
Abstract
The endometrial cycle in response to hormonal stimulation is essential for implantation. The female has endometrium that repeats this cycle through about half of a lifetime. The cycle includes three phases, proliferative, secretory, and menstrual, and each phase has distinct characteristics. The endometrial stromal cells (EnSCs) in each phase also have specialized characteristics, including cell cycle, morphologies, and cellular metabolic state. So we hypothesized that the cells in each phase have unique mitochondrial morphologies because they are generally linked to cellular metabolic state. To investigate the metabolic characteristics in each phase, we investigated the mitochondrial morphologies by transmission electron microscopy, oxygen consumption rate (OCR), and intracellular adenosine triphosphate (ATP) production. The decidualized EnSCs have shorter mitochondria than those in the proliferative phase. Besides, they also displayed distinct intracellular structural characteristics compared with the proliferative phase, such as ribosome-rich endoplasmic reticulum and increased formation of vesicles. OCR and luminescent ATP detection assay revealed that the basal respiration and ATP production in the decidualized EnSCs were lower than those in the proliferative phase. Thus, we concluded that morphological and intracellular structural changes were induced during the decidualization. Moreover, the decreased mitochondrial length was shown to correlate with decreased dependency on oxidative phosphorylation and ATP concentration in EnSCs.
Introduction
In the female reproductive system, endometrial changes in response to hormonal stimulation are essential for implantation of the embryo and pregnancy. The human endometrium repeats the endometrial cycle that includes periodic proliferation, decidualization (secretory phase), and menstruation, in response to the ovarian steroid hormones at least 400 times in a woman's lifetime [1]. Decidualization of endometrial cells is an essential process for pregnancy, and endometrial stromal cells (EnSCs) are responsible for the initial step of decidualization through cellular and molecular transformation [2,3].
EnSCs are mesenchymal cells that underlie the luminal endometrial epithelium layer of the uterus [4]. EnSCs and epithelial cells closely interact with each other in response to ovarian steroid hormones. EnSCs mediate the cyclic proliferative response of epithelial cells [5]. In this process, EnSCs are transformed into secretory decidual cells through mesenchymal–epithelial transition, which is induced by the upregulation of progesterone and cyclic adenosine monophosphate (cAMP) [6].
It has been reported that the decidualization of EnSCs entails morphological changes in intracellular organelles such as the expansion of the endoplasmic reticulum and the Golgi apparatus. It also induces multiple biochemical changes such as accumulation of glycogen and lipids in the cytoplasm, increased expression of extracellular proteins, and proinflammatory and anti-inflammatory responses [7 –9]. It is suggested that these dynamic changes are essential for implantation and postimplantation support to embryos [9].
EnSCs can be isolated and cultured in vitro to mimic the functions of their in vivo counterparts. Herein, we used an immortalized human EnSC line (T-hEnSCs), which could mimic the proliferation and secretory phase of the endometrium in response to hormonal and cAMP treatment. Immortalized T-hEnSCs are karyologically stable and morphologically similar to their parental cells [10]. As EnSCs are morphologically dynamic, we hypothesized that the changes in EnSCs should entail changes in organelle morphology and energy metabolism.
Thus, we aimed to investigate the morphological changes in organelles, specifically mitochondria, during the proliferation and decidualization phases of EnSCs. Furthermore, we aimed to assess the metabolic changes and elucidate the relationship between morphological changes in mitochondria and energy metabolism by electron microscopy and measurement of the oxygen consumption rate (OCR).
Materials and Methods
T-hEnSC culture
T
Decidualization of T-hEnSCs
To induce decidualization, proliferative T-hEnSCs in the proliferation media were washed twice in phosphate-buffered saline (Thermo-fisher) and cultured in DMEM/F-12 medium supplemented with 2% CCD-FBS, 100 U/mL penicillin, 0.1 mg/mL streptomycin, 1 μM medroxyprogesterone acetate (MPA; Sigma-Aldrich), and 0.5 mM cAMP (Sigma-Aldrich). Control cells were used to confirm whether the decidualization protocol using cAMP and MPA was successful. Accordingly, control cells were cultured in DMEM/F-12 medium supplemented with 2% CCD-FBS, 100 U/mL penicillin, and 0.1 mg/mL streptomycin without MPA and cAMP. The media were changed every 72 h, and the cultures were maintained for 6 days [11].
