
Abstract
This study was designed to assess whether the combination of the glucagon-like peptide-1 (GLP-1) analog exendin-4 (Ex4) and bone marrow-derived mesenchymal stem cell (BM-MSC) could enhance β-cell action in streptozotocin (STZ)-induced diabetic rats. Forty male Sprague–Dawley rats were randomly assigned to five groups: the normal control group (Normal), diabetes mellitus (DM) group, MSC-treated group (MSC), Ex4-treated group (Ex4), and MSC plus Ex4-treated group (MSC+Ex4). Body weight, blood glucose level, intraperitoneal glucose tolerance test, and in vitro glucose-stimulated insulin secretion were used to assess the treatment efficacy. The expression level of insulin, glucagon, pancreatic duodenal homeobox-1 (PDX-1), v-maf musculoaponeurotic fibrosarcoma oncogene homolog A (MafA), glucagon-like peptide-1 receptor (GLP-1R), and forkhead transcription factor 1 (FoxO1) was estimated by immunofluorescence analysis. Proliferation was assessed by Ki67 staining, and apoptosis was determined by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining in β-cells. Glucose-induced insulin secretion in the MSC+Ex4 group was significantly increased compared to that in the MSC group in vitro and in vivo. Compared to that of the other groups, the number of insulin-immunopositive cells was increased in both the MSC and MSC+Ex4 groups. However, β-cell proliferation and apoptosis in the MSC group and MSC+Ex4 group were not significantly different. Importantly, the expression level of PDX-1, MafA, FoxO1, and GLP-1R in β-cells in the MSC+Ex4 group was significantly higher than those in the MSC group. The numbers of insulin+ glucagon+ double positive cells and glucagon+ GLP-1+ double positive cells were significantly increased after MSC treatment and MSC+Ex4 combined treatment, suggesting the enhanced function of newly formed islet β-cells. Our findings showed that the combination of MSC and Ex4 enhanced the function of newly formed β-cells in STZ-induced diabetic rats by acting on multiple insulin transcription factors. Thus, combined MSC and Ex4 therapy provides a feasible approach for future diabetes treatments.
Introduction
Diabetes has become a challenging health care burden, with ∼366 million patients globally, and is predicted to reach 522 million by 2030 [1]. In type 1 diabetes (T1D), the autoimmune-mediated destruction of pancreatic β-cells triggers insulin deficiency. In type 2 diabetes (T2D), aberrantly generated insulin and peripheral insulin resistance are the main pathogeneses [2]. Current regimens involving both antidiabetic drugs and subcutaneous insulin injection cannot provide satisfactory blood glucose control in the same manner as functional pancreatic β-cells nor do these treatments prevent the progression or complications of diabetes. A therapeutic approach that replenishes β-cell mass in diabetic patients would not only restore normal glycemic control in the long term but also be beneficial to all diabetic patients regardless of the different causes of T1D and T2D.
Mesenchymal stem cells (MSCs) are characterized by their potent ability to differentiate into specific lineages, modulate the local microenvironment, secrete diverse factors, and activate endogenous progenitor cells. Therefore, MSCs are considered to be potential tools in the regeneration of damaged tissues and as treatments for multiple diseases [3].
The effect of MSCs on alleviating diabetes has been widely reported in numerous animal models [4 –10]. However, the exact contribution of MSC-mediated regeneration to the above-described treatment efficacy remains unclear. The fact that only 1.7%–3% of MSCs are capable of differentiating into β-cells with sufficient function in vivo [11] indicates that other potential mechanisms exist by which MSCs treat diabetes. In contrast, the immunomodulatory potential, anti-inflammatory, and antiapoptotic effects of MSCs contribute to the regenerative ability of MSCs [12 –15]. Therefore, MSCs are likely to modify the microenvironment by suppressing parenchymal cell apoptosis, as well as promoting the proliferation and differentiation of local progenitors [14,15].
MSC infusion can induce endogenous islet regeneration, yet the newly generated β-cells show improper maturation patterns and function. Our previous study showed that intravenously administered MSCs ameliorated hyperglycemia and restored pancreatic islets in streptozotocin (STZ)-induced diabetic rats, which were characterized by numerous newly formed pancreatic β-cells [4,16]. However, the newly formed β-cells did not appear to be fully functional, as random blood glucose level was not completely normalized. Approaches aimed at promoting the regeneration and maturation of β-cells demand further investigation.
