
Abstract
Liver transplantation is the only radical treatment for decompensated cirrhosis, but its use is limited owing to a shortage of donors; hence, there is an urgent need for new treatments. Previously, we developed a liver-regeneration therapy using autologous bone marrow-derived mesenchymal stem cells (BMSCs), which is under clinical investigation. Cell–cell interactions between BMSCs and macrophages (Mφs) participate in the improvement of liver function and alleviation of liver fibrosis, although the associated mechanisms have not been elucidated. Therefore, in this study, we investigated phenotypic changes in Mφs caused by interactions with BMSCs, as well as the underlying mechanisms. Co-culturing lipopolysaccharide (LPS)-stimulated murine bone marrow-derived Mφs (BMDMs) with BMSCs substantially upregulated matrix metalloproteinase 9 (Mmp9), Mmp12, and Mmp13 expression, and downregulated tumor necrosis factor alpha (Tnfα) expression. To identify humoral factors involved in phenotypic changes occurring in Mφs, microarray analysis was performed with microRNAs (miRNAs) derived from extracellular vesicles in the supernatant of co-cultured BMSCs and LPS-stimulated BMDMs. We found that miR-6769b-5p was highly expressed and that transfecting miR-6769b-5p mimic upregulated MMP9 in LPS-stimulated BMDMs and downregulated Tnfα and interleukin-1 beta (Il-1β). MiR-6769b-5p expression in BMDMs was decreased by LPS stimulation but was increased by co-culture with BMSCs. Microarray and pathway analyses of gene expression in LPS-stimulated, miR-6769b-5p-transfected BMDMs revealed changes in the eukaryotic initiation factor 2-signaling pathway and decreased the expression of activating transcription factor 4 (Atf4). LPS-stimulated BMDMs exhibited increased MMP9 expression and decreased the expression of Tnfα and Il-1β by ATF4 knockdown. These findings indicate that upregulating miR-6769b-5p in BMDMs induced a fibrolytic phenotype, where MMP9 was highly expressed and inflammatory cytokine expression was decreased by the suppression of ATF4 expression. These findings imply that regulating miR-6769b-5p or ATF4 expression in BMDMs may be helpful for treating chronic liver disease.
Introduction
Liver cirrhosis is the terminal stage of chronic liver injury. In recent years, the increased use of antiviral agents has made it possible to control the progression of liver cirrhosis in patients with chronic hepatitis B and C. However, the prevalence of nonalcoholic steatohepatitis (NASH) is increasing in patients with metabolic disease and obesity. Therefore, the incidence of NASH-associated cirrhosis is expected to increase in the future [1].
Existing medical treatments for cirrhosis are limited to maintenance of the hepatic reserve function. Improvement in hepatic function is not easy to achieve, and median survival time for patients with decompensated cirrhosis is 2 years or less [2]. In addition, although liver transplantation can be expected to cure decompensated cirrhosis, its use is limited because of the lack of donors, the requirement for surgery, and the long-term oral administration of immunosuppressive drugs [3]. At present, symptomatic treatment is mostly administered to patients with decompensated cirrhosis; hence, new treatments that can improve hepatic reserve function and patient prognoses are urgently needed.
To date, we have shown that liver fibrosis was reduced in murine and canine models of liver fibrosis after administering bone marrow-derived mesenchymal stem cells (BMSCs), along with increased hepatic matrix metalloproteinase 9 (MMP9) expression and decreased tumor necrosis factor alpha (TNF-α) expression [4 –6]. Next, based on these findings, we developed a liver-regeneration therapy using autologous BMSCs cultured from bone marrow fluid, and we are currently conducting a clinical trial on patients with decompensated cirrhosis. Although the efficacy of BMSC administration has been reported in patients with liver cirrhosis [7 –11], the detailed mechanism underlying this effect remains unclear; hence, further study is needed to elucidate this mechanism to develop more effective therapeutic methods.
Mesenchymal stem cells (MSCs) act as anti-inflammatory agents and promote liver regeneration through cell–cell interactions [12,13]. In recent years, microRNAs (miRNAs) derived from extracellular vesicles (EV-miRNAs) have been garnering increasing attention as a medium for such cell–cell interactions. Exosomes are EVs that contain proteins and nucleic acids, such as miRNAs, and are transported to other cells, where they cause functional changes. MiRNAs are 21–25-nucleotide, noncoding RNAs that suppress protein production by destabilizing target mRNAs and suppressing translation. Improved liver fibrosis was observed in mouse models of liver fibrosis after administering exosomes or EVs derived from healthy mouse serum or MSCs [14,15].
