
Abstract
Neural crest cells (NCCs) are highly migratory multipotent cells that play critical roles in embryogenesis. The generation of NCCs is controlled by various transcription factors (TFs) that are regulated by each other and combine to form a regulatory network. We previously reported that the conversion of mouse fibroblasts into NCCs was achieved by the overexpression of only one TF, Sox10; therefore, Sox10 may be a powerful inducer of the conversion of NCCs. We herein investigated whether Sox10 functions in the direct conversion of other somatic cells into NCCs. Sox10 directly converted bone marrow-derived mesenchymal cells, but not keratinocytes, into P75+ NCCs. However, by the co-expression of four TFs (Snail1, Snail2, Twist1, and Tcfap2a) that are involved in NCC generation, but unable convert cells into NCCs, Sox10 converted keratinocytes into P75+ NCCs. P75+ NCCs mainly differentiated into glial cells, and to a lesser extent into neuronal cells. On the other hand, when Sox10 was expressed after the four TF expression, which mimicked the expression order in in vivo NCC generation, it converted keratinocytes into multipotent NCCs. These results demonstrate that Sox10 functions as an inducer of direct conversion into NCCs in cooperation with the TFs involved in NCC generation. The sequence of expression of the inducer and cooperative factors is important for the conversion of somatic cells into bona fide target cells.
Introduction
Neural crest cells (NCCs) are migratory multipotent cells that give rise to diverse derivatives. During their movement or at target tissues, NCCs differentiate into many cell types, including neurons and glial cells of peripheral sensory and autonomic ganglia, Schwann cells, melanocytes, endocrine cells, smooth muscle, and skeletal and connective tissue cells of the craniofacial complex [1]. Due to their strong migratory ability and multipotency, NCCs play critical roles in embryogenesis; abnormalities in NCCs have been implicated in various diseases, including Waardenburg's syndrome, Hirschsprung's disease, CHARGE syndrome, and DiGeorge syndrome [2].
NCCs are generated in embryos by the following sequential regulatory steps: induction, specification, delamination, and migration, which are controlled by a number of transcription factors (TFs). NCC generation is initiated during gastrulation and before the appearance of a proper neural plate [3,4]; therefore, a neural plate border, which contains a progenitor cell population that is capable of specification to NCCs, is established [5]. Zic1, Msx1, Pax3/7, and Tcfap2 induce NCC specifiers, such as FoxD3, Snail1/2, and Ets1, and premigratory NCCs are specified at the neural plate border region [6 –8]. NCC specifier genes mutually cross-regulate each other; Snail1/2 positively regulates Twist, FoxD3, and Sox9 [9], while Foxd3 promotes Sox10 expression [10]. Sox10 has been shown to activate Snail2 and itself [11]. After specification, premigratory NCCs dissolve their adherens junctions, undergo epithelial-mesenchymal transition (EMT), delaminate, and disperse from the neural tube. Snail1/2, FoxD3, Twist, and Zeb2 (Sip1) play roles in the downregulation of cadherins in these processes. Zeb2 and Snail1/2 also mediate basement membrane degradation and changes in cytoskeletal organization, which are necessary for NCC delamination [12]. Snail1/2 is mainly linked to EMT in many systems, from cancer cell lines to gastrulating embryos [13,14]. Up to NCC delamination, trunk and cranial NCCs become biased toward neuronal and ectomesenchymal lineages, respectively, and differentiate into diverse derivatives through sequential binary decisions during migration [15]. The expression of several NCC specifiers, such as FoxD3, Ets1, and Sox8/9/10, was shown to be retained and functioned in the maintenance of NC progenitors in migration [7,16 –18]. Sox10 maintains the multipotency of neural crest stem cells (NCSCs), which are retained after migration [17]. Therefore, a number of TFs are closely linked, regulate each other in temporal and spatial manners, and constitute the gene regulatory network (GRN) for the generation of NCCs [5].
Recent studies demonstrated that the introduction of genes induced a shift in the developmental fate of cells, but not through a stem cell intermediate. Fibroblasts and other cell types may be directly transdifferentiated into various cell types by a combination of cell type-specific genes. The genes induced are fundamentally selected from TFs that constitute the GRN in the development of “target” differentiated cells. For example, mouse embryonic fibroblasts (MEFs) were converted into hepatocytes following the introduction of Hnf4a, Foxa1, and Foxa2 or Foxa3 [19] or into cardiomyocytes by the forced expression of Gata4, Mef2c, and Tbx5 [20]. We previously demonstrated that MEFs were directly converted into NCCs by the forced expression of only one TF, Sox10, suggesting that Sox10 is a powerful inducer for driving the conversion of NCCs and may be feasible in other somatic cells [21]. Based on these findings, we herein investigated the direct conversion of mouse somatic cells into NCCs by Sox10. The forced expression of Sox10 converted bone marrow-derived mesenchymal cells (ST2 cells) into P75-expressing NCCs, but did not convert keratinocytes (XB2 cells) into NCCs. However, by cooperating with TFs that function in the early stages of NCC generation, namely, Snail1, Snail2, Twist1, and Tcfap2a, Sox10 successfully converted XB2 keratinocytes into P75-expressing NCCs. We also showed that the differentiation potencies of the P75+ NCCs generated changed depending on the expression patterns of Sox10 and cooperative TFs.