It is well known that several potential β-cell growth factors have been identified, and glucagon-like peptide-1 (GLP-1), which is released by intestinal L-cells, is among the most valuable [17]. Exendin-4 (Ex4), a long-lasting GLP-1 receptor (GLP-1R) agonist, combined with GLP-1R could enhance β-cell mass in chemically or surgically induced diabetes models by suppressing β-cell apoptosis and stimulating islet neogenesis and β-cell proliferation [18 –21]. Moreover, GLP-1 is intensively involved in endocrine cell differentiation during pancreas development, possibly by inducing the expression of the pancreatic and duodenal homeobox gene [also known as pancreatic duodenal homeobox-1 (PDX-1)], which is necessary for β-cell-specific gene expression, as well as pancreas development [22,23].
In addition, previous studies have revealed that GLP-1 and Ex4 participate in the differentiation of embryonic stem cells and ductal progenitor cells into islet endocrine cells [18,24,25]. Thus, GLP-1 might critically participate in embryonic development of the pancreas and β-cell neogenesis in the islets of the adult pancreas [26]. In addition, it is hypothesized that GLP-1 could promote the maturation of immature β-cells by enhancing growth and survival [27]. Therefore, we wondered whether the combination of MSCs and Ex4 could stimulate β-cell regeneration in a diabetic model and whether islet function could be fully restored.
In the present study, we showed that combined treatment with MSCs and Ex4 could promote the regeneration and maturation of β-cells with fully restored islet function and long-term restoration of normal glycemia in an STZ-induced diabetic rat model.
Materials and Methods
Isolation, maintenance, and identification of bone marrow-derived MSCs
The isolation, purification, and identification of bone marrow-derived MSCs (BM-MSCs) were performed according to previously-described protocols [28,29]. The isolation, differentiation capacity, and phenotype of BM-MSCs can be found in our previous research and articles [4].
Animals and treatment
The animal procedures were approved by the Institutional Animal Care and Use Committee of the Chinese PLA General Hospital and were conducted in accordance with the guidelines of the China Council on Animal Care and Use. The animal-related experiments were conducted under the guidelines and approved by the Ethic Committee of the Chinese PLA General Hospital. The study included 40 male Sprague–Dawley (SD) rats of paired age and weight (8 weeks & 240–260 g). The animals were allowed free access to water and food and were maintained in a conditioned atmosphere at 25°C and 50%–55% humidity with a 12-h light/12-h dark cycle.
After 1 week of stabilization, eight rats were assigned to the normal control group (Group 1, Normal) and intraperitoneally injected with 0.1 mL of citrate buffer. The other animals were utilized to establish an STZ-induced diabetes model. Briefly, STZ powder (Sigma, St. Louis, MO, USA) was dissolved in sodium citrate buffer (pH 4.5). Then, a single administration of STZ was intraperitoneally injected at a dose of 65 mg/kg. Forty-eight hours after STZ administration, animals with random blood glucose level >25 mmol/L were included in further studies. During the whole study, the random blood glucose was measured at 7 am. After having fasted for 8 h, the fasting blood glucose was measured at 3 pm.
The diabetes mellitus (DM) model animals were randomly divided into four groups (n = 8) as follows:
Group 2 (DM): The diabetic rats were maintained with usual care without additional treatment.
Group 3 (MSC): A single dose of MSCs (2 × 106 cells/mL) was intravenously injected into the diabetic rats, followed by usual care for 4 weeks.
Group 4 (Ex4): Ex4 (Sigma) was intraperitoneally injected into the diabetic rats every day at a dose of 10 mg/kg for 4 weeks.
Group 5 (MSC+Ex4): After a single intravenous administration of MSCs (2 × 106 cells/mL), the diabetic rats were intraperitoneally injected with Ex4 daily at a dose of 10 mg/kg for 4 weeks [4].
The experimental flowchart is shown in Fig. 1.

Design and flowchart of the experiment.
BM-MSC application
MSCs at passages three to five were used. Briefly, diabetic rats of group 3 and group 5 were injected with MSCs [2 × 106 MSCs in 0.2 mL of normal saline (NS)] through the tail vein at the fifth day after STZ administration [4]. Untreated other diabetic animals were injected with 0.2 mL of NS.