Macrophages (Mφs) are not only involved in the pathogenesis of liver inflammation and fibrosis, but also play important roles in alleviating liver fibrosis through fibrolysis [16]. Although bone marrow-derived Mφs (BMDMs) recruited to the liver play a fibrogenic role in NASH [17,18], Mφs can exhibit antifibrotic effects during recovery from liver injury [17]. During the process of fibrosis resolution, Mφs can undergo a phenotypic switch from a fibrogenic phenotype characterized by TNF-α, interleukin (IL)-1β, and transforming growth factor (TGF)-β production to a fibrolytic phenotype characterized by the production of high levels of fibrolytic enzymes such as MMP9, MMP12, and MMP13, and suppressed inflammatory cytokine production [19 –22]. BMSCs have been reported to be involved in this phenotype switch [23].
Therefore, in this study, we aimed to investigate the effects of cell–cell interactions between BMDMs and BMSCs by co-culturing both types of cells and to determine the involvement of EV-miRNAs in these effects.
Materials and Methods
Animals
Animals were housed in a specific pathogen-free environment under standard conditions with a 12-h day/night cycle, with unlimited access to food and water. Male C57BL/6 mice (7–10 weeks old) were purchased from Japan SLC, Inc. (Shizuoka, Japan) and used in the experiments. All animal experiments were approved by the Animal Experiment Committee of Yamaguchi University School of Medicine (approval number 21-043).
Cell culture and co-culturing BMDMs with BMSCs
BMDMs derived from C57BL/6 mouse femurs were cultured at 37°C under 5% CO2 for 7 days in Ultra-Low attachment cell culture flasks (Corning, NY) containing Dulbecco's modified Eagle's medium (DMEM)/F12 medium with GlutaMAX supplement, 10% fetal bovine serum (Thermo Fisher Scientific, Waltham, MA), and 20% L929 conditioned medium (CM). BMSCs from C57BL/6 mouse were purchased from Cyagen (Santa Clara, CA; catalog number MUBMX-01001).
BMDMs were indirectly co-cultured with BMSCs for 24 h. For the transwell co-cultures, 4 × 105 BMDMs were seeded in six-well plates (Corning) in the lower compartments, and 2.0 × 105 BMSCs were seeded in transparent polyethylene terephthalate membrane cell-culture inserts with 0.4-μm pore (Corning) in the upper compartments. Lipopolysaccharide (LPS; Sigma-Aldrich, St. Louis, MO) was used to stimulate BMDMs at a final concentration of 100 ng/mL.
For the miRNA-mimic and small interfering RNA (siRNA) transfections, BMDMs were seeded in 24-well plates (Corning) at a density of 1 × 105 cells/well and cultured in DMEM/F12 medium with Lipofectamine RNAiMAX (Thermo Fisher Scientific) and stimulated with LPS after transfection. To evaluate the effect of BMSC-CM on miR-6769b-5p expression in BMDMs, BMDMs were plated in 100-mm tissue culture dishes (Iwaki, Shizuoka, Japan) at a density of 2 × 106 cells/dish and cultured in DMEM/F12 medium with or without 50% BMSC-CM.
Real-time polymerase chain reaction analysis
Total RNA and miRNA were extracted using the RNeasy Mini Kit and RNeasy MinElute Cleanup Kit (Qiagen, Venlo, the Netherlands) according to the manufacturer's instructions.
Complementary DNA was subsequently synthesized using ReverTra Ace qPCR RT Master Mix with gDNA Remover (Toyobo, Tokyo, Japan). Fast SYBR™ Green Master Mix (Thermo Fisher Scientific) was used for real-time polymerase chain reaction (PCR). The primer sequences are listed in Table 1. Ribosomal protein S18 mRNA expression was detected as an internal control.
Sequences of Primers Used for Real-Time Polymerase Chain Reaction
Atf4, activating transcription factor 4; Il-1β, interleukin-1 beta; Mmp, matrix metalloproteinase; Rps18, ribosomal protein S18; Tnfα, tumor necrosis factor alpha.
Complementary DNA was synthesized from miRNAs using the miRCURY LNA™ Universal RT microRNA PCR Universal cDNA Synthesis kit (Qiagen). MiRCURY LNA miRNA PCR Assays (Qiagen) were used to quantify the relative expression levels of mmu-miR-6769b-5p (catalog number YP02115393, target sequence: 5′-CCUGGUGGGUGGGGAAGAGC-3′) and U6 (catalog number YP00203907) as an internal control. The miRCURY LNA SYBR Green PCR Kit (Qiagen) was used for real-time PCR, which was performed using a Step One Plus Real-time PCR System (Thermo Fisher Scientific).
