
Abstract
Inflammatory cytokines such as interleukin-1α (IL-1α) are increased in teeth with periapical lesions. Primary teeth with periapical lesions have a propensity for accelerated eruption of the successors. In this study, we asked whether increased levels of IL-1α in the dental follicle (DF) occurring as the result of periapical lesions promote tooth eruption, possibly due to enhanced osteoclastic remodeling of DF cells (DFCs). To this end, we studied the effect and possible mechanism of IL-1α on osteogenic differentiation, osteoclastogenic activity, and matrix remodeling of DFCs. Results demonstrated that DFCs cultured with IL-1α exhibited reduced osteogenic capacity, higher osteoclastogenic activity, and stronger invasive ability. Phosphorylation of JNK and p38 was upregulated, and pretreatment with SB203580 and SP600125 reversed the effect of IL-1α on DFCs. Neonatal rats subjected to subcutaneous injection of an IL-1 receptor antagonist exhibited a reduced number in activated osteoclasts, increased expression of alkaline phosphatase and osteopontin, and delayed tooth eruption. These data support our hypothesis that increased IL-1α cytokine levels as they occur during periodontal and periapical inflammation cause osteoclastic remodeling of the alveolar socket as a requirement for tooth eruption and thus may indirectly promote the vertical eruption of teeth toward the occlusal plane.
Introduction
Tooth eruption is a process by which the tooth is elevated from its position in the alveolar bone socket to its functional position in the oral cavity. It is a complex physiological process involving bone resorption in the coronal portion of the alveolar bony crypt and osteogenesis at the base of the socket. Dental follicle (DF), a loose vascular connective tissue surrounding the tooth germ in early stages of tooth development, plays a central role in tooth eruption [1 –3]. It is reported that surgical removal of DF prevents the eruption of tooth and DF is essential for enlargement of the eruptive pathway and formation of trabecular bone [4]. And, substituting the crowns with replacements and keeping the DF intact result in normal eruption of these replicas [2]. Cells isolated from DF express a heterogeneous assortment of makers associated with stemness and can differentiate toward osteoblastic phenotype and express osteoblastic markers [5]. Moreover, stem cells in DF are also capable of differentiating into adipocytes and neurons [6]. As the tooth erupts into the oral cavity, DF forms the periodontal ligament (PDL), cementum, and alveolar bone proper.
Multiple cytokines are involved in tooth eruption. Interleukin-1α (IL-1α) may be the initial promoter of tooth eruption. Studies show that IL-1α enhances the gene expression of monocyte chemotactic protein-1 (MCP-1) and colony-stimulating factor-1 (CSF-1) [7 –9]. These two cytokines are secreted by the DF cells (DFCs) and are expressed maximally at an early postnatal (PN) age, correlating with the monocyte influx into the follicle, mediating the recruitment of mononuclear cells into DF, and stimulating osteoclast differentiation. IL-1α can also participate in the alveolar bone modeling process of tooth eruption through upregulation of receptor activator of nuclear factor-kappa B ligand (RANKL) and tumor necrosis factor alpha [10,11]. Furthermore, 31 proteins were upregulated and 7 proteins were downregulated in IL-1α-induced rat DFCs shown by proteomic analysis [12]. And null mice devoid of the IL-1 type 1 receptor (IL-1RI) gene showed delay of tooth eruption [13].
Eruption of successional teeth and the related exfoliation of their predecessors are complex processes. The successors interact with and depend on their primary teeth, and vice versa. Primary teeth with periapical lesions have a propensity for accelerated eruption of the successors, sometimes with immature root development [14,15]. And inflammatory cytokines, including IL-1α, are increased in teeth with periapical lesions [16,17]. In this study, we asked whether increased levels of IL-1α in the DF occurring as the result of periapical lesions promote tooth eruption, possibly due to enhanced osteoclastic remodeling of DFCs. The effect and possible mechanism of IL-1α on osteogenic differentiation, osteoclastogenic activity, and matrix remodeling of DFCs were investigated. The study will enhance our understanding of the process of tooth eruption regulated by IL-1α through DFCs and facilitate the clinical treatment for guiding the normal eruption of permanent tooth.
Materials and Methods
Animals and tissue preparation
Sprague-Dawley (SD) rats used in this experiment were commercially purchased from the experimental Animal Laboratory of Sichuan University. All experimental procedures were approved by the Ethics Committee of West China Hospital of Stomatology, Sichuan University, Chengdu, China (WCHSIRB-D-2016-193). The first mandibular molars were removed from embryo (E) rats at E16.5 and E18.5, and newborn rats at the age of PN days 1, 10, and 20. The tissues were then fixed in freshly prepared 4% paraformaldehyde for 24 h at 4°C and decalcified by soaking in 10% ethylenediaminetetraaceticacid in phosphate-buffered saline (PBS) for 30 days. Afterward, fully decalcified samples were dehydrated in a graded ethanol series and embedded in paraffin. Paraffin sections (5 μm thick) were mounted on poly-
Cell isolation and culture
DFCs from rats were isolated and cultured according to a method described in previous study [18]. Primary DFCs were cultured in 25-cm2 plastic flasks with the standard medium consisting of minimum essential medium alpha basic (1 × , α-MEM; GIBCO), 10% fetal bovine serum (FBS; GIBCO), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C maintained in 5% CO2. Medium was refreshed every 3 days. Cells were digested with 0.25% trypsin (GIBCO) and routinely passaged. Cells of the third passage were used. In some experiments, cell cultures were treated with IL-1α alone or 1 μM SB203580/10 μM SP600125 (Selleck) for 2 h before IL-1α treatment.
Identification of DFCs—immunofluorescence staining and transmission electron microscopy
To identify the DFCs, immunofluorescence staining was applied to characterize the expression of intracellular molecules. Antibodies included mouse anti-cytokeratin 14 (CK14, 1:100; Abcam) and mouse anti-vimentin (1:200; Millipore). Transmission electron microscopy (TEM) was adopted to detect the homogeneous electron-dense granules. These procedures were performed according to the previous report [18].
Stem cell properties of DFCs—flow cytometric analysis
Approximately 1 × 106 DFCs were incubated with fluorescein isothiocyanate (FITC) anti-CD90, FITC anti-CD45, FITC anti-CD44H, FITC anti-CD29, and FITC anti-CD11b/c (Biolegend) according to the manufacturers' protocols. Flow cytometry (FCM) was carried out using the Attune NxT Acoustic Focusing Cytometer (Thermo Fisher Scientific, Waltham, MA). The results were analyzed by FlowJo v10 software (Becton, Dickinson and Company.).
Cell proliferation assay (cell counting kit-8 and colony forming)
To determine the most suitable concentration of IL-1α, DFCs were seeded into 96-well plates (Becton, Dickinson and Company) at a cell density of 1 × 103 cells/well for 24 h. Cells were treated with 1, 5, 10, 25, and 50 ng/mL IL-1α (Escherichia coli-derived; PeproTech, Rocky Hill, NJ). After treatment, the cells were enumerated using a cell counting kit-8 (CCK-8; Dojindo, Tokyo, Japan) according to the manufacturer's instructions every other day. Colony forming was further applied to verify that the chosen concentration of IL-1α according to the result of CCK-8 had no effect on cell proliferation. It was performed as described previously [19]. Experiments were performed in triplicate.