Total RNA extraction and quantitative reverse-transcription polymerase chain reaction
Total RNA was isolated from control, proliferative, and decidualized T-hEnSCs using TRIzol reagent (Invitrogen, MA). One microgram of total RNA was reverse transcribed with SuperScript III Reverse Transcriptase Kit (Enzynomics, Daejeon, Republic of Korea) according to the manufacturer's instructions. For real-time polymerase chain reaction (PCR), glyceraldehyde-3-phosphate dehydrogenase (Gapdh) gene was used as reference control. Quantitative reverse-transcription-PCRs (qRT-PCRs) were set up in duplicate with the Power SYBR Green Master Mix (Enzynomics) and analyzed with the Roche LightCycler 5480 (Hoffmann-La Roche, Basel, Switzerland). PCR amplification was performed using gene-specific primers listed in Table 1. The mRNA expression of established biochemical markers of decidualization, prolactin (Prl), and insulin-like growth factor-binding protein-1 (Igfbp1), was measured in T-hEnSCs [12 –14].
List of Primers Used for Quantitative Reverse-Transcription Polymerase Chain Reaction
Gapdh, glyceraldehyde-3-phosphate dehydrogenase; Igfbp1, insulin-like growth factor-binding protein-1; Prl, prolactin.
Transmission electron microscopy
Transmission electron microscopy (TEM) was performed according to previous reports [15 –17]. For TEM observation, all samples were fixed in 2.5% glutaraldehyde (Sigma-Aldrich) in 0.1 M phosphate buffer overnight. After washing with 0.1 M phosphate buffer, the samples were fixed with 1% osmium tetroxide (Sigma-Aldrich) in 0.1 M phosphate buffer for 1 h. All samples were dehydrated in a series of graded ethanol. The samples were embedded in Epon 812 and then polymerized at 60°C for 3 days. Ultrathin sections (60–70 nm) were obtained using Leica Ultracut UCT ultramicrotome. The ultrathin sections placed on a grid (200 mesh) were examined with TEM (JEM 1010; JEOL, Tokyo, Japan) operating at 60 kV, and the images were recorded by a charge-coupled device camera (SC1000; Gatan, CA).
Mitochondrial length analysis
The mitochondria visualized in TEM images were analyzed using ImageJ 1.43 (National Institutes of Health, MD) to calculate the maximum/minimum (max/min) ratio of mitochondrial length. At least 50 mitochondria from random grid were measured and analyzed to obtain these data.
Measurement of OCR
For measuring the OCR, samples were analyzed using Seahorse XFe96 Analyzers (Seahorse Bioscience, MA) [18]. On day 6 of the culture, cells were seeded in Seahorse XFe96 Cell Culture Microplates (96-well) at a density of 2 × 106 per well and incubated at 37°C in a 5% CO2 incubator for 1 day. Before analysis, the medium was replaced with XF Assay Medium containing 5 mM sodium pyruvate (Thermo-fisher), 2.5 mM
Measurement of ATP level
To measure ATP levels, we used the Luminescent ATP Detection Assay Kit (Abcam, Cambridge, United Kingdom) with GloMax 96 Microplate Luminometer (Promega, WI). A total of 2 × 104 control, proliferative, and decidualized cells were cultured in 96-well cell culture plates (SPL life science, Gyeonggi-do, Republic of Korea) precoated with Matrigel 24 h before the assay. After treatment, the procedure was performed according to the manufacturer's instructions.
Statistical analysis
All experiments were performed in triplicate. qRT-PCR experiments were analyzed statistically using ΔCq values. Mean ± standard error of the mean was used to indicate the statistical results. The significance of differences was assessed by one-way analysis of variance (ANOVA) with Tukey's Honestly Significant Difference post hoc or Brown–Forsythe and Welch ANOVA with Tamhane's T2 post hoc for multiple comparisons. P values <0.05 were considered statistically significant. All of statistical analysis were performed using GraphPad Prism 8 according to the manufacturer's instructions.