Fold changes in relative gene expression levels (relative to the control) were calculated using the 2–ΔΔCt method.
Microarray and pathway analyses
Microarray analysis was performed by Toray Industries, Inc. (Tokyo, Japan). Comprehensive analysis of mRNA from BMDMs was performed with the 3D-Gene® mouse oligo chip 24K (Toray Industries, Inc.). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of the microarray data was performed using the web-based GeneCodis tool (
Comprehensive miRNA expression analysis was performed using the 3D-Gene miRNA Labeling Kit (Toray Industries, Inc.) and the 3D-Gene Mouse miRNA Oligo Chip, version 21 (Toray Industries, Inc.). All analyzed data were scaled by global normalization. Comparisons between two groups of mouse sera and culture supernatants were performed on the basis of the global normalization ratio, which is a corrected value where the median ratio for all genes is 1 for the reference samples in each group.
To identify pathways associated with significantly differentially expressed genes (P < 0.05, fold change >1.5) compared with the control groups, the data were analyzed using Ingenuity Pathway Analysis (IPA) software (version spring 2020,
EV isolation
EVs were isolated from the culture media of co-cultured BMDMs and BMSCs using the Total Exosome Isolation reagent (Thermo Fisher Scientific) after centrifuging each sample at 2,000 g for 30 min, according to the manufacturer's instructions.
miRNA and siRNA transfections
Thirty picomoles each of mirVana™ miRNA mimics (mmu-miR-135a-1-3p, mmu-miR-8113, mmu-miR-6769b-5p, mmu-miR-7662-3p, and mmu-miR-1247-3p, Assay ID: MC12315, MC26809, MC28166, MC28052, and MC19654, respectively); Silencer Select ATF4 (Assay ID: s62689); mirVana miRNA mimic Negative Control No. 1 (NC-miR); or Silencer™ Select Negative Control No. 1 siRNA (NC-siRNA) were reverse transfected into cells using the Lipofectamine RNAiMAX reagent (Thermo Fisher Scientific) for 48 h. All miRNA mimics and siRNAs were purchased from Thermo Fisher Scientific. To analyze the transfection efficiency, the cells were transfected with the BLOCK-iT Alexa Fluor Red Fluorescent Control (Thermo Fisher Scientific) for 48 h, after which the cells were examined using fluorescence microscopy (BioRevo BZ9000; Keyence, Osaka, Japan).
Gelatin zymography
After transfecting BMDMs with different miRNA mimics for 48 h, the medium was changed to serum-free medium, and the culture supernatant was collected after an additional 24-h incubation. To detect MMP9 activation, the culture supernatant was subjected to gelatin zymography using a Gelatin Zymography Kit (Cosmo Bio Co., Tokyo, Japan) according to the manufacturer's instructions. The gel was incubated for 60 h in reaction buffer at 37°C. The total intensity of MMP activity was calculated using ImageJ software v. 1.53a (National Institutes of Health, Bethesda, MD).
Statistical analysis
Statistical evaluation of multiple groups was performed using one-way analysis of variance followed by Tukey's test or Dunnett's test. Statistical analysis of two groups was performed using Student's t-test. Statistical analysis was performed using JMP® 14 software (SAS Institute, Inc., Cary, NC). Values of P < 0.05 were considered to reflect statistically significant differences. Data are presented as mean ± standard deviation (SD).
Results
BMSCs increased MMP expression and decreased inflammatory cytokine expression in LPS-stimulated BMDMs
Indirect co-culture of mouse BMSCs and BMDMs was performed to investigate phenotypic changes of Mφs owing to the interaction with MSCs, and gene expression levels in BMDMs were evaluated. Unstimulated and LPS-stimulated BMDMs were examined because of the reported involvement of LPS in chronic liver diseases, such as NASH [25 –27]. Compared with unstimulated BMDMs, Mmp9 was significantly upregulated in LPS-stimulated BMDMs (Fig. 1A). No changes in Mmp expression levels were observed in unstimulated BMDMs. However, after co-culturing LPS-stimulated BMDMs with BMSCs, Mmp12 and Mmp13 were significantly upregulated and Mmp9 expression showed a tendency to increase (Fig. 1A).