Intraperitoneal glucose tolerance tests (IPGTTs) two weeks after MSC infusion and/or Ex4 treatment, IPGTT, and in vitro glucose-stimulated insulin secretion (GSIS) were conducted in rats in the different groups with a 1-day washout in Ex4 administered and concurrent nonadministered control rats. In brief, GTTs were conducted using an intraperitoneal injection of 2 g/kg D-glucose after the rats were fasted for 12 h, followed by the measurement of glucose level at 0, 30, 60, 90, and 120 min and serum insulin level at 0, 5, 10, 30, and 60 min. Plasma glucose concentrations were measured using an ACCU-CHEK Advantage Meter glucometer (Roche Diagnostics GmbH, Mannheim, Germany). ELISA (Rat Insulin ELISA Kit; Millipore, St. Charles, MO) was used to determine serum insulin level.
Islet isolation and glucose-stimulated insulin secretion (GSIS) in vitro
Rats were fasted for 12 h, and their pancreatic islets were isolated using the collagenase digestion method [30,31].
Briefly, individual rats were anesthetized, and their common bile duct was clamped near the major duodenal papilla and the end of the common hepatic duct. Subsequently, ∼10 mL of cold Hanks' solution containing 0.5 mg/mL of collagenase P (Roche, Basel, Switzerland) was injected using a 27-gauge hypodermic needle near the hepatopancreatic ampulla, which allowed the Hanks' solution to perfuse the entire pancreas through the pancreatic duct. The distended pancreas was then gently dissected out and digested with collagenase at 4°C for 10 min with gentle shaking. Digestion was stopped using fetal bovine serum solution (FBS, 10 mL; Gibco, Grand Island, NY) and Hank's solution (30 mL), and the tissue products were passed through a 400-μm filter and centrifuged on a Histopaque 1077 (Sigma).
The resolved islets at the interface were collected carefully and washed with Hank's solution. Individual islets were then handpicked using micropipettes under a dissection microscope. The isolated islets were cultured in Roswell Park Memorial Institute (Sigma, St. Louis, MO, USA) (RPMI) 1640 medium with 10% FBS overnight, and the aliquots (50/well) of islets were incubated in triplicate in RPMI 1640 medium containing 2.8 mM glucose at 37°C for 30 min. The islets were then incubated in RPMI 1640 medium containing 2.8 mM glucose, 16.7 mM glucose, or 200 μM palmitic acid at 37°C for 1 h.
Supernatants were harvested for the measurement of immunoreactive insulin (IRI) using an enzyme immunoassay (Linco Research, Inc., St. Charles, MO). The values of released insulin were normalized to islet protein contents and expressed as mU IRI/ng protein/h. The contents of the islet proteins were measured using a BCA Kit (Thermo Scientific, Rockford, IL) according to the manufacturer's instructions. GSIS was measured by the ratio of the mean amount of insulin in each group to the mean amount of insulin released in response to the low concentration of glucose (2.8 mM) in the normal control group (the baseline).
Preparation of tissue samples
Animals were sacrificed at the indicated times through intraperitoneal injection of 1% pentobarbital sodium (50 mg/kg) and followed by perfusion throughout the left ventricle with 100 mL phosphate-buffered saline and 500 mL 4% paraformaldehyde. The pancreas was excised and subsequently incubated in 30% sucrose/phosphate buffer overnight and frozen at −80°C for further study.
Immunofluorescence analysis
Frozen slides were incubated for 14 h at 4°C with specific antibodies against insulin (1/150, guinea pig; Sigma), v-maf musculoaponeurotic fibrosarcoma oncogene homolog A (MafA) (1/200, rabbit, Bethyl Laboratories), GLP-1R (1/100, rabbit; Sigma), glucagon (1/2,000, mouse; Sigma), PDX-1 (1/50, goat; R&D System), forkhead transcription factor 1 (FoxO1) (1/100, rabbit; Cell Signaling), and Ki67 (1/50, mouse; BD), followed by an additional 2 h of incubation with secondary antibodies (1:500; Alexa-594, Alexa-405, or Alexa-488; Invitrogen, Basel, Switzerland) at room temperature. An In Situ Cell Death Detection Kit (Roche) was used to assess β-cell apoptosis, followed by the visualization of nuclei by 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) staining (Sigma). A confocal FluoView FV1000 microscope (Olympus, Tokyo, Japan) was used for photography.