Results
Morphological and molecular signature changes in T-hEnSCs during decidualization by treatment with cAMP and MPA
To recreate the secretory phase of the menstrual cycle, the decidualization of T-hEnSCs was induced by treatment with cAMP and synthetic progestin (MPA) for 3 days, followed by culturing in a medium free of cAMP and MPA. As the decidualized state only lasts 6 days, induction of decidualization of T-hEnSCs was also performed for 6 days [19,20]. MPA is a type of progestin, and cAMP is the downstream signaling molecule of progesterone, and both are crucial for EnSCs to be transformed into the decidualized state [21]. After cAMP and MPA treatment, the morphology of T-hEnSCs changed from thin and elongated fibroblast-like cells into relatively round epithelioid-like cells on days 3 and 6, comparable with the untreated control group (Fig. 1A). The proliferation phase of T-hEnSCs showed a similar morphology to the control group (Fig. 1A). Next, the decidualization of T-hEnSCs was confirmed by the expression level of Prl and Igfbp1. The qRT-PCR analysis revealed that deciduous markers, Prl and Igfbp1, were upregulated in the cAMP and MPA-treated group on day 3 and further upregulated on day 6 (Fig. 1B, C). The expression level of Prl in the cAMP and MPA-treated group was ∼500,000-fold (day 3) and 1,000,000-fold (day 6) higher than that in the control group (Fig. 1B), and the expression level of Igfbp1 was ∼400-fold (day 3) and 1,000,000-fold (day 