Osteogenic differentiation
A total of 1 × 105 DFCs were seeded into each well of a six-well plate. Osteogenic-inducing medium containing 10% FBS, 5 mM
Transwell migration assay
Transwell migration assay was used to determine the invasion ability of DFCs under treatment of IL-1α. It was conducted as previously described [19]. Briefly, DFCs were seeded in upper inserts with serum-free medium, while α-MEM containing 10% FBS was placed in the lower chamber as a chemoattractant. The upper inserts in the experimental group contained 5 ng/mL IL-1α. After 24 h of incubation, the invaded cells were fixed with 4% paraformaldehyde and stained with crystal violet, and then counted under a phase contrast microscope. Experiments were performed in triplicate.
RNA extraction and qRT-PCR
By using Trizol Reagent (Invitrogen, Carlsbad, CA), RNA was split, collected, purified, and extracted according to the manufacturer's protocol. Synthesis of cDNA was performed with a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific). qRT-PCR was carried out using ABI7300 real-time PCR System (Applied Biosystems, Inc.). Primer sequences for ALP, runt-related transcription factor 2 (RUNX2), and osteopontin (OPN), RANKL, osteoprotegerin (OPG), matrix metalloproteinase 9 (MMP9), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were synthesized by TsingKe Biotech (Beijing, China) listed in Table 1. Experiments were performed in triplicate.
Oligonucleotide Primer Sequences Utilized in the Quantitative Real-Time Polymerase Chain Reaction
ALP, alkaline phosphatase; RUNX2, runt-related transcription factor 2; RANKL, nuclear factor-kappa B ligand; OPN, osteopontin; OPG, osteoprotegerin; MMP9, matrix metalloproteinase 9; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Western blot analysis
Total cellular protein was harvested with RIPA (Beyotime Biotechnology). Western blot was conducted as previously described [19]. The primary antibodies were rabbit monoclonal to ALP (1:500; HuaAn Biotechnology), rabbit monoclonal to RUNX2 (1:1,000; Cell Signaling Technology), rabbit polyclonal to OPN (1:1,000; Abcam), rabbit monoclonal to MMP9, 1:500; HuaAn Biotechnology), rabbit polyclonal to heat shock protein 27 (HSP 27, 1:500; HuaAn Biotechnology), rabbit monoclonal to p38 mitogen-activated protein kinase phosphorylation (p-p38 MAPK), rabbit monoclonal to p38, rabbit monoclonal to JNK phosphorylation (p-JNK), rabbit monoclonal to JNK, rabbit monoclonal to ERK phosphorylation (p-ERK), rabbit monoclonal to ERK, rabbit monoclonal to p65 phosphorylation (p-p65), and rabbit monoclonal to p65 (these eight, 1:1,000; Cell Signaling Technology). Rabbit monoclonal to GAPDH (1:3,000; Abcam) was used as the internal control. The intensity of each band was calculated after normalization to GAPDH. Experiments were performed in triplicate.
In vitro osteoclast formation
Bone marrow stromal cells from 10-day-old SD rats were seeded into 12-well plates (106 cells) with 20 ng/mL CSF-1. DFCs (20,000 cells) were seeded in 12-well hanging cell culture inserts (0.4 μm) to create DFC-bone marrow stromal cell co-cultures. DFCs in the experimental group were treated with 5 ng/mL IL-1α. Six days later, the induced osteoclasts were detected by tartrate-resistant acid phosphatase (TRAP) staining. TRAP-positive giant multinucleated (≥3 nuclei) cells were considered osteoclasts. Osteo assay surface (Corning, NY) was also conducted to directly assess the osteoclast activity in vitro under the instructions. The resorption area was assessed. Experiments were performed in triplicate.
Animal experiment
IL-1 receptor antagonist (IL-1Ra) is one naturally occurring cytokine whose function is to prevent the biologic response of IL-1α by its competitive binding to IL-1RI [20]. IL-1Ra has been used to block IL-1α signaling in animal studies [21,22]. Fifteen neonatal rats from the same nest were divided into three groups, five in each group. Control group received no injection, and IL-1Ra group received subcutaneous injection of 500 ng recombinant rat IL-1Ra (E. coli-derived; R&D Systems, Biokom, Poland) every other day since the following day after birth. PBS group underwent the same procedure of IL-1Ra group, except that they got same volume of PBS. Rats were anesthetized by isoflurane inhalation. They were euthanized by decapitation after anesthesia on the 14th PN day for TRAP and immunohistochemical (IHC) staining, and on the 19th PN day for micro-computed tomography (micro-CT) analysis. Two visual fields (400 times magnification) on the lingual and buccal side of mandibular first molars were selected. Osteoclast numbers around the coronal portion of the tooth were counted. Positive IHC staining in each group was semiquantitatively determined using the Image-pro Plus software (version 6.0; Media Cybernetics, Shanghai, China), and the mean optical density (OD) was calculated.
IHC staining
The paraffin-embedded sections were incubated with one of the following primary antibodies: rabbit polyclonal to IL-1α (1:200; Cloud Clone Corp.), rabbit polyclonal to IL-1RI (1:800; Cloud Clone Corp.), rabbit monoclonal to ALP (1:100; HuaAn Biotechnology), rabbit monoclonal to OPN (1:200; Abcam), mouse monoclonal to RUNX2 (1:100; HuaAn Biotechnology), rabbit polyclonal to cathepsin K (1:200; Cloud Clone Corp.), rabbit monoclonal to MMP9 (1:100; HuaAn Biotechnology), rabbit polyclonal to p-p38 (1:100; Servicebio), and rabbit polyclonal to p-JNK (1:100; Servicebio). IHC was performed following the manual of the IHC assay kit (Zhongshan Jinqiao Biological Technology Co., Beijing, China) (streptavidin-perosidase method). The sections were developed with diaminobenzidine tetrahydrochloride. No primary antibody was applied in the negative controls.
TRAP staining
The TRAP staining of cells and paraffin sections was done as previously described [19]. TRAP-kit (Beyotime Biotechnology) was applied according to the manufacturer's instructions.
Micro-CT analysis
Micro-CT scans were conducted with SkyScan1176 (Bruker, Kontich, Belgium). The samples were scanned at 70 kV and 357 μA with 17.75 μm resolution. The final three-dimensional (3D) images were reconstructed with MIMICS 19.00 (Materialise HQ, Leuven, Belgium) and vertical dimension between the molar cusp and alveolar ridge was measured as eruption distance.
Statistical analysis
All data are presented as means ± standard deviations. The statistical significance between two groups was assessed by t-test. P < 0.05 was considered statistically significant. And the statistical significance among multiple groups was assessed by analysis of variance (ANOVA). The Bonferroni test was used to compare all pairs of means after the ANOVA. Values of P < 0.05/n were considered to have statistical significance (n means the number of groups brought into Bonferroni test). Statistical testing was performed with SPSS software (version 20.0; IBM, Armonk, NY).
Results
The expression of IL-1α and IL-1RI in rat mandibular first molar
Immunolocalization of IL-1α and IL-1RI in the first mandibular molars of SD rats from embryo and newborn rats showed that the protein was absent at bud (E16.5) and cap (E18.5) stage. At the age of PN 1, IL-1α and IL-1RI were markedly expressed in the ameloblast layer, DF, and bone around the tooth germ. There was some weak staining of IL-1α in the dental papilla, and the stellate reticulum was devoid of stain. No obvious expression of IL-1RI was detected in stellate reticulum and dental papilla. At the age of PN 10, the expression of IL-1α was still intense, especially in the ameloblast layer. And cells in DF were also stained. The results of IL-1RI showed a faint staining in the ameloblast layer, while cells in the DF showed a moderate staining. At the age of PN 20, prominent expression of IL-1RI was observed in the PDL. And relatively weaker staining of IL-1α was also detected in the PDL. There was also faint staining of IL-1α and IL-1RI in the odontoblasts and alveolar bone (Fig. 1).