Materials and Methods
Cell culture
ST2 stromal cells and XB2 cells (gifted by Dr. Howard Green) were maintained in RPMI 1640 media (GIBCO, Life Technologies, Carlsbad, CA) supplemented with 5% fetal calf serum (FCS) and 50 μM 2-mercaptoethanol (2-ME) and in Dulbecco's modified Eagle's medium (DMEM) high glucose media (GIBCO) supplemented with 10% FCS, respectively. All cells were cultured at 37°C in an incubator with an atmosphere containing 5% carbon dioxide (CO2).
Retroviral transfection and NCC generation
The retrovirus vectors pMXs-GW and pMYs-GW, in which the coding regions of Snail1, Snail2, Twist1, Tcfap2a, and Sox10 were cloned, were a gift from the Human Proteome Expression Resource (HuPEX) library (HuPEX clones in HGPD,
Flow cytometry and cell sorting
To isolate P75+ cells, infected cells were dissociated by an incubation at 37°C for 6 min in Dispase II (Sanko-Jyunyaku Co., Ltd., Tokyo, Japan). After washing with staining medium (SM; phosphate-buffered saline [PBS] containing 3% FSC), infected cells were stained with an anti-P75 antibody (ab8875; Abcam, Cambridge, UK) at 4°C for 30 min. After washing with SM, cells were stained with A649-conjugated anti-Rabbit IgG (BioLegend, San Diego, CA) in the same manner. After washing with SM, the cells were suspended in SM containing 3 μg/mL propidium iodide (Calbiochem, La Jolla, CA) to eliminate dead cells. All cell sorting and analyses were performed by a FACS Aria flow cytometer (Becton, Dickinson and Company, Franklin Lakes, NJ).
Maintenance and differentiation of P75+ cells
Isolated P75+ cells were cultured under NCC culture conditions based on those described by Morrison et al. [22]: a 5:3 mixture of DMEM-low:neurobasal medium (GIBCO) supplemented with 15% Chick Embryo Extract (CEE; US Biological, Salem, MA), 1% N2 (GIBCO), 2% B27 (GIBCO), 50 μM 2-ME (Sigma-Aldrich), 35 ng/mL all-trans retinoic acid (Sigma-Aldrich), 20 ng/mL IGF-1 (R&D systems), 100 ng/mL EDN3 (Peptide Institute, Inc.), and 20 ng/mL bFGF (R&D Systems). P75+ cells were incubated at 37°C under 5% CO2. After 6 days of cultivation under NCC culture conditions, P75+ cells were differentiated into NC derivatives using the following media: regarding differentiation into neural cells, glial cells, or smooth muscle cells, P75+ cells were incubated in NCC differentiation medium, which had the same components as medium for NCC culture conditions, except that it contained 1% CEE and 10 ng/mL bFGF [22]. Regarding specific neural cell differentiation or glial cell differentiation, P75+ cells were incubated in NCC differentiation medium supplemented with 50 ng/mL bone morphogenetic protein-2 (BMP-2) (R&D Systems) or 1 nM forskolin (Sigma-Aldrich) and 1 nM Nrg-1 (R&D Systems), respectively. All differentiation cultures were performed at 37°C under 5% CO2.
Immunohistochemical analysis
Colony fixation, permeabilization, and blocking were performed as described previously [23]. Primary antibodies, diluted in 0.5% bovine serum albumin (BSA) PBS, were then added and allowed to react at room temperature. The primary antibodies used for immunohistochemistry were as follows: mouse anti-mouse neuronal class III β-tubulin (1:500; TuJ-1; COVANCE, Princeton, NJ), rabbit anti-mouse glial fibrillary acidic protein (GFAP, 1:500; Z0334; Dako Cytomation, Glostrup, Denmark), anti-mouse Nestin (1:500; Rat401; Chemicon), anti-Neurofilament (1:500; AB1981; Millipore), anti-Islet 1 (1:250; ab109517; Abcam), and anti-mouse α-smooth muscle actin (1:500; 1A4; Sigma-Aldrich). After being washed in PBS, cells were stained with secondary antibodies in the same manner. The following secondary antibodies were used: Texas Red-conjugated anti-mouse IgG (1:500; Molecular Probes, Life Technology) and Alexa Fluor 488-conjugated anti-rabbit IgG (1:500; Molecular Probes, Life Technology). Nuclei were stained with Hoechst 33258 (Sigma-Aldrich). Colonies were examined using an Olympus IX-71 fluorescence microscope (Olympus, Tokyo, Japan).
Reverse transcriptase-polymerase chain reaction (RT-PCR) analysis
Total RNA was purified using Isogen (Nippon Gene, Tokyo, Japan), and first-strand complementary DNA synthesis was performed with Superscript III (Invitrogen, Carlsbad, CA). PCR primers were as follows:
Sox10: 5′-CAGTCCGGCAAGGCAGACCC-3′ (forward) and 5′-GCAGGTATTGGTCCAGCTCAGTCAC-3′ (reverse).
FoxD3: 5′-TCTTACATCGCGCTCATCAC-3′ (forward) and 5′-TCTTGACGAAGCAGTCGTTG-3′ (reverse).
P75: 5′-ATACGGTGACCACTGTGATG-3′ (forward) and 5′-TCCACAATGTCAGCTCTCTG-3′ (reverse).