6) higher than that in the control group (Fig. 1C). However, no significant differences in the expression level of Prl and Igfbp1 were observed in the proliferative and control groups (Fig. 1B, C). Collectively, these results indicate that day 6 T-hEnSCs are more representative of decidualized EnSCs.
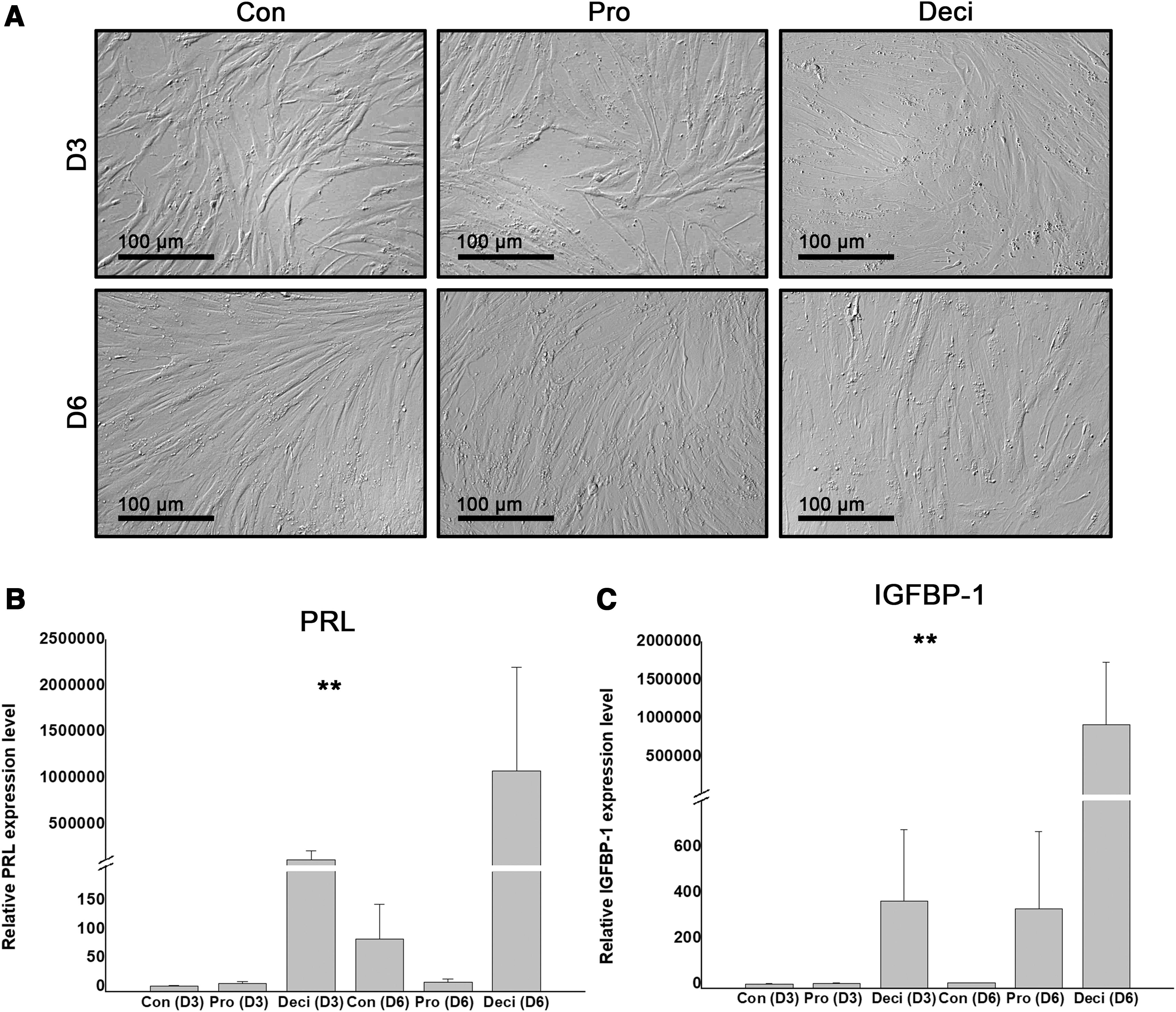
In vitro decidualization of immortalized hEnSCs (T-hEnSCs) by treatment with cAMP and MPA.
Ultrastructural analysis of decidualized T-hEnSCs
For further characterization of the decidualized T-hEnSCs, ultrastructural analysis using TEM was conducted in three groups of T-hEnSCs: control, proliferative, and decidualized cells (Fig. 2A–C). One of the most prominent features was the presence of phagocytotic vesicles. Decidualized T-hEnSCs showed many more phagocytotic vesicles in the cytoplasm than the other two groups of T-hEnSCs (Fig. 2A, B). Another prominent feature was the number of ribosomes. TEM analysis revealed more ribosomes in decidualized T-hEnSCs than control and proliferative T-hEnSCs (Fig. 2C). These results are consistent with previous reports that decidualized human EnSCs contain abundant ribosomes and vesicles [22,23].
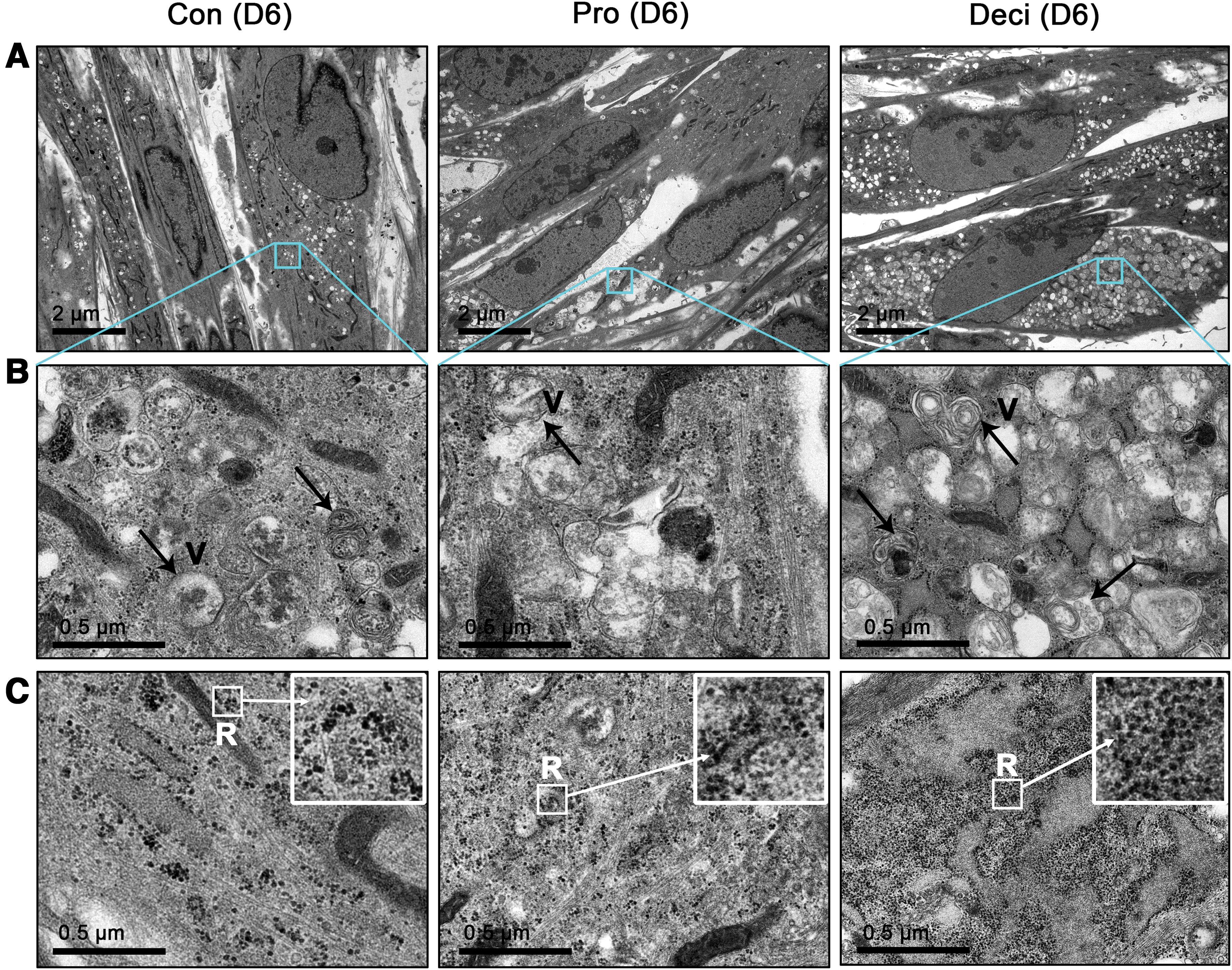
TEM images of Con, Pro, and Deci T-hEnSCs on day 6 after cAMP and MPA treatment.
Comparative analysis of mitochondrial morphology in proliferative and decidualized T-hEnSCs