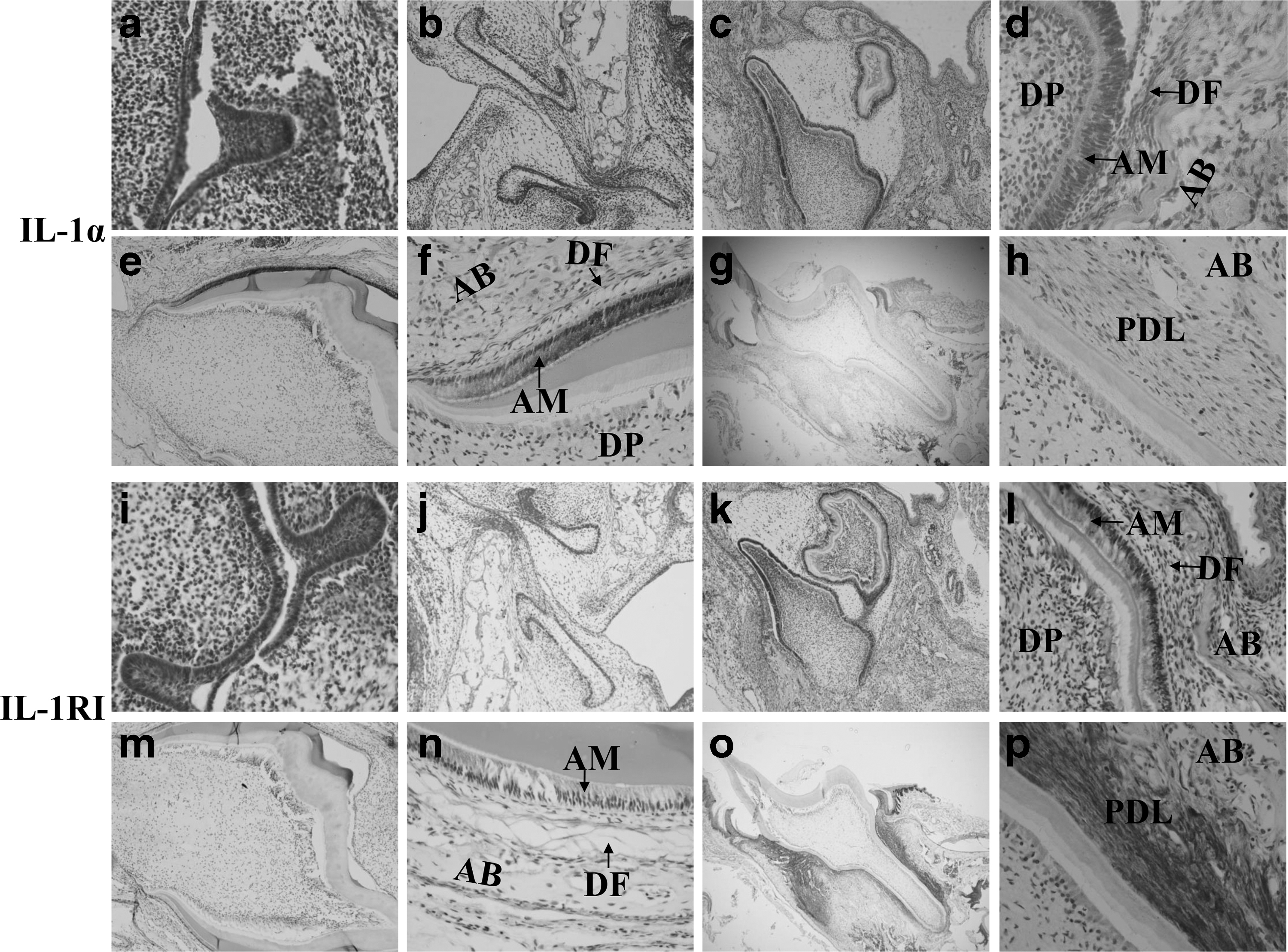
Immunolocalization of IL-1α
Culture and identification of DFCs
After an attachment period of 24 h, primary DFCs climbed out from the tissue blocks. The purified cells of the first passage displayed a fusiform shape mostly (Fig. 2a). Intracellular antigen profiles showed that DFCs were positive for vimentin, a marker of mesenchymal cells. Meanwhile, they were negative for CK14, a marker of epithelial cells (Fig. 2b). This result indicated that these cells were mesenchymal cells and were not contaminated by epithelial cells. TEM evaluations showed homogeneous electron-dense granules in DFCs, regarded as an identifying marker of DFCs (Fig. 2c).
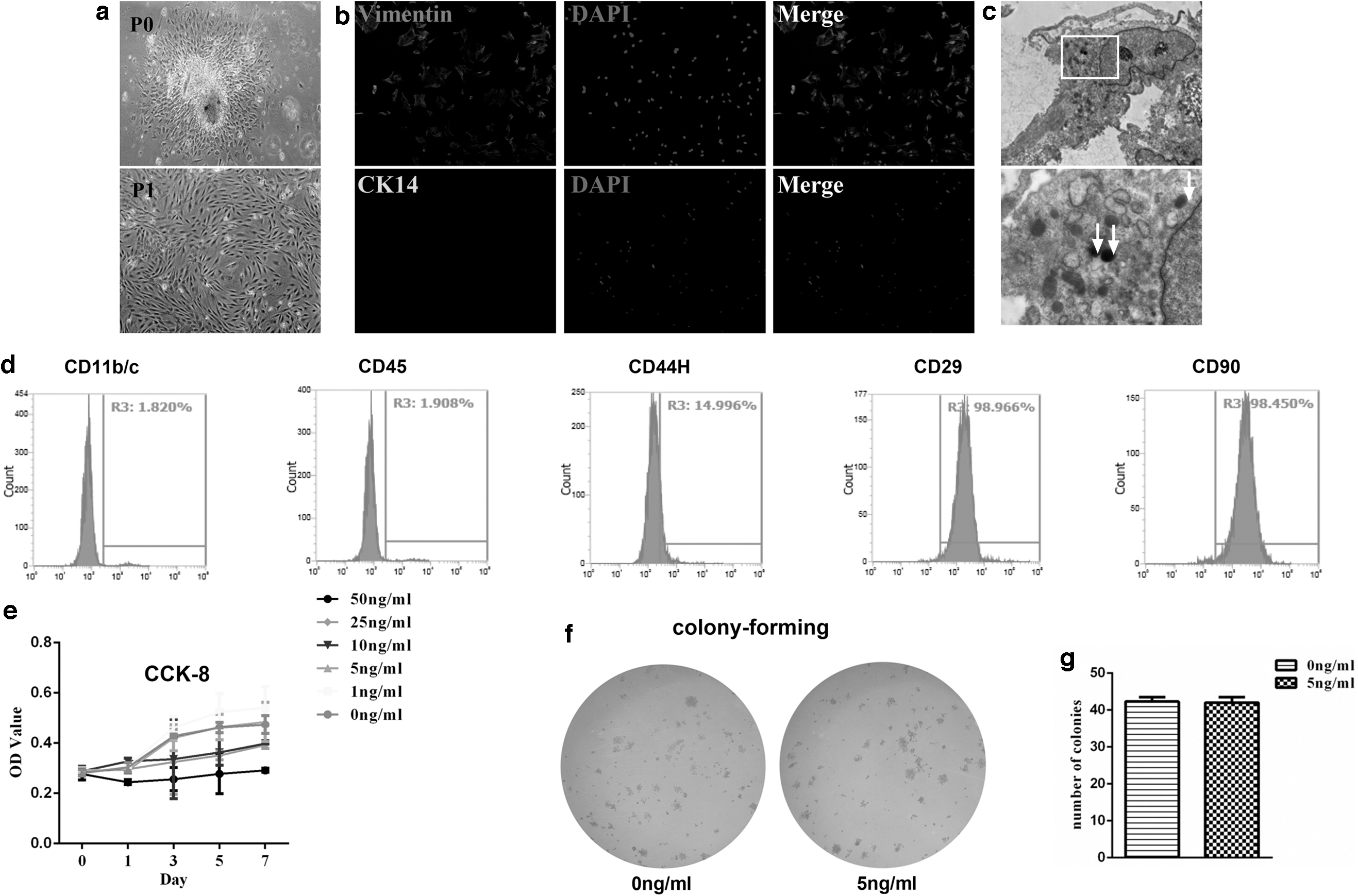
The culture, identification, and proliferation of DFCs.