Morphological analysis and quantification
Fifty sequential 7-μm sections were obtained from each pancreatic sample. After IF staining, the number of different cell types in each islet was determined under a microscope by randomly counting 40 islets from 5 rats. The area was measured and analyzed with densitometry using ImageJ software (NIH, Bethesda, MD).
Statistical analysis
SPSS v.14.0.1 software was used for statistical analysis. All data were presented as the mean ± standard error of the mean. Differences between normal and diabetic vehicle groups were assessed for significance using Student's t-test (one-side). Statistical comparisons between the diabetic vehicle group and the treatment groups were performed using one-way analysis of variance (ANOVA), followed by Dunnett post hoc test and Tukey's post hoc test. A value of P < 0.05 was considered statistically significant.
Results
Combined MSC and Ex4 therapy ameliorated hyperglycemia in diabetic rats
To elucidate the role of MSC and Ex4 in STZ-induced diabetic rats, body weight, fasting blood glucose, and random blood glucose were measured.
Significantly reduced random blood glucose was detected in the MSC and MSC+Ex4 groups (P < 0.01) compared to the DM and Ex4 groups (Fig. 2A). In addition, there was a more obvious reduction in the mean blood glucose level in the MSC+Ex4 group than in the MSC group. Compared to that of the DM and Ex4 groups, fasting blood glucose was significantly lower (P < 0.01) in the MSC and MSC+Ex4 groups and was similar to that of the normal group (Fig. 2B). Fasting blood glucose was maintained at normal level during the study period.

Combined MSCs and Ex4 therapy lowers glucose and improves glucose intolerance in diabetic rats.
Moreover, IPGTT was carried out to evaluate the function of β-cells in the different groups 2 weeks after MSC treatment. As shown in Fig. 2D and E, rats in the MSC and MSC+Ex4 groups exhibited improved glucose intolerance and reduced area under curves (AUCs) of glucose compared to those of the other groups. Moreover, MSC+Ex4-treated rats had lower glycemic level during the IPGTT than MSC-treated rats.
Regarding body weight, treatment with MSCs alone or in combination with Ex4 induced obvious increases in body weight compared with those of untreated diabetic rats (Fig. 2C). Moreover, the treatment of diabetic rats with Ex4 only led to body weights similar to those of diabetic control rats.
MSC and Ex4 cotreatment increased the glucose-stimulated insulin release in vitro and in vivo
To clarify the effects of MSC and Ex4 on islet beta function, we analyzed pancreatic islet function in vitro and in vivo.
The IPGTT showed significantly decreased and delayed increases in blood insulin level in diabetic control rats compared with normal rats. The basal insulin secretion level in the MSC+Ex4 and MSC groups was higher than those in the other diabetic groups, but glucose-stimulated insulin secretion in the MSC group was not significantly different from basal insulin secretion level. However, compared to that of the MSC group and other groups, GSIS in the MSC+Ex4 group was significantly increased. Moreover, the highest level of insulin release stimulated by glucose in MSC+Ex4 treated diabetic rats was lower than that in normal animals (Fig. 3A).