To compare the mitochondrial morphology, T-hEnSCs at the proliferation and decidualization stages were compared with control cells using TEM analysis at a magnification of 25,000 × to examine the mitochondrial structure (Fig. 3). Similar to other differentiated somatic cells, all T-hEnSCs groups showed elongated mitochondria (Fig. 3A). However, we could detect the difference in length. Decidualized T-hEnSCs showed shorter mitochondrial length than control and proliferative groups. To quantify the morphological differences, we measured the length of maximum (max) and minimum (min) axes of mitochondria (Fig. 3B) and calculated the max/min ratio (Fig. 3C). Although there was no significant difference in the max values of mitochondria between control and proliferative T-hEnSCs, a significant difference was observed between control and decidualized T-hEnSCs (Fig. 3B). Also, there was no difference in min values in all groups, indicating that mitochondrial length could be compared without other variations (Fig. 3B). A comparison of the mean values of the mitochondrial max/min ratio also showed a significant difference between control and decidualized T-hEnSCs (Fig. 3C). The mean values of the max/min ratios in control, proliferative, and decidualized T-hEnSCs were 23.5, 18.0, and 12.7, respectively (Fig. 3C). These results indicate that decidualized T-hEnSCs contained a short length of mitochondria compared with control and proliferative T-hEnSCs, which prompted us to further investigate the difference of in energy metabolism in the three groups.