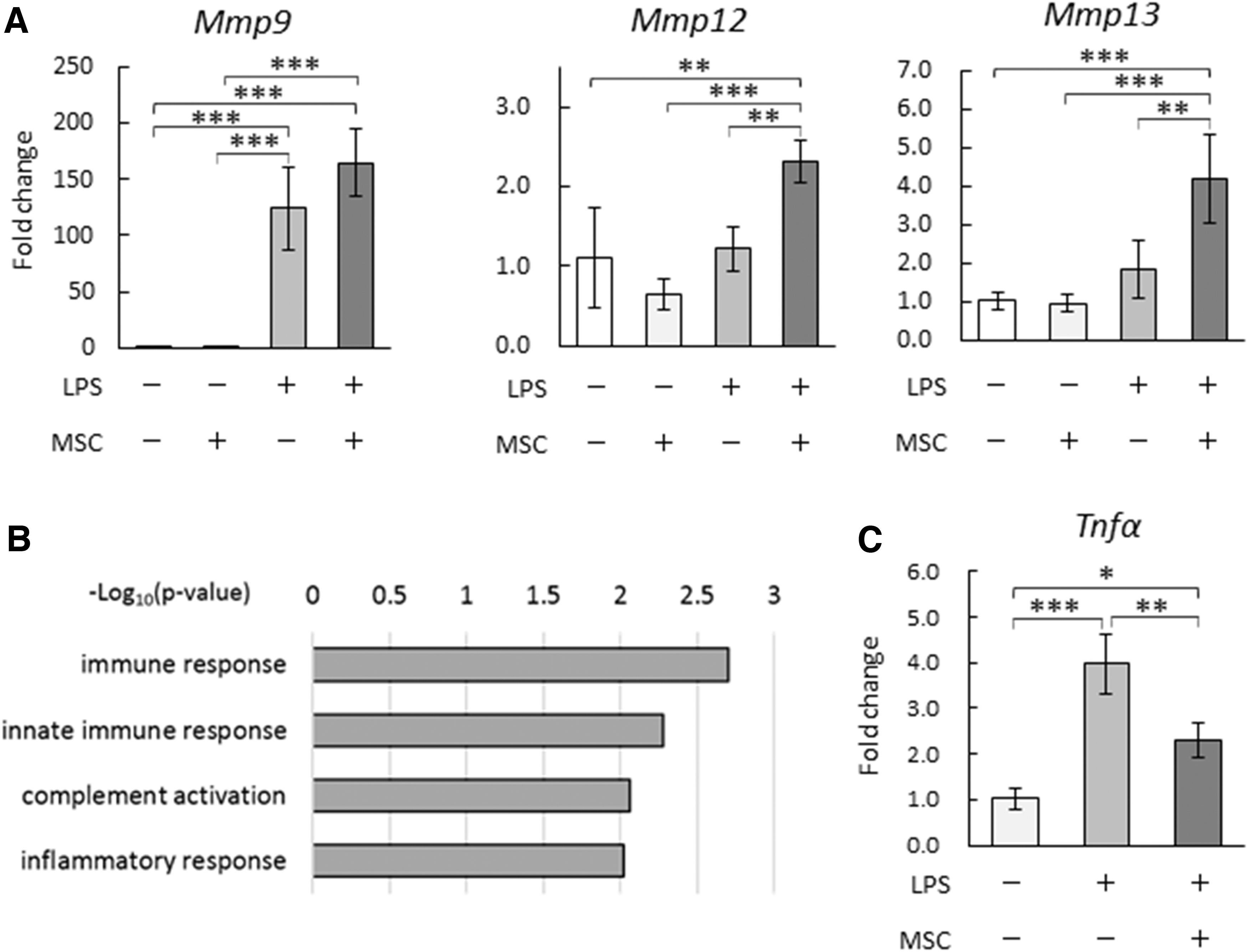
Co-culture with BMSCs increased Mmp expression and decreased Tnfα expression in LPS-stimulated BMDMs.
We also used a microarray to perform a comprehensive analysis of gene expression changes in LPS-stimulated BMDMs by co-culturing with BMSCs. KEGG pathway-enrichment analysis was performed with genes that showed a significant decrease in expression of ≥1.5-fold in the co-culture group, compared with the control group (Supplementary Table S1). Our analysis revealed enrichments of genes related to KEGG pathways, such as immune response, innate immune response, complement activation, and inflammatory response (Fig. 1B). Of note, among the inflammatory cytokines examined, Tnfα was significantly downregulated in LPS-stimulated BMDMs after co-culturing them with BMSCs (Fig. 1C).
miR-6769b-5p was highly expressed in EVs from the supernatants of co-cultured BMSCs and LPS-stimulated BMDMs
Humoral factors secreted by LPS-stimulated BMDMs and BMSCs were considered to be involved in the phenotypic changes of BMDMs by indirectly co-culturing with BMSCs. In this study, we examined the effects of EV-miRNAs on BMDMs co-cultured with BMSCs. MiRNAs were extracted from EVs in the co-culture supernatants of BMSCs and LPS-stimulated BMDMs, and array analysis was performed.