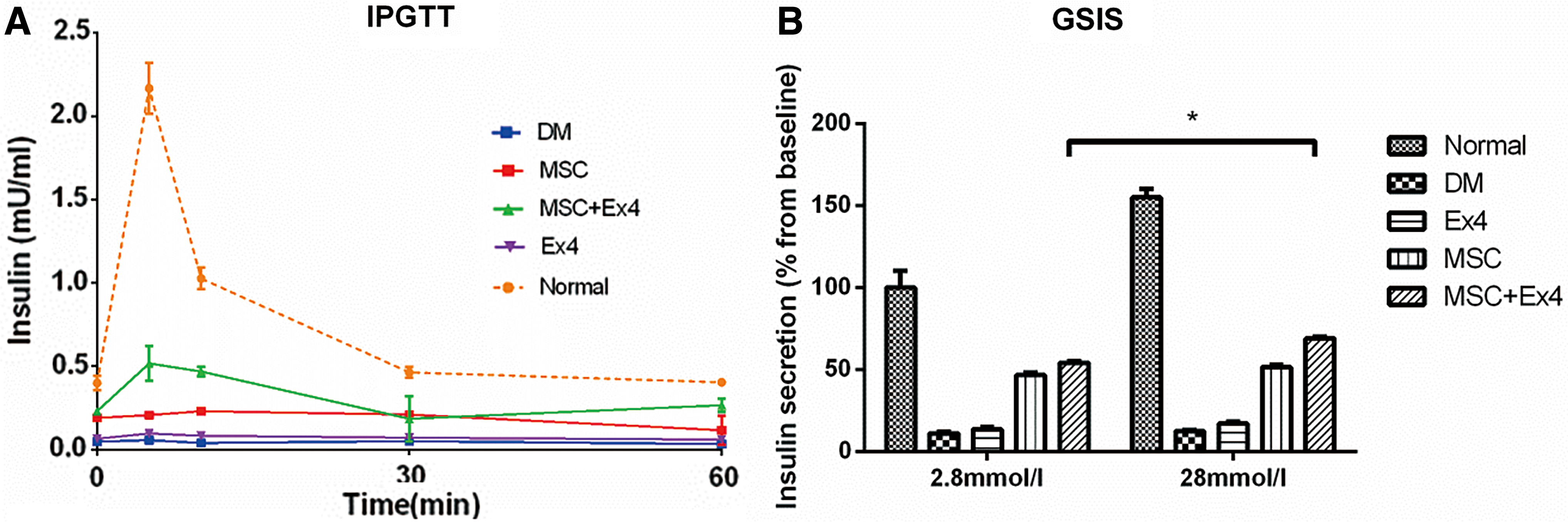
MSCs and Ex4 combined therapy can promote the secretion of insulin from newly formed islet β-cells both in vitro and in vivo.
To exclude the influence of external factors on blood glucose outside of the pancreas, repeated stimulation of the islets was performed by alternating administration of 2.8 mM glucose for 1 h and 16.7 mM glucose for another hour in the five groups, followed by the analysis of insulin level every hour. As expected, islets from normal rats exhibited robust and desired insulin secretion, whereas islets from Ex4 and DM-treated rats failed to secrete adequate insulin when challenged with glucose (Fig. 3B). The islets from the MSC and MSC+Ex4 groups showed a moderate pattern of insulin secretion. Furthermore, significantly elevated insulin levels were detected in the MSC+Ex4 group compared with the MSC group, indicating that the combined treatment of MSC and Ex4 induced effective insulin secretion under glucose stimulation.
These findings suggest that MSC treatment alone can lead to hyperglycemia due to delayed insulin secretion and improperly reduced level of circulating insulin in response to glucose challenge compared with those of the combined treatment group.
MSC and Ex4 increased the number of insulin positive cells
To determine the effect of MSCs and Ex4 on the numbers of α and β-cell in islets, we performed immunofluorescence staining for insulin and glucagon in samples from all five groups. As shown in Fig. 4, the ratio of the number of insulin positive cells on total islet cells was significantly decreased, and the ratio of the number of glucagon positive cells on total islet cells was significantly increased in untreated diabetic rats compared with normal control rats. Both MSC and MSC+Ex4 administration led to significant increases in the proportion of insulin immunopositive cell counts on total islet cells and diminished the proportion of glucagon immunopositive cell counts on total islet cells compared to those of the untreated diabetic group and Ex4 group. However, the percentage of the number of insulin positive cells on total islet cells was insignificantly different between the MSC group and the MSC+Ex4 group.