Nestin: 5′-GAAGCCCTGGAGCAGGAGAAGCA-3′ (forward) and 5′-TCCAGGTGTCTGCAAGCGAGAGTTC-3′ (reverse).
Islet 1: 5′-GGCTAACCCAGTAGAGGTGCAAAG-3′ (forward) and 5′-CTGCCAGTAGAATTAGAGCCTGGTC-3′ (reverse).
Twist1: 5′-AGCTGAGCAAGATTCAGACCCTCA-3′ (forward) and 5′-TGCAGCTTGCCATCTTGGAGT-3′ (reverse).
Id2: 5′-ACGACCCGATGAGTCTGCTCTACAA-3′ (forward) and 5′-AAGCTCAGAAGGGAATTCAGATGCC-3′ (reverse).
Gfap: 5′-GCCAAGGAGCCCACCAAACT-3′ (forward) and 5′-ATCTCCTCCTCCAGCGATTCAAC-3′ (reverse).
B-fabp: 5′-TACATGAAAGCTCTGG-3′ (forward) and 5′-AGCTGGCTAACTCTGG-3′ (reverse).
S100β: 5′-TAGGCTAGGCATTCCC-3′ (forward) and 5′-TGAATCTCAACTTCCG-3′ (reverse).
Plp: 5′-GGCTTGTTAGAGTGTTGTGCTAGA-3′ (forward) and 5′-GCAGTTCCATAGATGACATACTGG-3′ (reverse).
β-Actin: 5′-CATCCGTAAAGACCTCTATGCCAAC-3′ (forward) and 5′-ATGGAGCCACCGATCCACA-3′ (reverse).
Gapdh: 5′-CTTCACCACCATGGAGAAGGC-3′ (forward) and 5′-GGCATGGACTGTGGTCATGAG-3′ (reverse).
Results
Sox10 converts mesenchymal cells into NCCs, but not keratinocytes, into NCCs
We previously reported that the forced expression of only one TF, Sox10, directly converted MEFs into NCCs [21]. This finding revealed that Sox10 is a powerful inducer of direct conversion of cells into NCCs and suggested that the forced expression of Sox10 converts other somatic cells into NCCs. We attempted to overexpress Sox10 in mouse bone marrow-derived mesenchymal cells, the ST2 cell line, and cultured them under NCC conversion conditions (Fig. 1A). After 10 days of culture, Sox10-overexpressing ST2 cells generated P75-expressing cells (P75+ cells) (Fig. 1B). P75 (CD271, LNGFR) is expressed in human and rodent NCCs and is used as a marker for the isolation and purification of NCCs [24,25]. We isolated P75+ cells and cultured them under the NCC culture conditions previously established by Morrison et al. [22] (Fig. 1A). P75+ cells were maintained and expanded under NCC culture conditions, and expressed the NCC marker genes Foxd3, P75, Id2, and Sox10 (Fig. 1C). After 6 days of expansion under NCC culture conditions, the medium was changed into previously reported NCC differentiation medium [22]. After 3 weeks of culture, P75+ cells differentiated into TuJ1-positive neural cells (TuJ1+ N cells) and GFAP-positive glial cells (GFAP + G cells) (Fig. 1A, D). RT-PCR also showed that differentiated P75+ cells expressed the G cell marker genes Gfap and B-Fabp and the N cell marker gene Nestin (Fig. 1E). These results showed that P75+ cells generated from Sox10-overexpressing ST2 cells were NCCs, and suggested that Sox10 functions as a powerful inducer of the conversion of mesenchymal cells into NCCs.