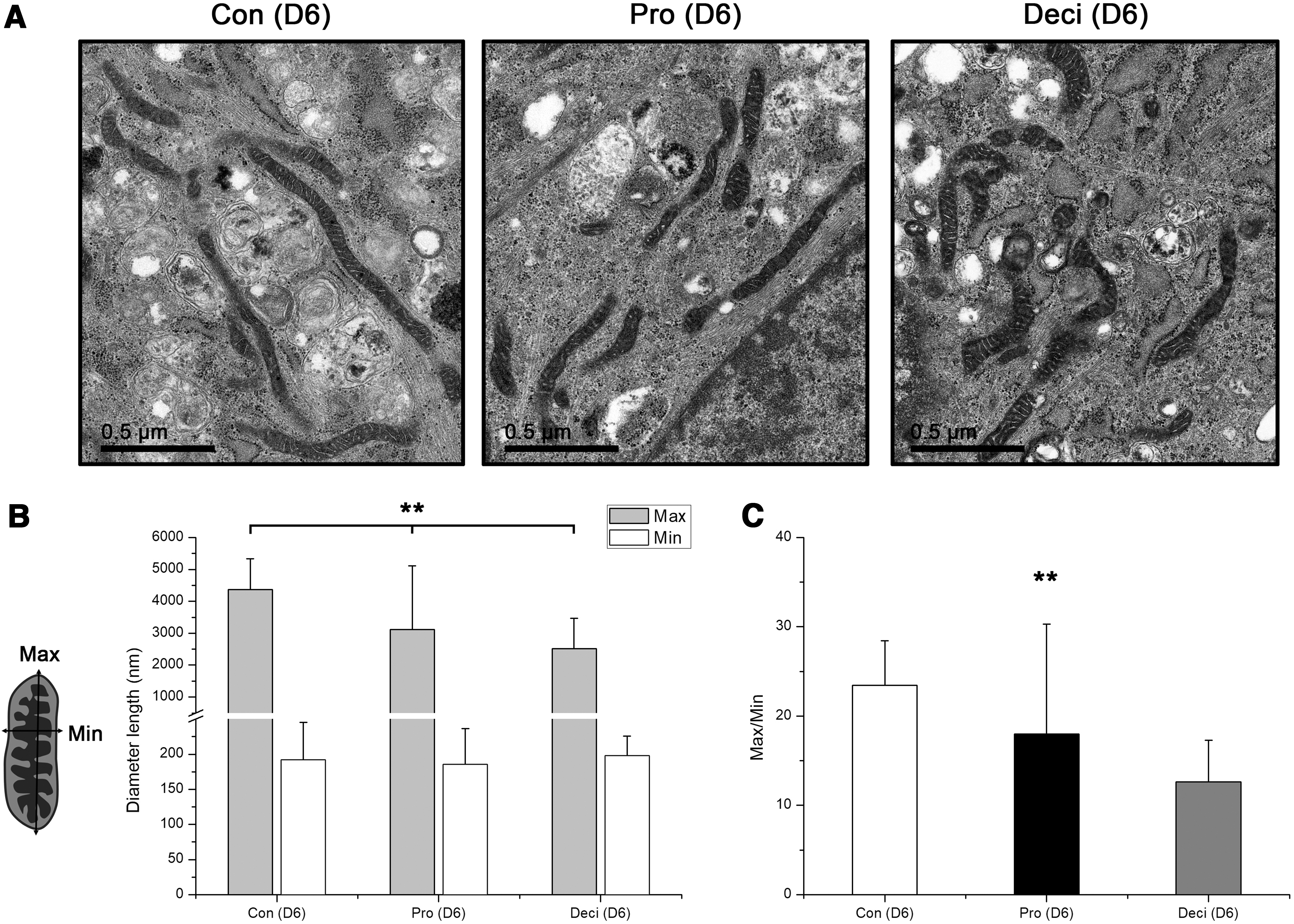
Comparison of mitochondrial morphology in T-hEnSCs during decidualization.
Mitochondrial OCR analysis according to developmental stages of T-hEnSCs
Mitochondria are energy-producing organelles whose shape changes dynamically depending on the cell type or the source of main energy production. Since the decidualized T-hEnSCs showed different mitochondrial morphology from control and proliferative T-hEnSCs, we hypothesized that there may be differences between decidualized T-hEnSCs and control or proliferative T-hEnSCs in terms of energy metabolism as well. To compare the energy metabolic differences, we measured OCR, which represents ATP production through mitochondrial OXPHOS (Fig. 4A). OCR is used to determine how much a certain cell depends on the OXPHOS system for its energy source, that is, the main parameters of mitochondrial function [24 –26]. Comparing the level of basal respiration, which represents the amount of oxygen consumption under normal conditions, significant differences were not observed among the three groups (Fig. 4B). After treatment with oligomycin, an ATP synthase inhibitor, ATP production levels were measured. Control and proliferative T-hEnSCs showed a higher level of ATP production than decidualized T-hEnSCs (Fig. 4C). Maximal respiration and spare respiratory capacity were measured after treatment with FCCP, which abolishes the proton gradient and mitochondrial membrane potential. Maximal respiration and spare respiratory capacity were much lower in decidualized T-hEnSCs than in control and proliferative T-hEnSCs (Fig. 4C, D). Collectively, XF Cell Energy Phenotype analysis showed that decidualized T-hEnSCs were less dependent on OXPHOS than control and proliferative T-hEnSCs for energy production (Fig. 4F). To confirm the ATP production levels of each T-hEnSC group, we further analyzed the total cellular ATP concentration using the Luminescent ATP Detection Assay Kit (Fig. 4G). Intracellular ATP concentration was the lowest in decidualized T-hEnSCs among all the T-hEnSCs. It was ∼1.6- and 1.7-fold higher in decidualized T-hEnSCs than in control and proliferative T-hEnSCs, respectively (Fig. 4G). These results demonstrate that decidualized T-hEnSCs exhibit an energy metabolism different from that in control and proliferative T-hEnSCs, which appears to be correlated with mitochondrial morphology.