As reference EV-miRNAs present in sera from healthy mice, 23 miRNAs were extracted from the microarray data that showed a fold change in expression of ≥2 and a global normalization ratio of >100 in the co-culture supernatant. Among these miRNAs, seven miRNAs exhibited at least a 5-fold change in expression (miR-135a-1-3p, miR-8113, miR-6769b-5p, miR-7662-3p, miR-1247-3p, miR-6914-5p, and miR-8099); miR-6769b-5p exhibited the highest expression level (Table 2).
Upregulated MicroRNAs in Extracellular Vesicles from Co-Culture Supernatants of Lipopolysaccharide-Stimulated Bone Marrow-Derived Macrophages and Bone Marrow-Derived Mesenchymal Stem Cells
miR-6769b-5p increased MMP9 expression and suppressed inflammatory cytokine expression in LPS-stimulated BMDMs
To investigate the effects of miRNAs that were highly expressed in co-culture supernatants (from BMSCs and LPS-stimulated BMDMs) on Mφs, each miRNA mimic of the five miRNAs with >6-fold change in expression was transfected into LPS-stimulated BMDMs, followed by real-time PCR analysis to detect gene expression changes in MMPs and inflammatory cytokines. The transfection efficiency of BMDMs was confirmed using a fluorescent oligonucleotide and was found to be ≥80% (data not shown).
Of the five miRNAs with >6-fold changes in expression, miR-6769b-5p was the only miRNA that increased Mmp9 and decreased the expression of inflammatory cytokines such as Tnfα and Il-1β in LPS-stimulated BMDMs, compared with the control group (NC-miR-transfected group) (Fig. 2A). Gelatin zymography confirmed a significant increase in MMP9 secretion in the culture supernatant of miR-6769b-5p-transfected, LPS-stimulated BMDMs (Fig. 2B).
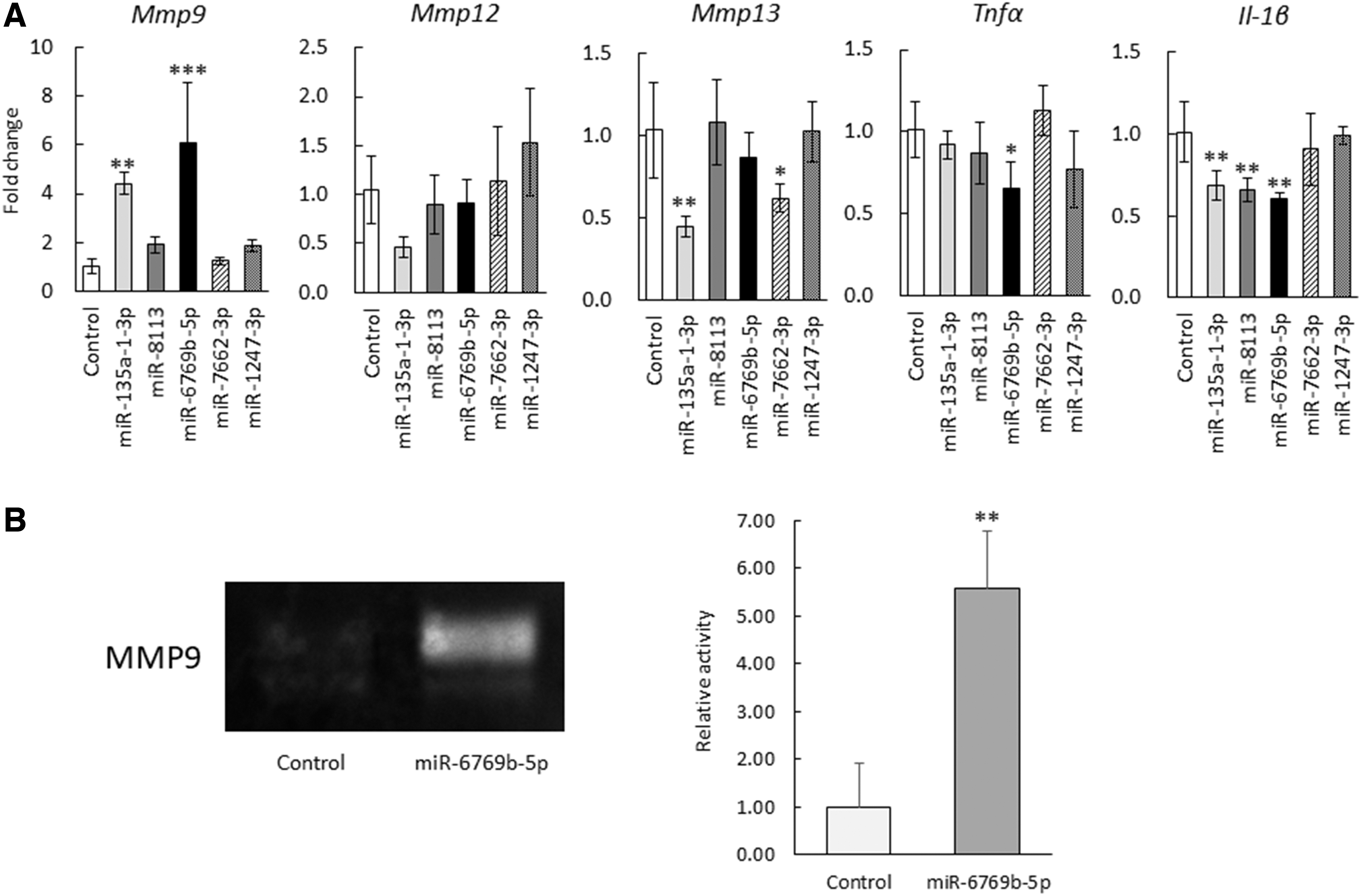
miR-6769b-5p increased MMP9 expression and reduced inflammatory cytokine expression in LPS-stimulated BMDMs.