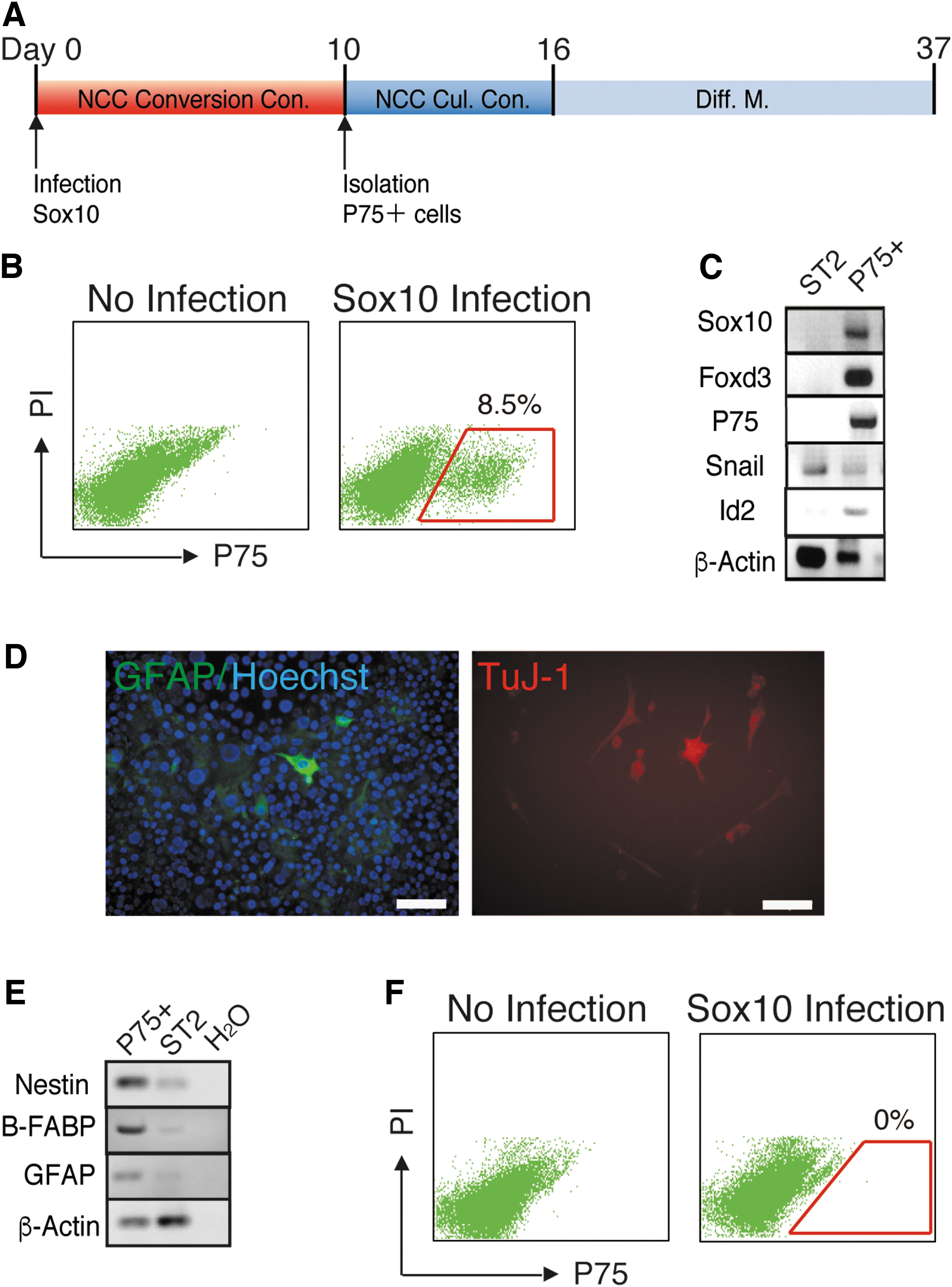
Sox10 converted mesenchymal cells into P75+ NCCs.
We also investigated whether Sox10 induces the direct conversion of mouse keratinocytes into NCCs. We overexpressed Sox10 in keratinocytes, the XB2 cell line, and cultured them under NCC conversion conditions. XB2 cells did not generate P75+ cells after 10 days of culture (Fig. 1F). Even after the duration of the culture had been extended, P75+ cells were not generated from Sox10-overexpressing XB2 cells (data not shown). This result demonstrated that Sox10 was unable to convert keratinocytes into NCCs.
Sox10 converts XB2 keratinocytes into NCCs in cooperation with Snail1, Snail2, Twist1, and Tcfap2a
Many direct conversions were previously shown to be feasible with the forced expression of a combination of TFs that functioned in the development of “target” differentiated cells [26]. We hypothesized that Sox10 will directly convert keratinocytes into NCCs in cooperation with TFs involved in NCC development. We previously identified TFs that were significantly expressed and suggested to function in the generation of NCCs in E9.5 mouse embryos [21]. Snail1, Snail2, Twist1, and Tcfap2a are well-known TFs for NCC generation and are expressed at the stage of NCC specification [5]. We selected these four TFs as cooperative factors for the direct conversion of XB2 keratinocytes into NCCs.
We investigated whether the simultaneous overexpression of Snail1, Snail2, Twist1, and Tcfap2a converted XB2 keratinocytes into NCCs. We previously reported that the individual overexpression of Snail1, Snail2, Twist1, and Tcfap2a did not convert MEFs into NCCs using Sox10-IRES Venus MEFs designed to express Venus under the control of Sox10 [21]. XB2 cells simultaneously overexpressed Snail1, Snail2, Twist1, and Tcfap2a with the retrovirus, and were cultured under NCC conversion conditions. After 10 days of culture, overexpressed XB2 cells unexpectedly generated P75+ cells (Fig. 2A). We isolated P75+ cells and cultured them under NCC culture conditions; however, the majority of P75+ cells were not maintained, and the few that survived did not differentiate into TuJ1+ N cells or GFAP + G cells in NCC differentiation medium (Fig. 2B). We also verified that the simultaneous expression of the four TFs did not convert Sox10-IRES Venus MEFs into Sox10+ NCCs (Fig. 2C). These results showed that the four TFs did not convert keratinocytes into NCCs.

Snail1, Slug, Twist1, and Tcfap2a did not convert keratinocytes into NCCs.