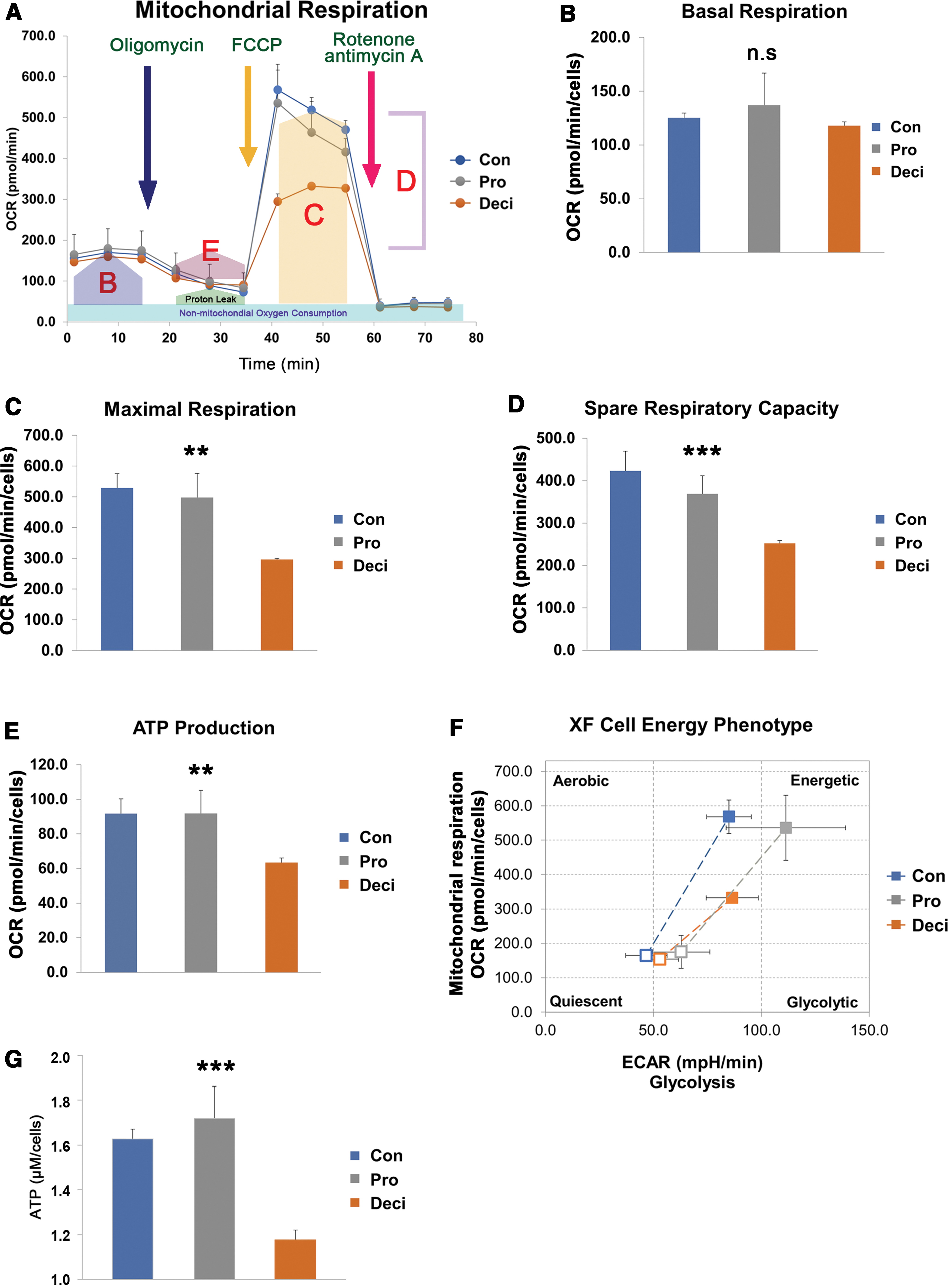
Mitochondrial OCR analysis according to developmental stage of T-hEnSCs.