BMSC-derived humoral factors increased miR-6769b-5p expression in LPS-stimulated BMDMs
Changes in miR-6769b-5p expression in BMDMs caused by BMSC-derived humoral factors were examined using LPS-stimulated BMDMs indirectly co-cultured with BMSCs and LPS-stimulated BMDMs cultured with BMSC-CM. In LPS-stimulated BMDMs, miR-6769b-5p was upregulated after co-culture with BMSCs (Fig. 3A). Comparison of unstimulated BMDMs, LPS-stimulated BMDMs, and LPS-stimulated BMDMs cultured with BMSC-CM revealed that LPS stimulation reduced miR-6769b-5p expression in BMDMs, whereas the addition of BMSC-CM increased miR-6769b-5p expression in LPS-stimulated BMDMs to levels comparable with those observed in unstimulated BMDMs (Fig. 3B). These results suggest that BMSCs secrete humoral factors that upregulate miR-6769b-5p in LPS-stimulated BMDMs.
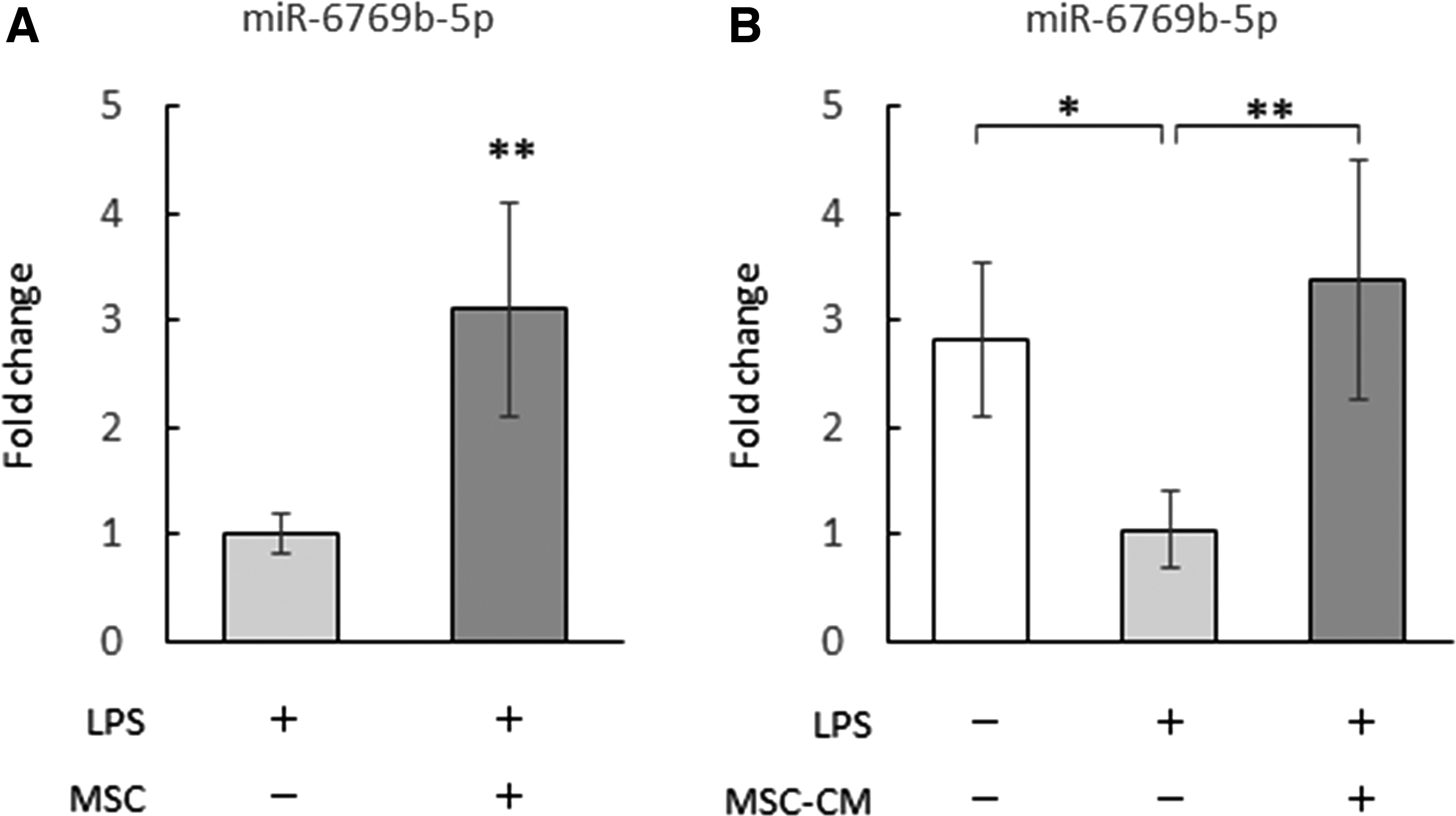
BMSC-derived humoral factors increased miR-6769b-5p expression in LPS-stimulated BMDMs.
miR-6769b-5p controlled the phenotype of LPS-stimulated BMDMs by downregulating ATF4
To elucidate the mechanism of action of miR-6769b-5p, we used a microarray to analyze gene expression levels in LPS-stimulated BMDMs that were transfected with miR-6769b-5p mimic or NC-miR. IPA analysis was conducted with a group of dysregulated genes (818 upregulated genes and 524 downregulated genes) that showed significantly different expression levels (≥1.5-fold) compared with the control group (NC-miR-transfected group). The IPA analysis revealed a significant change in the eukaryotic initiation factor (eIF) 2-signaling pathway, which suggested that decreased activating transcription factor 4 (Atf4) expression was to be expected (Fig. 4A, B). Genes related to eIF2 signaling that showed a > 1.5-fold change in expression (with P < 0.05) are given in Supplementary Table S2.