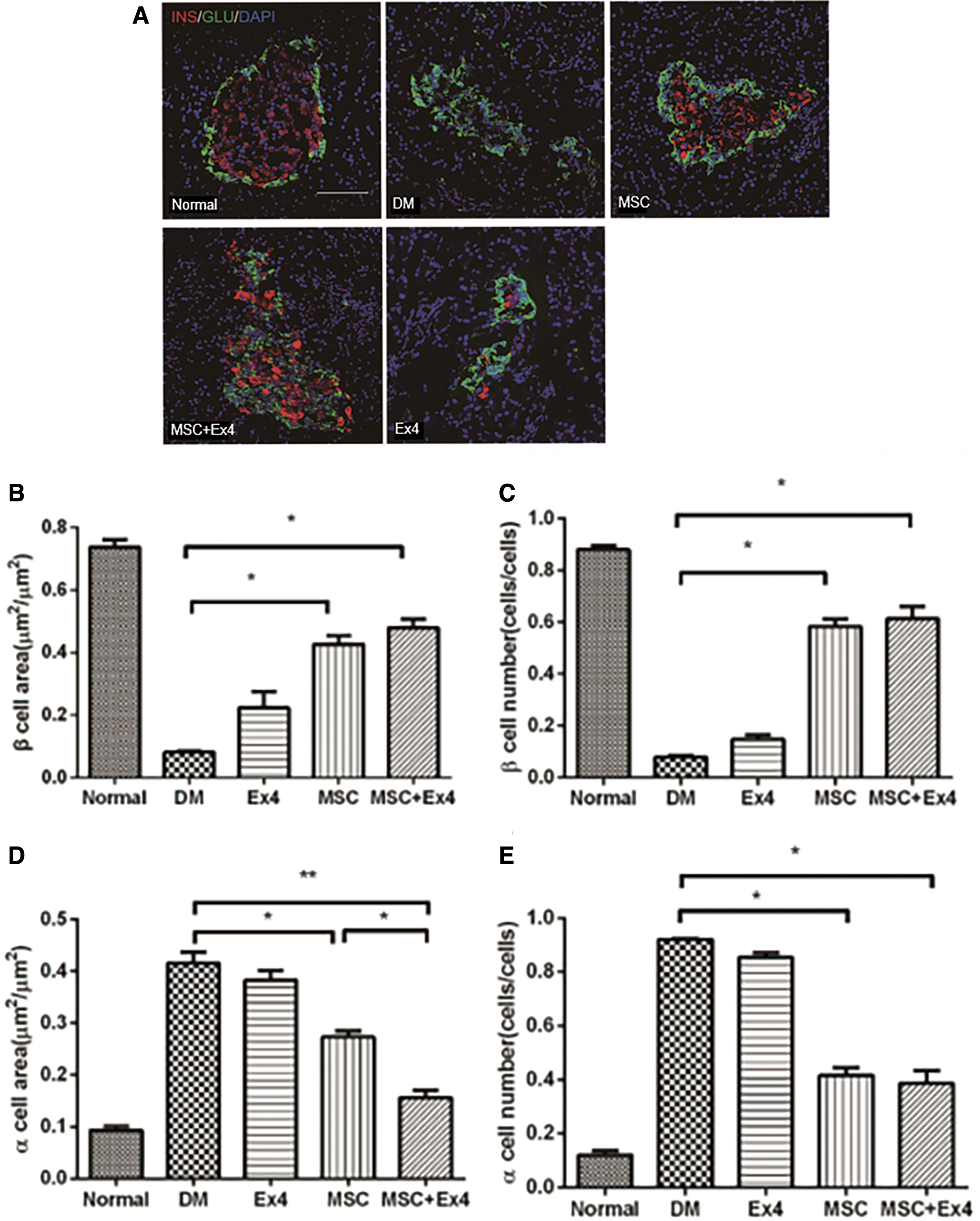
Effects of MSCs and Ex4 on the number of insulin+ and glucagon+ immunopositive cells.
MSC and Ex4 promoted the proliferation of newly formed β-cells
Next, we examined whether MSCs and Ex4 enhanced β-cell regeneration by stimulating β-cell proliferation and/or suppressing β-cell apoptosis in vivo.
Insulin-positive cells were stained for Ki67, which is expressed during replication and indicates mitotic cells. As shown in Fig. 5A, significant enhancement in β-cell proliferation was observed in rats treated with MSCs alone and in combination with Ex4 compared to rats in the diabetic control group and Ex4 group. However, the difference between the MSC group and the MSC+Ex4 group was not significant. Figure 5B shows an image of immunofluorescence staining, and the nuclei of proliferative cells of normal control group are green color. As shown in Fig. 5C, there were no differences in the proportion of insulin+ cells expressing terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) positive among the five groups, indicating that the combination of MSCs and Ex4 had no effect on the apoptosis of newly formed β-cells. Figure 5D shows an image of immunofluorescence staining, and the nuclei of apoptotic cells of MSC+Ex4 group are pink color.