Discussion
In this study, we examined the changes in the morphology of intracellular organelles and mitochondrial energy metabolism in T-hEnSCs using TEM and OCR analysis. Our results suggested that decidualization could be successfully simulated in T-hEnSC cells by treatment with MPA and cAMP. Decidualized T-hEnSCs displayed distinct rough endoplasmic reticulum, vesicles, mitochondrial morphology, and mitochondrial energy metabolism compared with proliferative T-hEnSCs. Since changes in EnSCs in the uterus are essential for implantation and embryonic development, it is predictable that intracellular organelles and energy metabolism will change to meet the cellular demand during the menstrual cycle.
To mimic the in vivo menstrual cycle, T-hEnSCs derived from myomas were treated with MPA and cAMP in an in vitro culture system. MPA and cAMP treatment successfully induced the decidualization of T-hEnSCs, as confirmed by ultrastructural analysis and deciduous markers, Prl and Igfbp1. Decidualized T
Mitochondria are dynamic organelles that play an important role in apoptosis, metabolism, reactive oxygen species (ROS) detoxification, and OXPHOS during proliferation and for cell survival [30]. Mitochondria readily transform and can be elongated or fragmented through fusion or fission, relatively efficiently [31 –34]. This transition allows mitochondria to regulate their function and adapt to their environment. Mitochondria elongate when exposed to oxidative stress. Otherwise, mitochondria become fragmented and ATP production by OXPHOS decreases, indicating that the shape and function of mitochondria are closely related [35,36].
Our results also showed that when T-hEnSCs transformed into the decidualized state, mitochondria became shorter, followed by a decrease in OCR and intracellular ATP concentration. The reasons underlying mitochondrial morphological changes during decidualization remain unclear. However, it is widely accepted that in response to cellular and environmental stresses, mitochondria undergo morphological transitions. In response to dynamic hormone levels, decidualization results in menstrual shedding and cyclic endometrial regeneration [37]. The underlying reason is that the cells are exposed to hypoxic conditions during decidualization and consequently upregulate the expression of vascular endothelial growth factor for angiogenesis [38]. This might be a potential reason why the decidualized T-hEnSCs exhibit lower oxygen consumption than those in their proliferative phase. Accordingly, clinical studies on how mitochondrial OCR or oxidative stress in EnSCs differs from the normal pregnancy group in patients with recurrent miscarriages or repeated implantation failures are required.
Recent reports also suggested that ectopic EnSCs from patients with endometriosis generated more ROS and ATP as well as superoxide dismutase [39]. However, decreased mitochondrial energy production was observed in the endometrium in a nonhuman primate with endometriosis, which might be caused by mitochondrial dysfunction [40]. Further studies are needed to determine the implication of the differences in species-specific metabolism.
Herein, we used an immortalized EnSC cell line, T-hEnSCs, instead of primary endometrium samples from women in different stages of the menstrual cycle. The in vitro EnSC line differs from primary EnSCs because cells in vivo may be affected by diverse hormones and unknown factors. In this study, however, we demonstrated that decidualization could be successfully simulated in the T-hEnSC line by treatment with MPA and cAMP, indicating that in vitro decidualization experiments using established cell lines could represent EnSCs from women in various stages of the menstrual cycle. However, further study is needed to elucidate whether the results obtained in vitro could be applicable to the decidualized EnSCs in vivo.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Research Foundation of Korea (NRF) grant funded by the government (MSIT) of the Republic of Korea (grant nos. 2016M3A9B6946835, 2015R1A5A1009701, and 2020R1A2C3007562).