FCM results demonstrated DFCs were positive for CD29 and CD90, marker of mesenchymal stem cells. And there were also some cell populations expressing CD44H. The result for CD11b/c, a marker of immune cells, such as macrophages, monocytes, granulocytes, and dendritic cells, was negative. There were also no cell populations expressing CD45, a marker of hematopoietic stem cells (Fig. 2d).
Weakened osteogenic capacities, promoted osteoclast-inductive capacities, and matrix remodeling of DFCs by IL-1α treatment
One nanogram per milliliter IL-1α promoted the proliferation rate of DFCs, while 10, 25, and 50 ng/mL IL-1α had an inhibition effect, illustrated by the CCK-8 assay. The proliferation rate of 5 ng/mL IL-1α group was similar to that of DFCs without treatment, as evidenced by the OD value in the CCK-8 assay (Fig. 2e) and similar colony-forming efficiency (Fig. 2f, g). Five nanogram per milliliter IL-1α was applied on the following study to exert as little impact as possible on the proliferation rate. To evaluate the effect of IL-1α (5 ng/mL) on the osteogenic capacity of DFCs, alizarin red staining and ALP activity assay were applied. The results displayed that DFCs under the treatment of IL-1α exhibited lower osteogenic capacity (Fig. 3a–d). qRT-PCR and western blot analysis also indicated prominently lower expression of osteogenic markers ALP, RUNX2, and OPN during DFC culture, no matter under the osteogenic induction (Fig. 3e–g) or not (Fig. 4a–c).
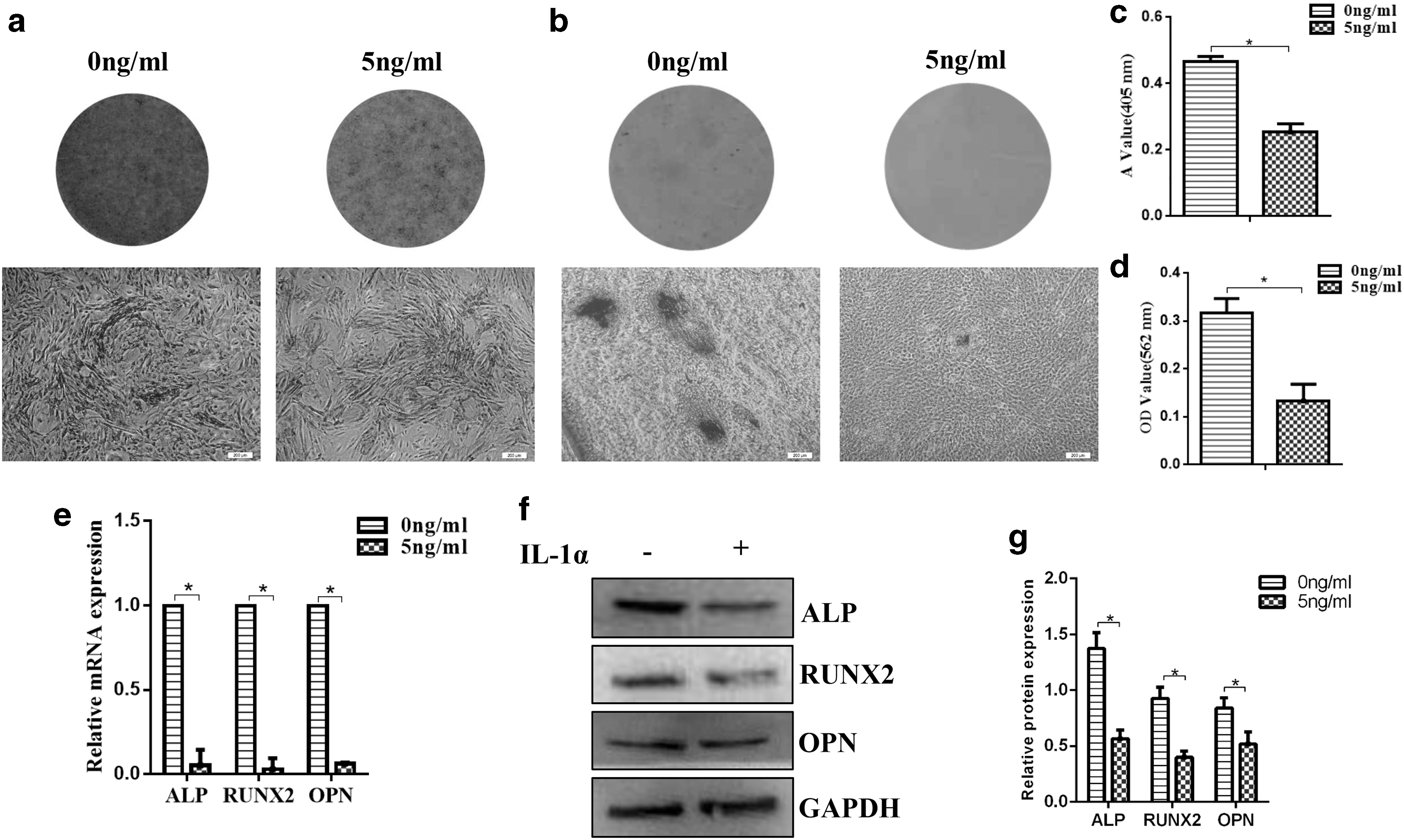
Osteogenic differentiation of DFCs under the treatment of IL-1α.