miR-6769b-5p regulated the phenotype of LPS-stimulated BMDMs by suppressing Atf4 expression.
No significant changes in eIF2α phosphorylation were observed (data not shown), but a significant decrease in Atf4 expression was observed (Fig. 4C). Furthermore, in LPS-stimulated BMDMs in which Atf4 was knocked down using siRNA, upregulation of Mmp9 and downregulation of both Tnfα and Il-1β were confirmed, relative to the corresponding expression levels in the control group (NC-siRNA transfection group) (Fig. 4D). Of interest, the same changes in gene expression observed in miR-6769b-5p-transfected, LPS-stimulated BMDMs were also observed in Atf4-silenced, LPS-stimulated BMDMs; hence, these findings suggest that miR-6769b-5p regulated the phenotype of LPS-stimulated BMDMs by suppressing ATF4 expression.
Discussion
In this study, we investigated phenotypic changes in BMDMs owing to their interaction with BMSCs and further analyzed EV-miRNAs as potential factors involved in the mechanisms underlying the phenotypic changes. We found that miR-6769b-5p was highly expressed in EVs in the co-culture system with BMSCs and LPS-stimulated BMDMs, and that miR-6769b-5p suppressed proinflammatory cytokine expression and upregulated MMP9 in LPS-stimulated BMDMs by downregulating ATF4. This is the first report describing the effects of miR-6769b-5p on BMDMs.
In liver cirrhosis and alcoholic liver injury, the LPS concentration in the portal blood increases owing to increased permeability of the intestinal mucosa [25 –27]. In NASH, increased sensitivity to LPS through Toll-like receptor (TLR)-4 causes increased intrahepatic TNF-α production by Mφs, resulting in inflammation [27]. Thus, LPS derived from the intestinal tract participates in the progression of liver disease, and it is important to control Mφ activation caused by LPS stimulation. Therefore, in this study, we studied LPS-stimulated BMDMs.
