina, and cerebral cortex [95,97 –100]. The esophagus is probably the least studied segment of the gastrointestinal tract. Esophagus organogenesis is yet to be clearly understood, and critical modulators facilitating development and subsequent epithelial morphogenesis remain to be explored. Zhang et al. and Trisno et al. described stepwise protocols to differentiate iPSCs into esophageal organoids (Fig. 6) [6,7]. The first critical step was to induce the iPSCs into an endodermal fate. Attempts to generate DE are heavily influenced by our understanding of specific cell type formation during embryogenesis. Nodal signaling is central to the generation of the endoderm precursors [101]. The duration and magnitude of nodal signaling further specify a mesoderm or endodermal fate [102]. Trisno et al. subjected iPSCs to high concentrations of Activin A and BMP4 for 1 day, followed by Activin A only for 2 days with increasing serum concentrations [6]. Zhang et al. used high levels of Activin A, FGF2, and BMP4 for two consecutive days [7]. Both teams produced DE populations efficiently with decreased expression levels of pluripotency markers, NANOG and SOX2, and high expression of FOXA2, an endodermal marker. BMP, TGF-β, and Wnt pathways were manipulated to promote the formation of AFE. Studies in mice have shown that Noggin is expressed on the dorsal side of the foregut endoderm where the esophagus will arise, and BMP on the ventral side [42,103]. Zhang et al. used inhibitors of BMP, TGF-β, and Wnt pathways to promote the generation of SOX2+ dorsal AFE population. Increased levels of SOX2 expression are a crucial indicator of the specification of the AFE toward an esophageal fate [7]. On the other hand, Trisno et al. inhibited the BMP pathway, while activating the FGF pathway to generate AFE spheroids [6]. AFE was then directed into esophageal progenitor cells (EPCs), a cell population expressing markers for basal esophageal cells such as p63, PAX9, and FOXE1, lacking the expression of NKX2.1 [6]. EPCs were obtained by adding FGF10 to activate the FGF signaling (Trisno et al.) or inhibiting BMP and TGF-β signaling pathways (Zhang et al.) [6,7]. Esophageal organoids matured and were maintained in culture for up to 2 months. Zhang et al., however, first selected EPCs with the cell surface markers ITGb4+ EPCAM+ [7]. These iPSC-derived EPCs were able to form organoids with high efficiency, expressing regional keratins with p63+ proliferative cells on the external periphery of the organoid and cells at the center highly expressing KRT13, a squamous suprabasal epithelium marker. Zhang et al. showed that EPCs were also able to form a tubular structure when seeded in Matrigel into the kidney capsule. This tubular structure was lined with p63+ stratified epithelium with differentiated KRT13 suprabasal layer similar to the organoids in vitro [7].
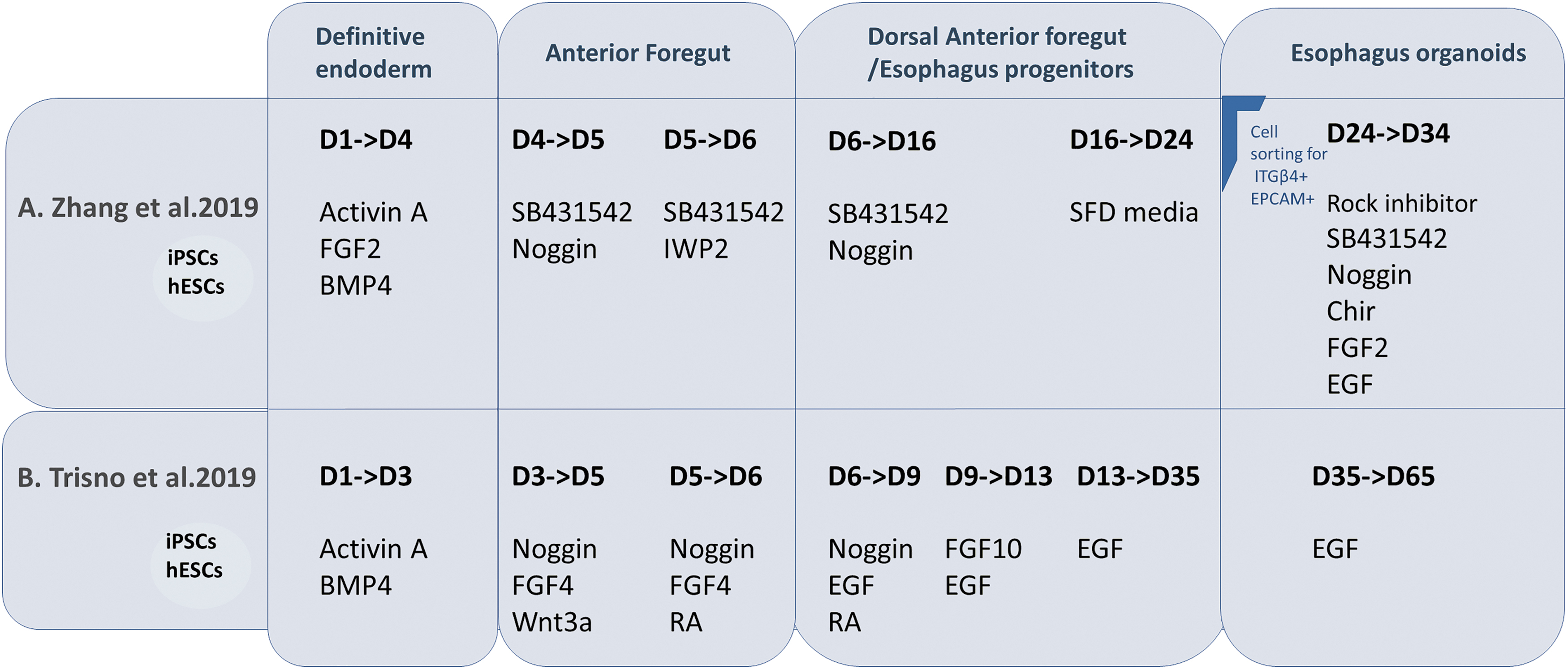
Schematic presentation of two published differentiation protocols of iPSCs/hESCs into esophageal organoids:
iPSC-derived foregut epithelium as a model to understand AF bifurcation into esophagus and trachea
In the septation model of foregut separation, the main driver of compartmentalization is the epithelium, which regulates the behavior of the mesenchyme. At the developmental stage, the endodermal foregut tube is made up of a 2–5 cell layer before bifurcation occurs (Fig. 2). Kim et al. showed a specific epithelial progenitor cell population (MECs, Isl1+ SOX2+ NKX2.1+) present at the D/V boundary of the foregut wherein separation occurs (Fig. 4B) [51]. This epithelial population is capable of generating both the esophagus and trachea. iPSCs can be used to mimic the D/V patterning of the anterior foregut tube by targeted manipulation of crucial signaling pathways, allowing the generation of the trachea and esophageal progenitors. Many investigators report iPSC differentiation protocols into trachea