We then simultaneously overexpressed the four TFs and Sox10 in XB2 cells with a retrovirus, and cultured XB2 cells under NCC conversion conditions (Fig. 3A). After 10 days of culture, overexpressed XB2 cells generated P75+ cells (Fig. 3B). We isolated P75+ cells and P75 nonexpressing cells (P75− cells), and cultured them under NCC culture conditions. P75+ cells were maintained, proliferated, and formed colonies under NCC culture conditions, whereas P75− cells were not (data not shown). The colonies that P75+ cells generated were large and many cells migrated from these colonies, suggesting the strong proliferation and migratory abilities of P75+ cells (Fig. 3C). The P75+ cells that were maintained expressed the NCC marker genes Sox10, Foxd3, P75, Twist1, and Id2 (Fig. 3D). To investigate the differentiation potencies of P75+ cells, we cultured them in medium for neural cell differentiation (N-Diff. M.) or glial cell differentiation (G-Diff. M.) after 6 days of cultivation under NCC culture conditions (Fig. 3A). After 21 days of culture in G-Diff. M, the colonies generated from P75+ cells contained GFAP + G cells (Fig. 3E, left). In N-Diff. M., a small number of colonies contained TuJ-1+ N cells, and the number of TuJ-1+ N cells in colonies was also very small (Fig. 3E, right). In the culture using NCC differentiation medium in which NCCs were able to differentiate into N and G cells, a large number of GFAP + G cells and only a few TuJ-1+ N cells were generated (Fig. 3F). In NCC differentiation medium, in vivo NCSCs and NCCs that were converted from MEFs differentiated into α-smooth muscle actin-expressing smooth muscle cells (αSMA+ SM cells) [21,22]; however, none of the αSMA+ SM cells observed in the colonies formed from P75+ cells (data not shown). RT-PCR also showed that P75+ cells cultured in G-Diff M. expressed the G cell marker genes Gfap, B-Fabp, S100β, and Plp, while P75+ cells cultured in N-Diff M expressed the N cell marker gene Nestin (Fig. 3G). These results suggested that the overexpression of Snail1, Snail2, Twist1, Tcfap2a, and Sox10 converted XB2 cells into P75+ NCCs; however, their differentiation potency was for G cells, not multipotency.
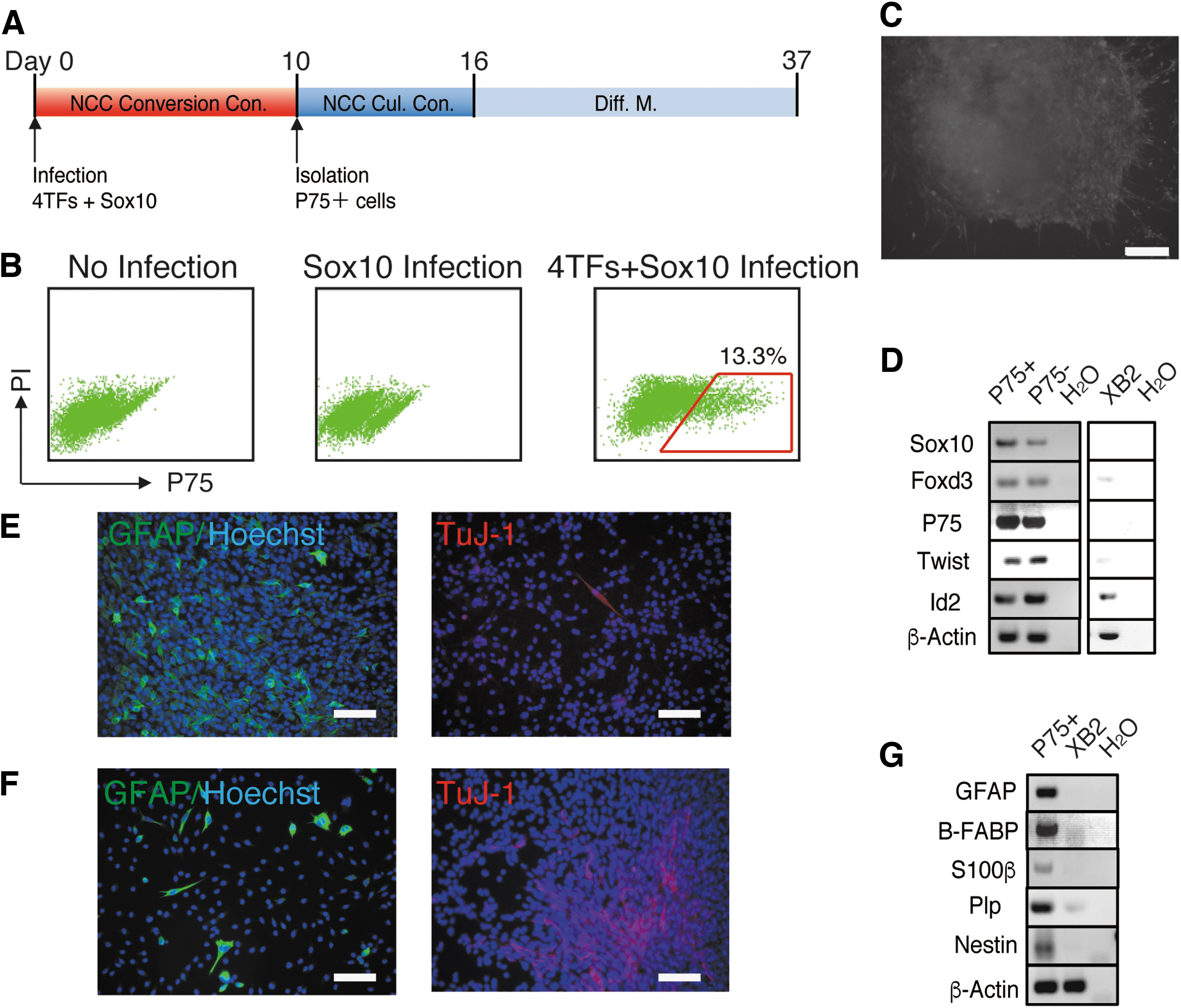
Sox10 with four TFs converted keratinocytes into P75+ NCCs.