Liver fibrosis occurs when the balance between fibrolysis (caused by Mφs) and fibrogenesis (caused by hepatic stellate cells) is disrupted. During fibrosis, Mφs secrete inflammatory cytokines and profibrotic factors. In contrast, when fibrosis resolution occurs, Mφs change from a fibrogenic phenotype to a fibrolytic phenotype, produce high levels of MMP, and reduce inflammatory cytokine production [19 –22]. Therefore, it is important to induce this Mφ phenotypic switch to alleviate liver fibrosis. In this study, we found that co-culture with BMSCs and transfection of miR-6769b-5p upregulated MMPs and downregulated inflammatory cytokines in BMDMs. These results suggest that miR-6769b-5p at least partially contributes to the phenotypic switch to the fibrolytic phenotype.
Indirect co-culturing with MSCs has been reported to suppress the expression of inflammatory cytokines such as TNF-α, IL-1β, IL-6, and IL-12 in LPS-stimulated Mφs [28 –31]. EVs derived from adipose tissue-derived MSCs have also been found to change the phenotype of LPS-stimulated Mφs to alternately activate Mφs (M2-Mφs) [32]. In this study, we found that miR-6769b-5p expression in LPS-stimulated BMDMs was increased by BMSC-derived humoral factors, and inflammatory cytokine expression was suppressed by transfecting miR-6769b-5p mimic, suggesting the potential involvement of miR-6769b-5p in the anti-inflammatory effect of BMSCs on BMDMs.
Watanabe et al. reported that MMP13 expression was increased by co-culturing BMSCs and colony-stimulating factor-1-induced BMDMs [23]. Our results show that co-culturing LPS-stimulated BMDMs with BMSCs increased the expression of Mmp12, Mmp13, and Mmp9. These results suggest that MMPs may be more markedly expressed in Mφs in an inflammatory environment, which could be effective in treating chronic liver disease in which fibrosis follows inflammation. Moreover, along with its anti-inflammatory effect on Mφs, the increase in MMP9 expression caused by miR-6769b-5p suggests that it may be an effective treatment for liver fibrosis.
In this study, changes in eIF2 signaling were observed in miR-6769b-5p-transfected, LPS-stimulated BMDMs along with decreased Atf4 expression. ATF4 is a member of the ATF/CREB transcription factor family and plays important roles in various stress responses, such as endoplasmic reticulum stress. In the human THP-1 monocytic cell line, ATF4 translocates to the nucleus after LPS stimulation through the TLR4-myeloid differentiation primary response 88 (MyD88) pathway and increases the expression of inflammatory cytokines such as TNF-α, IL-6, and IL-8 [33].
In addition, in a study involving murine BMDMs and intraperitoneal Mφs, ATF4 was activated by saturated fatty acids and increased the expression of inflammatory cytokines such as IL-6 and TNF-α, and it was reported to elicit a synergistic effect with the TLR4-signaling pathway [34]. These findings showed that ATF4 downregulation resulted in suppressed production of inflammatory cytokines [33,34]. Both saturated fatty acids and LPS are involved in hepatic fat conversion and inflammation in NASH; therefore, regulation of ATF4 expression in Mφs (as demonstrated in this study) may prove to be a promising therapeutic approach for treating NASH. Furthermore, regulating ATF4 expression increased the expression of MMP9 in Mφs, suggesting that it may be potentially useful for treating NASH with fibrosis, although in vivo experiments are desirable.
MiR-6769b-5p has been detected in intestinal epithelium-derived exosomes, and it suppresses the expression of the target molecule, ataxin-1, in nerve cells, thereby activating nerve cells [35]. Therefore, exosomal miR-6769b-5p has been linked to cell–cell interactions. As Mφs regulate the activation of other immune cells by exosomes [36], it is possible that the EVs secreted by BMDMs, whose miR-6769b-5p expression was increased by BMSCs, can change the phenotype of surrounding Mφs.
In conclusion, we found that miR-6769b-5p, whose expression in BMDMs was increased by BMSCs, induced a phenotypic switch to fibrolytic Mφs, in which MMP9 expression was increased, and inflammatory cytokine production was suppressed owing to reduced ATF4 expression. Future studies are warranted to investigate the humoral factors involved in regulating Mφ miR-6769b-5p expression by MSCs. In addition, according to database predictions, Atf4 is not included in the list of target genes of miR-6769b-5p; hence, it is necessary to investigate the mechanism whereby miR-6769b-5p suppresses ATF4 expression. Further elucidation of the Mφ phenotypic-control mechanism by MSCs through such studies will yield useful findings for developing antifibrotic treatments.
Footnotes
Acknowledgments
The authors thank Mariko Yamada, Kumie Ota, and Risa Mochizuki for providing technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by JSPS KAKENHI grants to Isao Sakaida (grant no. JP17H04162), Taro Takami (grant no. JP17K09428), and Toshihiko Matsumoto (grant no. JP16K211932).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.