epithelium, suggesting that the activation of BMP, Wnt, and RA signaling pathways promote trachea fate, whereas recent findings suggest the inhibition of BMP and TGF-β to be critical for esophagus fate [7] (Fig. 5). Investigating the fate of the cells and their expression patterns after anterior foregut commitment by closely mimicking signaling pathways is an exciting approach to better understand AF bifurcation in vitro. Koike et al. used iPSCs to model hepato-biliary-pancreatic organogenesis, where the bifurcation and budding of multiple organs were demonstrated [104]. This model could be adapted to generate AF spheroids to understand bifurcation into the esophagus and trachea. In this context, using iPSCs generated from patients with EA/TEF would also allow deciphering the mechanisms involved in the malformation.
Limitations of the use of iPSC-derived tissues to study development and disease mechanisms of the esophagus must be known, despite their great potential in uncovering the earliest developmental stages. iPSCs may have a limited capacity to recapitulate healthy cell/organ development, and several discrepancies are known to exist among different iPSC clones such as epigenetic modifications, genetic variations, the source of iPSCs (including male vs. female source), and residual transgenes [86,105]. Directed differentiation strategies of iPSCs to DE remain highly variable due to different endodermal subtypes. Different methods of generating DE have distinct identities that will control the subsequent patterning into anterior and posterior foregut, or mid hindgut [106]. Furthermore, it is essential to mention that differentiation propensity among different DE is not uniform. Different DE populations could differentiate into different endodermal lineages in vitro [107]. Therefore, subtypes of DE cells may have different commitments to different lineages depending on the method used to generate endodermal cells from iPSCs. Finally, even though the compartmentalization of the AF is thought to be mainly driven by the epithelium, the interactions between foregut endoderm with splanchnic mesoderm are bidirectional [8]. This highlights the necessity to study both iPSC-derived endoderm and iPSC-derived mesoderm and their interactions to decrypt better the mechanisms of normal and abnormal development of tracheal-esophageal separation and esophageal development.
Perspectives and Conclusion
Mouse models represent an insightful tool in revealing key signaling molecules and molecular pathways regulating esophagus development. However, there are structural and physiological differences between human and mouse esophagus. Thus, it is crucial to have a representative model of human esophageal tissue, and human iPSCs are useful models of human development and disease. Two protocols of directed differentiation of iPSCs describe the generation of committed esophageal tissue [6,7]. They based their temporal manipulation of vital signals on previous findings in mouse models, revealing striking similarities between human and mouse esophagus development. Following the same strategy, directed differentiation of iPSCs reprogrammed from EA/TEF patients could help identify potential mechanisms underlying normal or impaired esophageal or tracheal development. Recreating D/V patterning across the foregut during development using 3-dimensional cultures of human iPSCs on microfluidics chips could help to mimic and understand the temporal and spatial dependent interplay between crucial signaling pathways and modulators usually occurring during human development. This approach can help understand the bifurcation process at the cellular and molecular levels, potentially deciphering the mechanisms leading to EA/TEF, a mechanism poorly understood in humans.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Funded by Fondation du CHU Sainte-Justine and Association québécoise de l'atrésie de l'æsophage (AQAO).