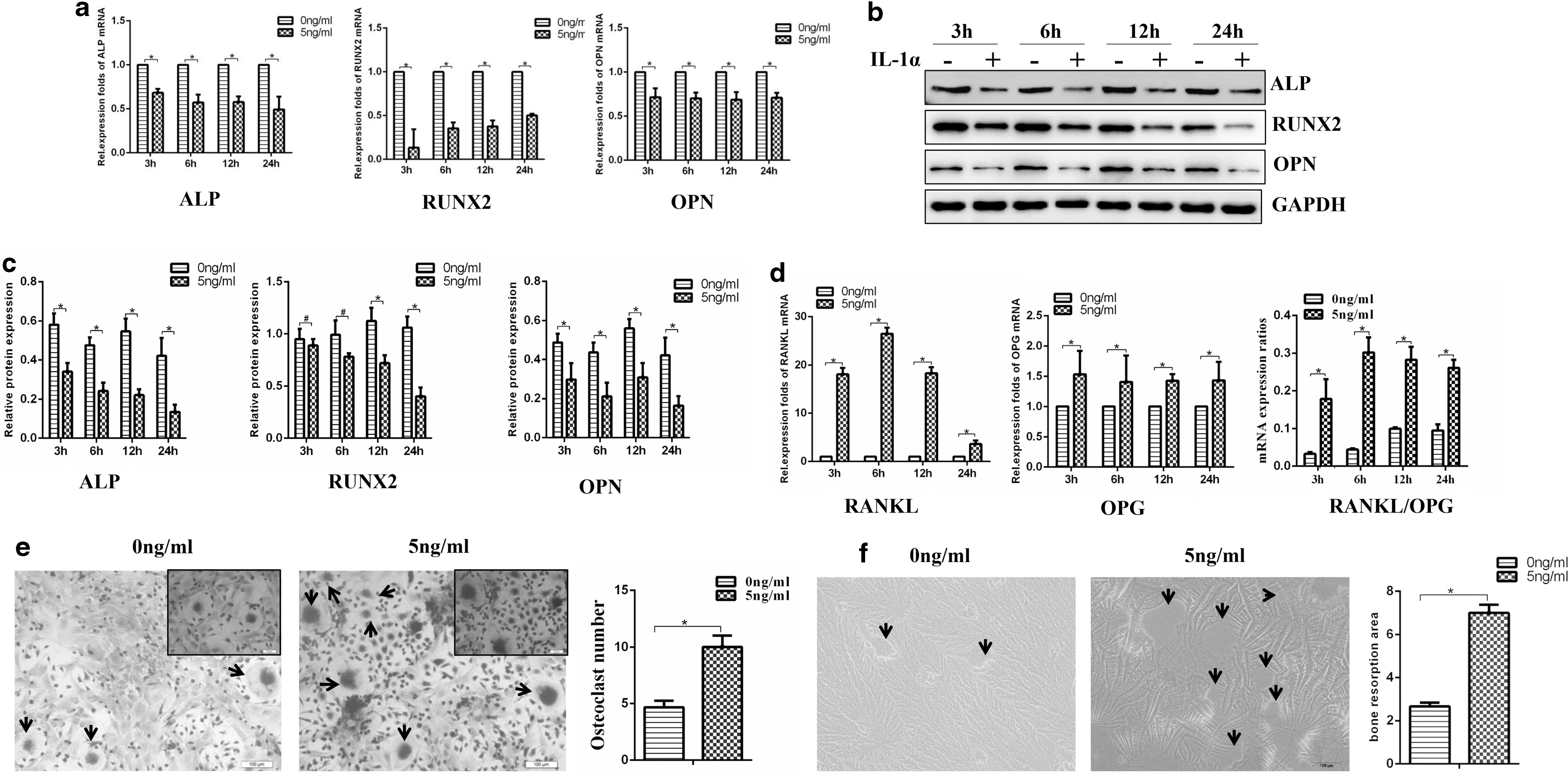
Weakened osteogenic capacities promoted osteoclast-inductive capacities of DFCs treated by IL-1α.
Data showed that the DFCs treated with IL-1α had higher gene expression levels of RANKL and OPG at 3, 6, 12, and 24 h compared with the DFCs in the control group. And the ratios of RANKL/OPG were higher in DFCs treated by IL-1α (5 ng/mL) (Fig. 4d). To further assess the effect of IL-1α on the osteoclast-inductive capacities of DFCs, DFC-bone marrow stromal cell co-cultures were performed. The TRAP-positive multinucleated osteoclasts were nearly twice in the IL-1α-treated DFCs than the group without IL-1α treatment (Fig. 4e). And more resorption area was detected on the osteo surface in the IL-1α-treated DFCs (Fig. 4f).
A modified transwell matrigel invasion assay was used to evaluate invasive ability of DFCs under the treatment of IL-1α (5 ng/mL). IL-1α promoted the invasive ability of DFCs significantly as shown in Fig. 5a and b. Consistently, the expression of MMP9 was also increased by IL-1α in DFCs, as evidenced by the qRT-PCR and western blot results (Fig. 5c–e).
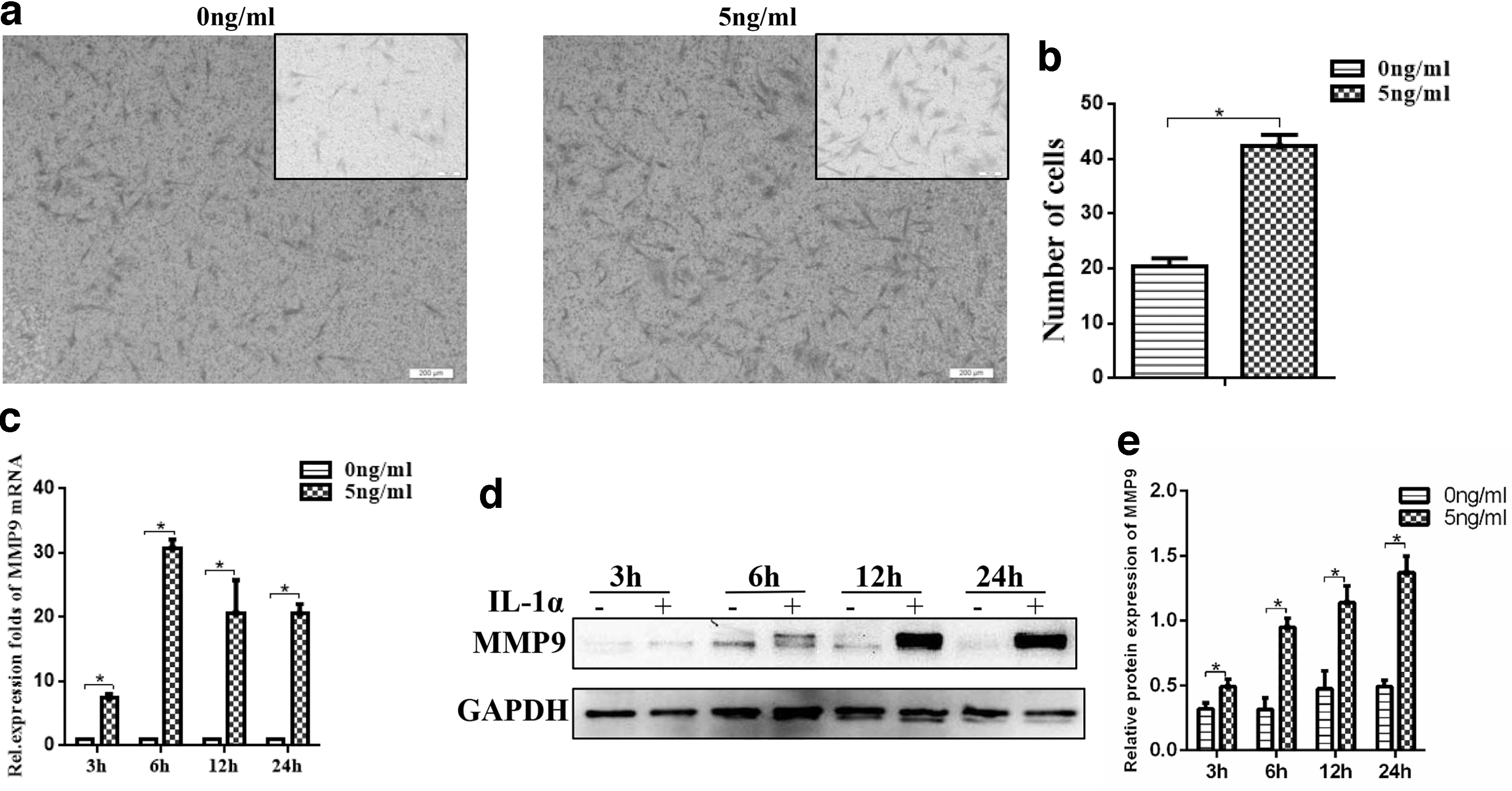
Promoted matrix remodeling capacity of DFCs by IL-1α treatment.