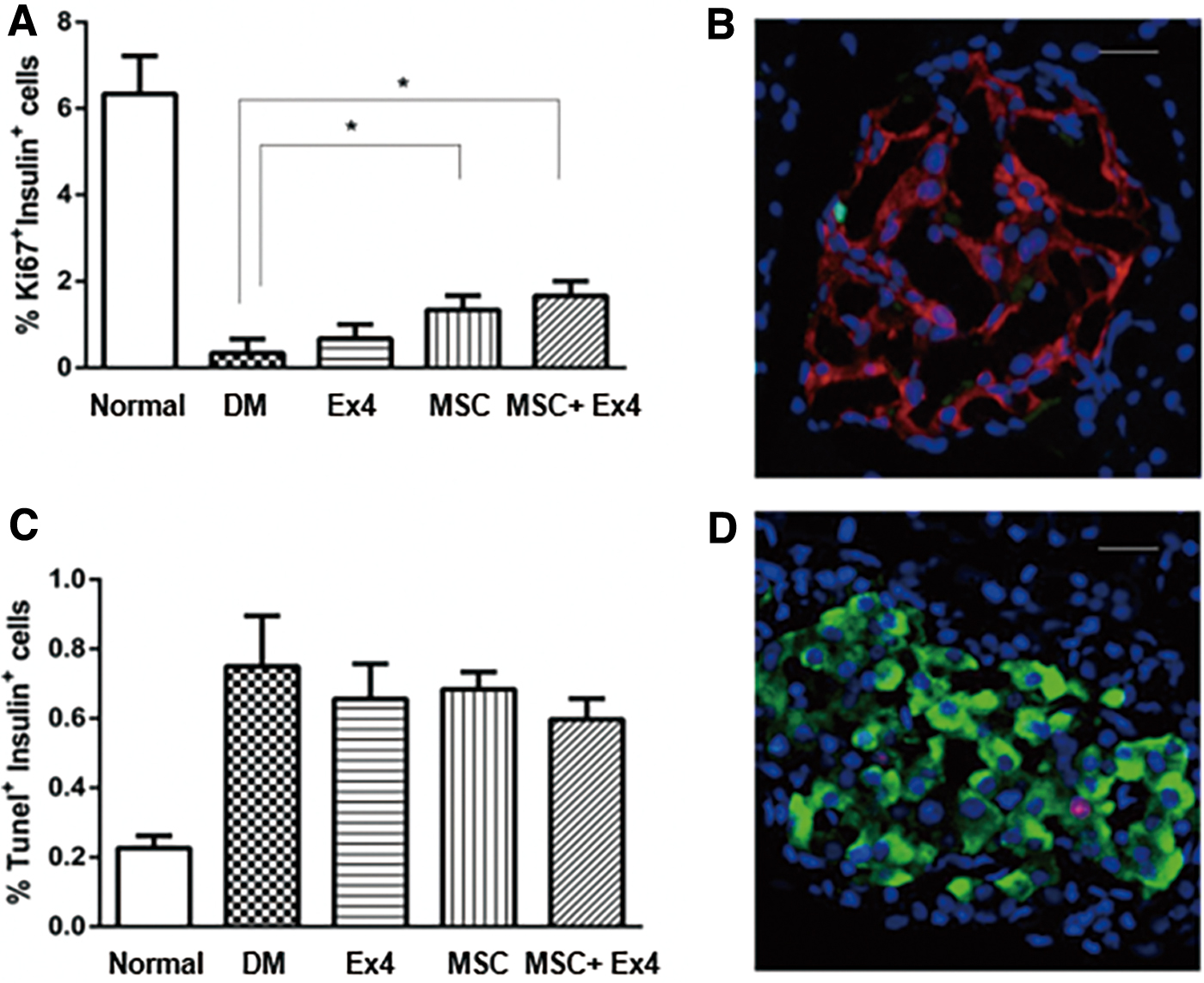
Effects of MSCs and Ex4 on proliferation and apoptosis of β-cells.
Taken together, these results indicated that the proliferation of newly formed β-cells was remarkably promoted by treatment with MSCs alone or together with Ex4, which contributed to the regeneration of newly formed β-cells.
The expression of MafA, FoxO1, and GLP-1R was significantly enhanced by MSC and Ex4
PDX-1 exerts a crucial effect on the formation and progression of endocrine and exocrine pancreases and is a determinant of β-cell identity. As shown in Fig. 6A and B, PDX-1 protein expression was significantly increased in rats administered MSCs alone or together with Ex4 compared with that of the other groups. The β-cell expression level of PDX-1 in rats that were cotreated with MSCs+Ex4 was significantly increased compared with those of rats treated with MSCs alone.