Sox10 overexpression after the expression of Snail1, Snail2, Twist1, and Tcfap2a converts XB2 keratinocytes into multipotent NCCs
The simultaneous overexpression of Snail1, Snail2, Twist1, Tcfap2a, and Sox10 did not convert XB2 keratinocytes into multipotent NCCs. Therefore, we hypothesized that recapitulating the expression sequence of TFs in NCC generation may directly convert keratinocytes derived from the ectoderm into multipotent NCCs. In the embryo, NCCs are induced in the ectodermal germ layer during gastrulation. In in vivo NCC development, Tcfap2a plays roles in the identification of a non-neural ectoderm and neural plate border from which NCCs are generated in early embryogenesis, and Snail1, Snail2, and Twist1 specify premigratory NCCs in the neural plate border, showing that Tcfap2a, Snail1, Snail2, and Twist1 function in the early stage of NCC generation [5]. In contrast to Tcfap2a, Snail1, Snail2, and Twist1, Sox10 functioned in the late stage of NCC generation [5]. Therefore, Sox10 expression after the expression of Snail1, Snail2, Twist1, and Tcfap2a appeared to convert XB2 keratinocytes into multipotent NCCs. XB2 cells overexpressed Snail1, Snail2, Twist1, and Tcfap2a and were cultured under NCC conversion conditions. After 10 days of culture, XB2 cells expressing the four TFs overexpressed Sox10 and were cultured under NCC conversion conditions (Fig. 4A). Flow cytometry showed that overexpressing XB2 cells generated P75+ cells on day 10 of the culture after Sox10 overexpression (Fig. 4B). We isolated P75+ and P75− cells and cultured them under NCC culture conditions. P75+ cells were maintained, proliferated, and generated colonies under NCC culture conditions (Fig. 4C), whereas P75− cells were not (data not shown). P75+ cells formed enlarged colonies from which many cells migrated, suggesting the strong proliferation and migratory abilities of P75+ cells (Fig. 4C). The colonies generated from P75+ cells expressed the NCC marker genes Sox10, Foxd3, P75, Twist1, and Id2 (Fig. 4D). To investigate the differentiation potencies of the P75+ cells generated, we cultured these cells in G-Diff. M., N-Diff. M., and NCC differentiation medium after 6 days of cultivation under NCC culture conditions (Fig. 4A). After 21 days of differentiation, P75+ cells generated colonies that contained GFAP + G cells in G-Diff. M. (Fig. 4E) and those that contained a large number of TuJ-1+ N cells in N-Diff. M. (Fig. 4F, upper left), in contrast to P75+ cells generated from XB2 cells simultaneously overexpressing the four TFs and Sox10. Some TuJ-1+ N cells also expressed Islet 1 (Fig. 4F, upper right). The colonies in N-Diff. M. contained cells that expressed another N marker, Neurofilament or Nestin (Fig. 4F, lower left, and right, respectively). In the culture with NCC differentiation medium, many colonies were generated and contained GFAP + G cells and TuJ-1+ N cells (Fig. 4G). It is important to note that many GFAP + G cells and TuJ-1+ N cells were generated from cells that migrated from colonies (Fig. 4G). This result suggests that the P75+ cells generated exhibited high mobility and multipotency, similar to NCCs. In the culture with NCC differentiation medium, αSMA+ SM cells were also generated in colonies (Fig. 4H), in contrast to the absence of αSMA+ SM cells in colonies that formed from P75+ cells generated with the simultaneous overexpression of four TFs and Sox10. RT-PCR showed that the N cell marker genes Islet 1 and Nestin and G cell marker genes Gfap, B-Fabp, S100β, and Plp were expressed in P75+ cells cultured in differentiation medium (Fig. 4I). These results suggest that Sox10 overexpression after the expression of four TFs converted XB2 cells into multipotent NCCs.