IL-1α participating in tooth eruption through JNK and p38 MAPK pathways
Western blot analysis showed a significant increase in HSP 27, p-p38, and p-JNK in DFCs treated with 5 ng/mL IL-1α, while the expression of p-ERK and p-p65 did not change (Fig. 6a, b).
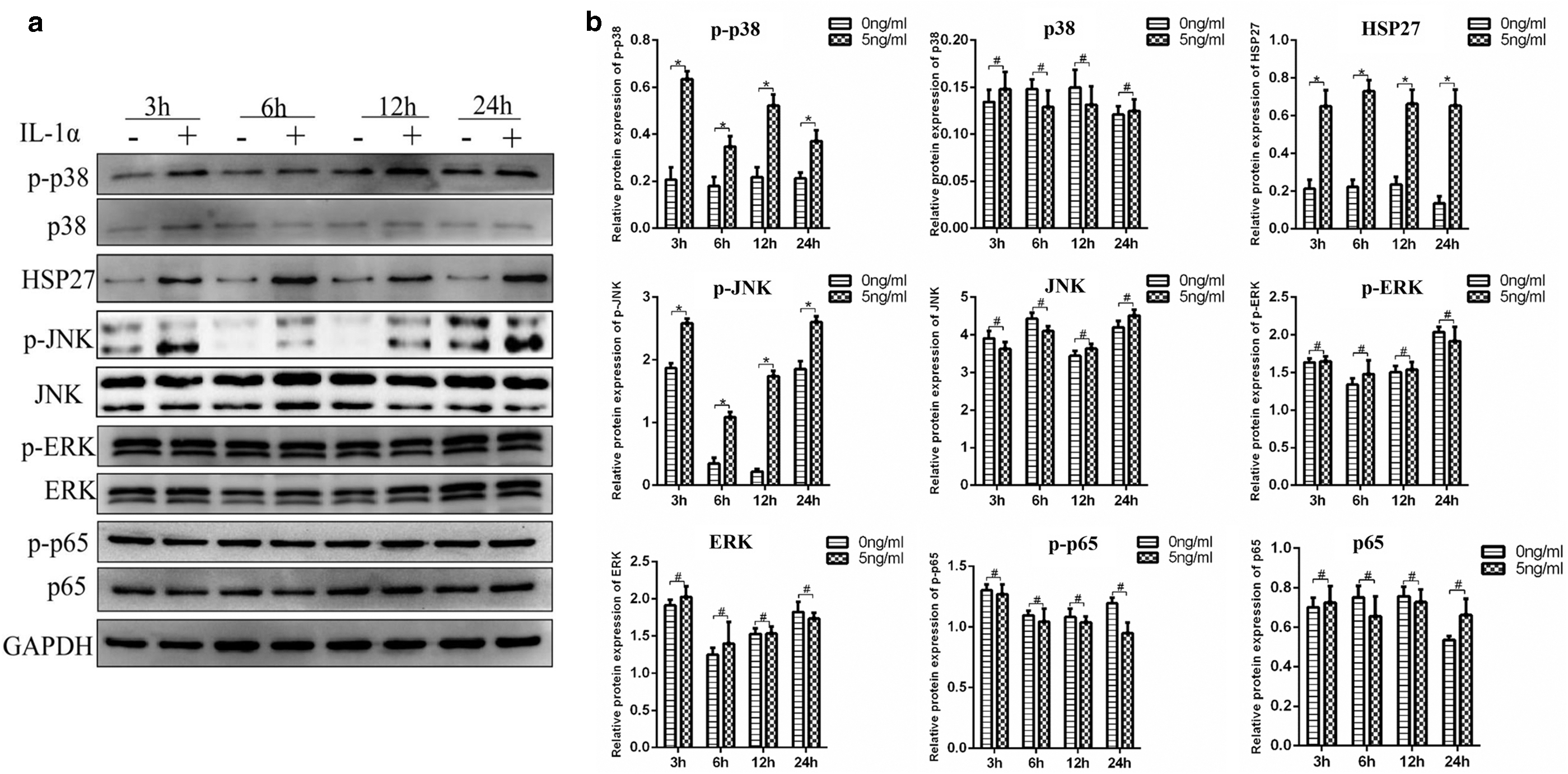
IL-1α regulates osteogenesis and osteoclastic activity of dental follicle cells through JNK and p38 MAPK pathways.
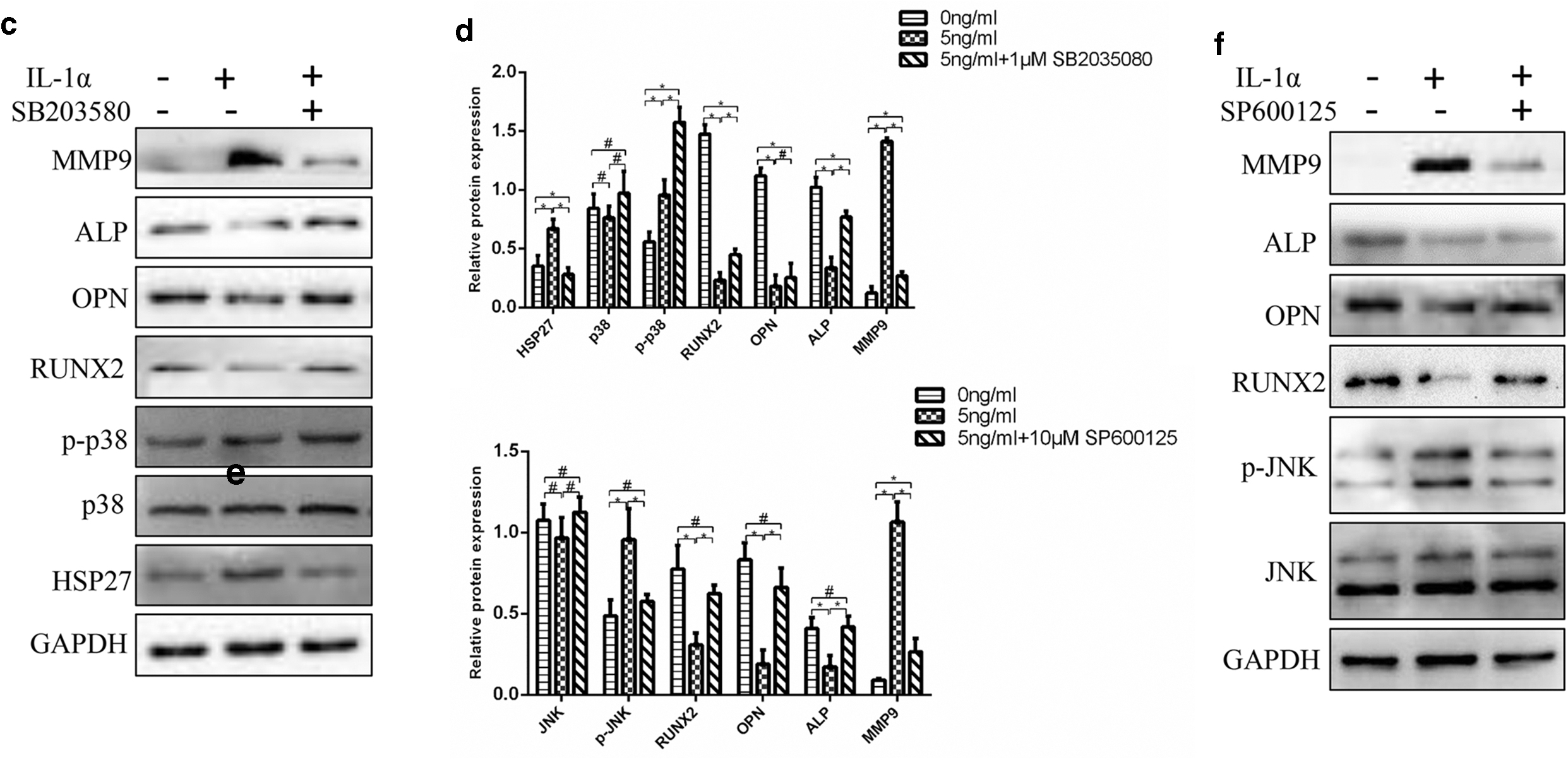
To further confirm the role of the p38 and JNK signaling pathways in the process of IL-1α regulating osteogenesis and osteoclastic activity of DFCs, the p38 MAPK inhibitor SB203580 and JNK inhibitor SP600125 were used. When pretreated with SB203580, the level of p-p38 was increased. However, the expression of HSP 27 was substantially decreased. SB203580 not only reversed the downexpression of ALP, RUNX2, and OPN but also weakened the expression of MMP9 (Fig. 6c, d). The application of SP600125 before IL-1α treatment decreased the expression of p-JNK. Similarly, MMP9, ALP, OPN, and RUNX2 also trended to come back to the level of control groups by the treatment of SP600125 (Fig. 6e, f). The effect of IL-1α was reversed by the treatment with SB203580 and SP600125. This strongly implies that p38 MAPK and JNK signaling pathway are involved in this process.