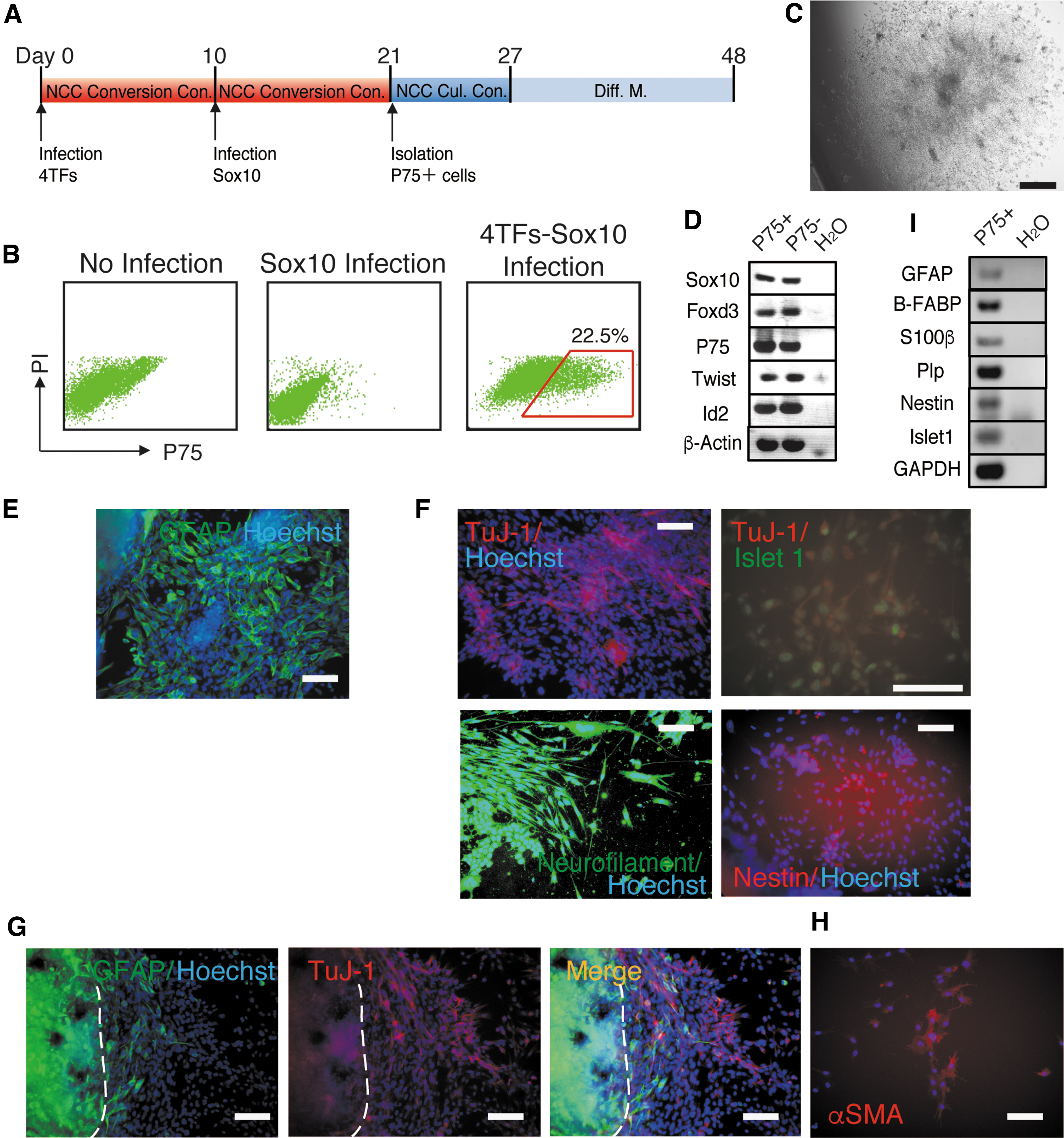
Infection with Sox10 after the expression of four TFs converted keratinocytes into multipotent P75+ NCCs.
Discussion
We previously reported that Sox10 directly converted MEFs into multipotent NCCs, suggesting that Sox10 is a powerful inducer of direct lineage conversion from MEFs into NCCs [21]. In this study, we showed that Sox10 directly converted mouse bone marrow-derived ST2 mesenchymal stroma cells, but not XB2 keratinocytes, into NCCs. This result indicated that Sox10 functioned alone as an inducer of the direct conversion of mesenchymal cells, such as MEFs and ST2 stroma cells, into NCCs. Since NCCs have the capacity to give rise to diverse mesenchymal cell types, such as chondrocytes, osteocytes, cardiac mesenchymal cells, myoblasts, and adipocytes, mesenchymal cells may have unique transcriptional states that closely resemble NCCs. On the other hand, conversion between different types of cells, such as keratinocytes and NCCs, may require the cooperation of other TFs with an inducer. Cooperative TFs were sought from the previously reported group of TFs expressed during NCC development [21]. All stages of NCC development, namely, induction, specification, and delamination, are controlled by a number of TFs that are regulated by each other and form GRN [5,27]. In TFs, we selected Snail1, Snail2, Twist1, and Tcfap2a, which are involved in NCC generation in the early stages. Snail1 and Snail2 function as transcriptional repressors and downregulate type 1 cadherins during EMT in NCC generation. Tcfap2a acts as a general regulator of NCC specification in the neural plate border [5]. Twist1 is a part of the NCC specification module, similar to Snail1, Snail2, and Tcfap2a, and confers an ectomesenchymal bias to cranial NCCs [15]. Snail1, Snail2, Twist1, and Tcfap2a play important roles in NCC development; however, they did not function as inducers of the direct conversion into NCCs in this study (Fig. 2A, B).
XB2 keratinocytes, in which the expression of Snail1, Snail2, Twist1, Tcfap2a, and Sox10 was forced, generated P75+ cells after 10 days of culture. P75+ cells were maintained and proliferated under NCC culture conditions, and then differentiated into GFAP + G cells. P75+ cells also generated TuJ1+ N cells; however, the number of N cells was small (Fig. 3E, F). Although Sox10 is required for the generation of NCCs and the maintenance of their multipotency [17], it continues to be expressed in some NCC derivatives, such as the glial cell lineage and melanocytes, after differentiation and regulates their terminal differentiation [28,29]. The forced expression of Sox10 in corporation with Nkx6.2 and Olig2 or with Zfp536 and Olig2 was shown to directly convert mouse fibroblasts into oligodendrocyte progenitor cells [30,31], suggesting that Sox10 also plays a role as an inducer of glial cell lineage conversion. In this study, P75+ cells generated with the overexpression of Sox10 together with Snail1, Snail2, Twist1, and Tcfap2a mainly differentiated into G cells (Fig. 3E, F). Although Sox10 functioned as an inducer of direct conversion into NCCs, it also appeared to contribute to the lineage restriction of glial cells.