Subcutaneous injection of IL-1Ra delayed tooth eruption in neonatal rats
Subcutaneous injection of recombinant rat IL-1Ra into neonatal rats was used to inhibit the combination between IL-1α and IL-1RI. The quantification data from 3D images on PN 19 showed the eruption distance of the first molar on the lingual side was reduced in the IL-1Ra-treated rats. However, the eruption distance of the first molar on the buccal side was similar (Fig. 7a, c). The number of osteoclasts on the buccal side of the first molar was similar among these three groups, while it was significantly decreased on the lingual side in the IL-1Ra-treated group (Fig. 7b, d). The IHC results showed that the expression of cathepsin K, ALP, OPN, RUNX2, and MMP9 was similar on the buccal side of the first molar in the three groups. However, injection of IL-1Ra decreased the expression of cathepsin K, and increased the expression of ALP and OPN on the lingual side of the first molar (Figs. 7b, e and 8a, b). No difference was observed on the expression of RUNX2 and MMP9 on the lingual side of the first molar (Fig. 8b). The expression of p-p38 and p-JNK was decreased on the lingual side in the IL-1Ra-treated group, while the expression on the buccal side was similar among these three groups (Fig. 9). This was consistent with the results in vitro.
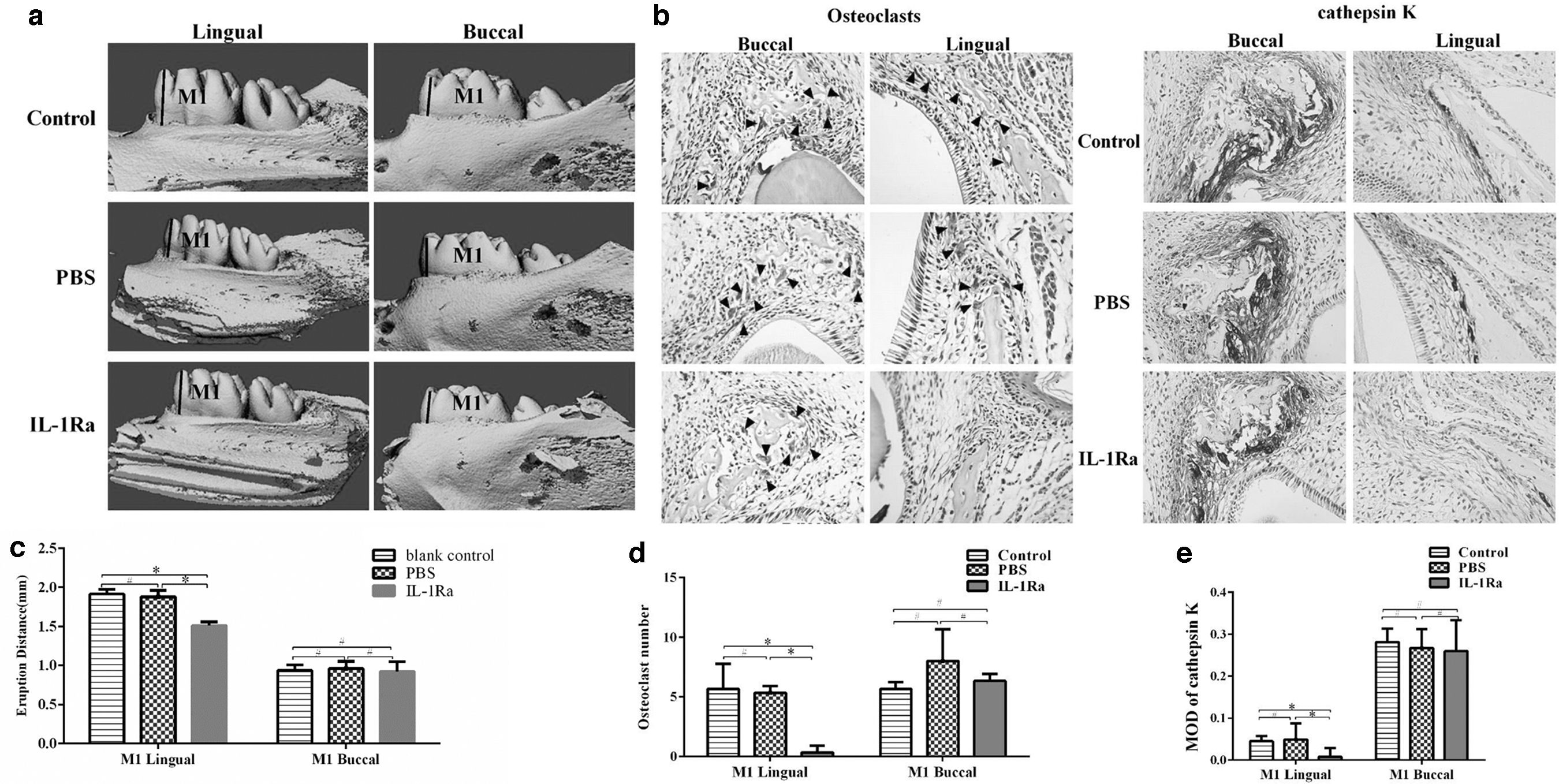
Subcutaneous injection of IL-1Ra delayed tooth eruption in neonatal rats.

Expression of ALP, OPN, RUNX2, and MMP9 illustrated by IHC staining.

Expression of p-p38 and p-JNK illustrated by IHC staining. Control, rats without injection; PBS, rats injected with same volume of PBS. IL-1Ra, rats injected with IL-1Ra. n = 5 in each group. # P > 0.05. *P < 0.05.
Discussion
Tooth eruption is a series of metabolic events in alveolar bone characterized by osteoclastogenesis, osteogenesis, and matrix remodeling. In this study, we investigated the role of IL-1α in regulation of tooth eruption through in vitro culture of DFCs and in vivo injection of IL-1Ra. Results demonstrated that DFCs cultured with IL-1α exhibited reduced osteogenic capacity, higher osteoclastogenic activity, and stronger invasive ability. Phosphorylation of JNK and p38 was upregulated, and pretreatment with SB203580 and SP600125 reversed the effect of IL-1α on DFCs. Neonatal rats subjected to subcutaneous injection of an IL-1Ra exhibited a reduced number in activated osteoclasts, increased expression of ALP and OPN, and delayed tooth eruption.
IL-1α was immunolocalized in the epithelial stellate reticulum adjacent to the DF in the first mandibular molars of rats from day 0 to 11 postnatally and DF was devoid of stain [23]. Xu et al. [24] illustrated that enamel-secreting ameloblasts, dentine-secreting odontoblasts, and osteoclasts on surfaces of alveolar bone expressed IL-1RI and -II mRNA, and no transcript was detected in stratum intermedium cells and other cells in DF, stellate reticulum, dental papilla, or pulp. However, results of immunostaining showed that IL-1RI was primarily localized in the DF [25]. And qRT-PCR analysis revealed the expression of IL-1 receptor mRNA in the DF [23]. Studies illustrated that IL-1α influenced synthesis of several genes and proteins of DFCs in vitro and in vivo [7,8,10,11,26], indicating the expression of IL-1RI in DFCs. Our research confirmed the expression of IL-1α and IL-1RI in the ameloblast layer, DF, and bone around the tooth germ at the age of PN 1 and 10. As the result showed, immunostaining of IL-1RI was observed in the PDL. And relatively weak positive IL-1α reactivity was also detected in the PDL at the age of PN 20. The IL-1RI alone on the cell surface does not work. Lossdörfer et al. [27] assumed that the small amount of IL-1α might regulate the periodontal tissue homeostasis and remodeling events involved in physiological tooth movement. Since IL-1α functions in the autocrine or/and paracrine manner and relatively higher amount of IL-1RI was expressed in the PDL, the functions quoted for IL-1α might be achieved by IL-1α secreted by other cells as well. IL-1α produced and secreted by other cells under nonphysiological conditions can bind to the IL-1RI at the cell surface to initiate an inflammatory response to bacterial irritants and subsequent periodontal tissue destruction. After the application of mechanical forces, the expression of IL-1α in the bone and PDL along the roots of orthodontically moved molars was increased, implying its role in the alveolar bone-modeling process [28].
The eruption of first mandibular molar in rats undergoes a major burst of osteoclastogenesis on PN day 3 and a minor burst of osteoclastogenesis on PN day 10. RANKL is one essential molecule required for osteoclast development, activation, and survival [29]. The expression of RANKL in rat DFCs was significantly elevated on PN days 9–11, corresponding to the minor burst of osteoclastogenesis in the alveolar bone of the first molars [10,30]. Failure of tooth eruption was observed in mice devoid of the RANKL gene [31]. OPG is a soluble decoy receptor, which can neutralize the effect of RANKL. The gene expression of OPG is reduced in the DF of rats on day 3 postnatally, correlating with the days of maximal mononuclear cell influx and osteoclast numbers [32]. The ratios of RANKL/OPG are important signals regulating osteoclastogenesis and bone resorption. Researches showed that IL-1α could upregulate RANKL and downregulate OPG in human dental pulp cells [33]. Meanwhile, IL-1α could also decrease the expression of osteoblast-related genes (ALP, RUNX2, etc.) in the MC3T3-E1 cells [34]. In our study, IL-1α upregulated ratios of RANKL/OPG, increased TRAP-positive multinucleated osteoclasts and resorption area, and decreased activity of osteogenesis, suggesting the affected osteoclastogenic and osteogenic signals of DFCs by IL-1α.
Inadequate MMP9 was presumably related to the delayed tooth eruption in patients with cleidocranial dysplasia, characterized by impaired tooth eruption in the permanent dentition stage [19]. Cerri et al. [35] showed MMP9 was expressed in multiple cells in different phases of tooth eruption and it was involved in the complex process of degradation of the eruptive pathway extracellular components. It was also located in odontoclasts and played a role in deciduous tooth resorption [36]. Previous studies showed that IL-1α could increase expression of MMPs, such as MMP 1, 3, 7, 9, and 13, during infection or the formation of endodontic and periodontal osteolytic lesions [33,37,38]. In this study, MMP9 was upregulated in IL-1α-DFCs, which might be related to abnormal tooth eruption.
Myeloid differentiation factor 88 (MyD88) is an adaptor molecule involved in IL-1α signaling, linking the upstream pathways and triggering the subsequent activation of MAPK and nuclear factor-kappa B [39 –41]. MyD88 is expressed in DFCs and knockdown of MyD88 nullifies the upregulatory effect of IL-1α on MCP-1 and RANKL gene expression [42]. In this study, we explored the downstream of MyD88 and found that a pronounced level of p38 and JNK phosphorylation was elicited by IL-1α. The activation of p-JNK was impeded by SP600125. Using SB2035080 unexpectedly improved the activation of p-p38, but the expression of HSP 27, a specific physiological substrate of p38 MAPK, was decreased. Kumar et al. [43] revealed that SB2035080 inhibited only the activity, but not the activation of p38 MAPK. It can potently inhibit the activity of p38 MAPK as demonstrated by the inhibition of the activation of HSP 27, but not its activation by upstream MAPK kinases regardless of stimuli or cell type. Further analysis showed that the upregulation of MMP9 and downregulation of ALP, OPN, and RUNX2 by IL-1α were prevented by blocking the p38 and JNK pathways. These results indicated that these two signaling pathways are involved in the process of IL-1α acting on DFCs.
Since DFCs of the mandibular first molar in rats encase the tooth germ on PN 14, and first molars erupt into the oral cavity on PN 19, these two time points were chosen for histologic and radiologic analyses after in vivo injection of IL-1Ra. In vivo IL-1Ra decreased the number of TRAP-positive osteoclasts and the expression of cathepsin K, promoted the expression of ALP and OPN, and retarded normal tooth eruption on the lingual side of the first molars in neonatal rats, indicating that IL-1Ra inhibited alveolar bone resorption and facilitated bone formation surrounding the lingual side of the first molars. The expression of IL-1RI between the buccal and lingual sides was not fully explored in this study, which might be the reason why subcutaneous injection of IL-1Ra only affects tooth eruption distance on the lingual side of the mandibular first molars significantly. More in-depth research will be conducted to obtain more comprehensive conclusions and provide guidance for clinical practice.
In conclusion, our findings indicate that IL-1α promotes tooth eruption by weakening osteogenic capacities, promoting osteoclastogenesis, and matrix remodeling of DFCs. The p38 MAPK and JNK signaling pathways are involved in the process of IL-1α influencing the activity of DFCs. These data support our hypothesis that increased IL-1α cytokine levels as they occur during periodontal and periapical inflammation cause osteoclastic remodeling of the alveolar socket as a requirement for tooth eruption and thus may indirectly promote the vertical eruption of teeth toward the occlusal plane. Getting a better appreciation of the molecular and cellular events that regulate osteoclastogenesis and osteogenesis in eruption is not only central to our understanding of how these processes occur but also is needed for ultimate development of means to control them.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the International Science and Technology Innovation Cooperation Project of Science and Technology, Department of Sichuan Province/Hong Kong, Macao and Taiwan (grant number 2019YFH0025).