On the other hand, the overexpression of Sox10 after the expression of Snail1, Snail2, Twist1, and Tcfap2a also generated P75+ cells equivalently differentiated into TuJ1+ N cells and GFAP + G cells (Fig. 4E–G). In this conversion, we recapitulated the gene working sequence in NCC development; Snail1, Snail2, Twist1, and Tcfap2a function in the identification and specification of premigratory NCCs before Sox10 acts in the generation of NCCs. Since keratinocytes are derived from the ectoderm, the expression of TFs along the NCC developmental sequence may have converted keratinocytes into multipotent NCCs, as if NCCs were generated from the dorsal neuroectoderm in the embryo. On the other hand, Snail, Slug, and Twist1 function in EMT [32] and the forced expression of Snail or Twist1 induced EMT in mammary gland cells [33]. Although we did not investigate whether EMT was generated by the 4 TFs in this study, the four TFs may have converted XB2 keratinocytes into mesenchymal-like cells by EMT, and Sox10 converted mesenchymal-like cells into NCCs. Further studies are warranted to clarify these issues.
The TuJ1+ N cells generated from P75+ cells by the overexpression of Sox10 after the expression of four TFs expressed Nestin and Neurofilament, which are immature neuronal cell markers (Fig. 4F), but did not express mature sensory neuronal cell markers, such as Peripherin (data not shown), showing that P75+ cells only generated immature N cells in NCC differentiation culture. The conversion for P75+ cell generation was performed by the constitutive expression of Sox10 and four TFs in XB2 keratinocytes using a retrovirus. The constitutive expression of these factors may inhibit mature neuronal differentiation. In the differentiation of other NCC derivatives, only a small number of αSMA+ SM cells and no melanocytes or ectomesenchymal derivatives were generated from P75+ cells (Fig. 4H, data not shown). This result also suggests that constitutive expression suppresses or inhibits the differentiation of other NCC derivatives. The inducible expression of these factors, similar to the usage of the Tet on/off expression system, is considered to be necessary for the mature differentiation of NCC derivatives.
We used P75 as a marker for the identification, isolation, and purification of NCCs, while the broad expression of P75 outside of the NCCs has also been observed [25]. We utilized P75 for the following reasons: there is currently no optimal marker other than P75 for the identification and isolation of mouse NCCs with flow cytometry. The expression of P75 covers all NCCs, namely, trunk, cranial, and enteric NCCs [34,35], and P75 has been utilized for the purification of the NCCs [22,36]. Furthermore, even if P75-expressing cells contained cells other than NCCs, non-NC cells were unable to grow under NC culture conditions. We found that NC culture conditions maintained and expanded converted NCCs, but not cells other than NCCs. Noninfected cells, P75− cells and, P75+ cells generated from XB2 cells infected four TFs were not maintained under NC culture conditions (Fig. 2A, B).
Although P75− cells unexpectedly expressed some NCC marker genes in RT-PCR (Figs. 3D and 4D), P75− cells were not maintained under NC culture conditions (data not shown). P75− cells may be partially or incompletely converted NCCs. Partial or incomplete conversions were previously reported in the case of reprogramming into induced pluripotent stem (iPS) cells [37,38].
We previously demonstrated that MEFs were converted into NCCs by the expression of Sox10, c-Myc, and Klf4 [21], whereas XB2 keratinocytes in this study were converted into NCCs by the expression of Sox10 and four TFs without c-Myc and Klf4. Although MEFs were also converted into NCCs by the sole expression of Sox10, the NCCs generated poorly expanded in the culture without the support of c-Myc and Klf4, which played roles in stimulating cell cycles and proliferation. On the other hand, XB2 cells were converted into expanding NCCs by the expression of Sox10 and four TFs without the support of c-Myc and Klf4. Therefore, four TFs may play the role of c-Myc and Klf4 or the high proliferative properties of XB2 cells may enable the conversion to expanding NCCs.
The mechanisms responsible for direct conversion into NCCs have not yet been elucidated in detail; however, Sox10 functions as a powerful inducer of the conversion methods used in this study. Generated NCCs altered their differentiation potency depending on the order of Sox10 expression in the direct conversion. Since the forced expression of Sox10 after the expression of Snail1, Snail2, Twist1, and Tcfap2a converted XB2 keratinocytes into multipotent NCCs, similar to in vivo NCCs, the forced expression of TFs in the working sequence of developmental stages may be important for direct conversion into bona fide target cells. Although we did not clarify whether Snail1, Snail2, Twist1, and Tcfap2a in the direct conversion played the same roles as those in in vivo NCC development, namely, the identification of a non-neural ectoderm, specification of the premigratory NCCs, and the generation of EMT, further studies to elucidate the mechanisms underlying this direct conversion may provide novel insights into the molecular mechanisms contributing to in vivo NCC development.
Footnotes
Acknowledgments
We thank Dr. Howard Green (Department of Cell Biology Harvard Medical School) for providing XB2 cells, and the other members of our laboratory for their discussions and critical reading of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Gifu University Graduate School of Medicine Research Grant Program and by the JSPS KAKENHI grant number JP18K06826.