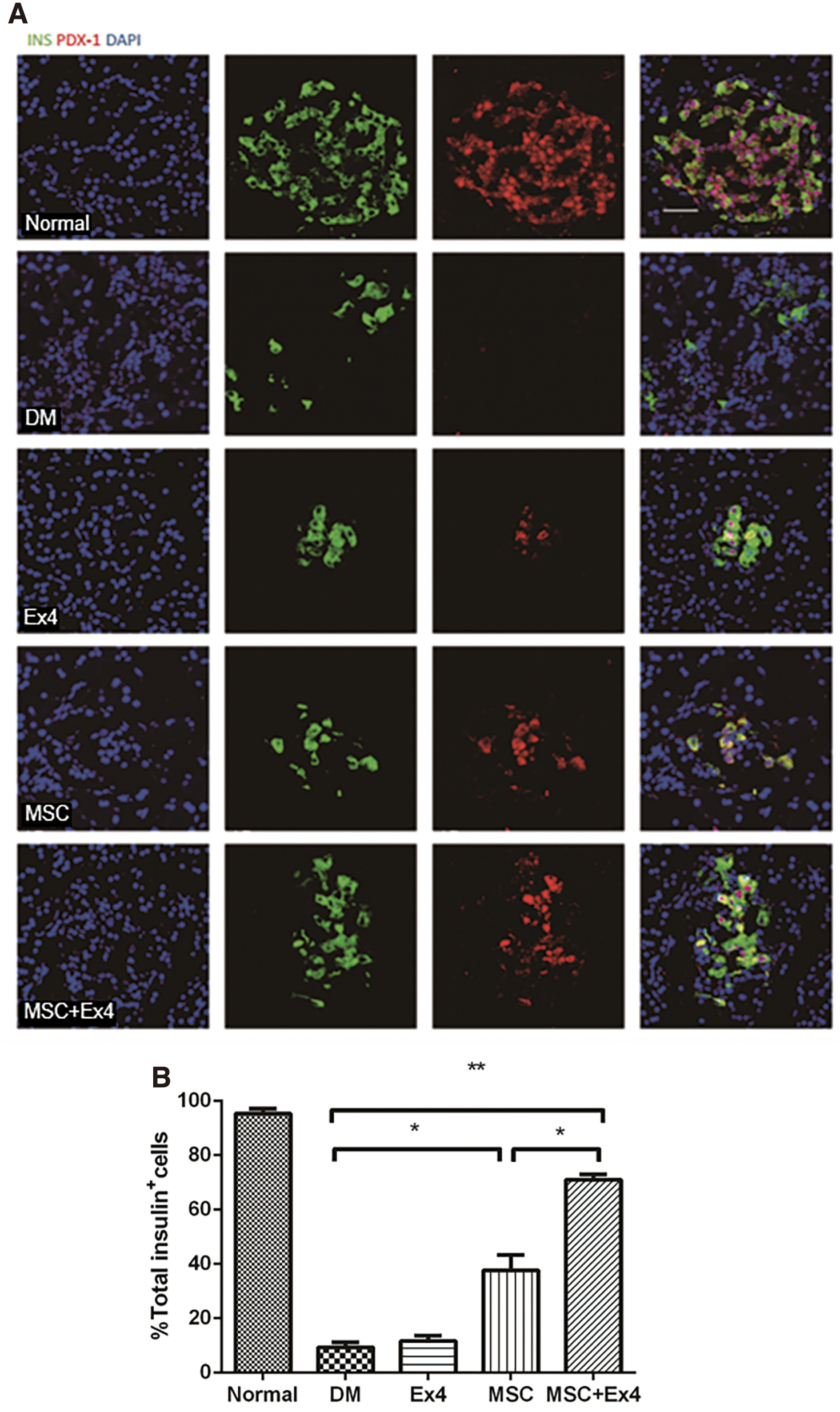
Effect of MSCs and Ex4 on PDX-1.
Similar procedures were carried out to examine the effect of MSCs and Ex4 on the expression of other nuclear transcription factors, including MafA, FoxO1, and GLP-1R (Fig. 7A). Consistently, the expression of MafA, FoxO1, and GLP-1R was significantly enhanced in the MSC and MSC+Ex4 groups compared with the other groups (Fig. 7B–D). Moreover, the β-cell expression level of MafA, FoxO1, and GLP-1R in the MSC+Ex4 group was significantly higher than those in the MSC group.
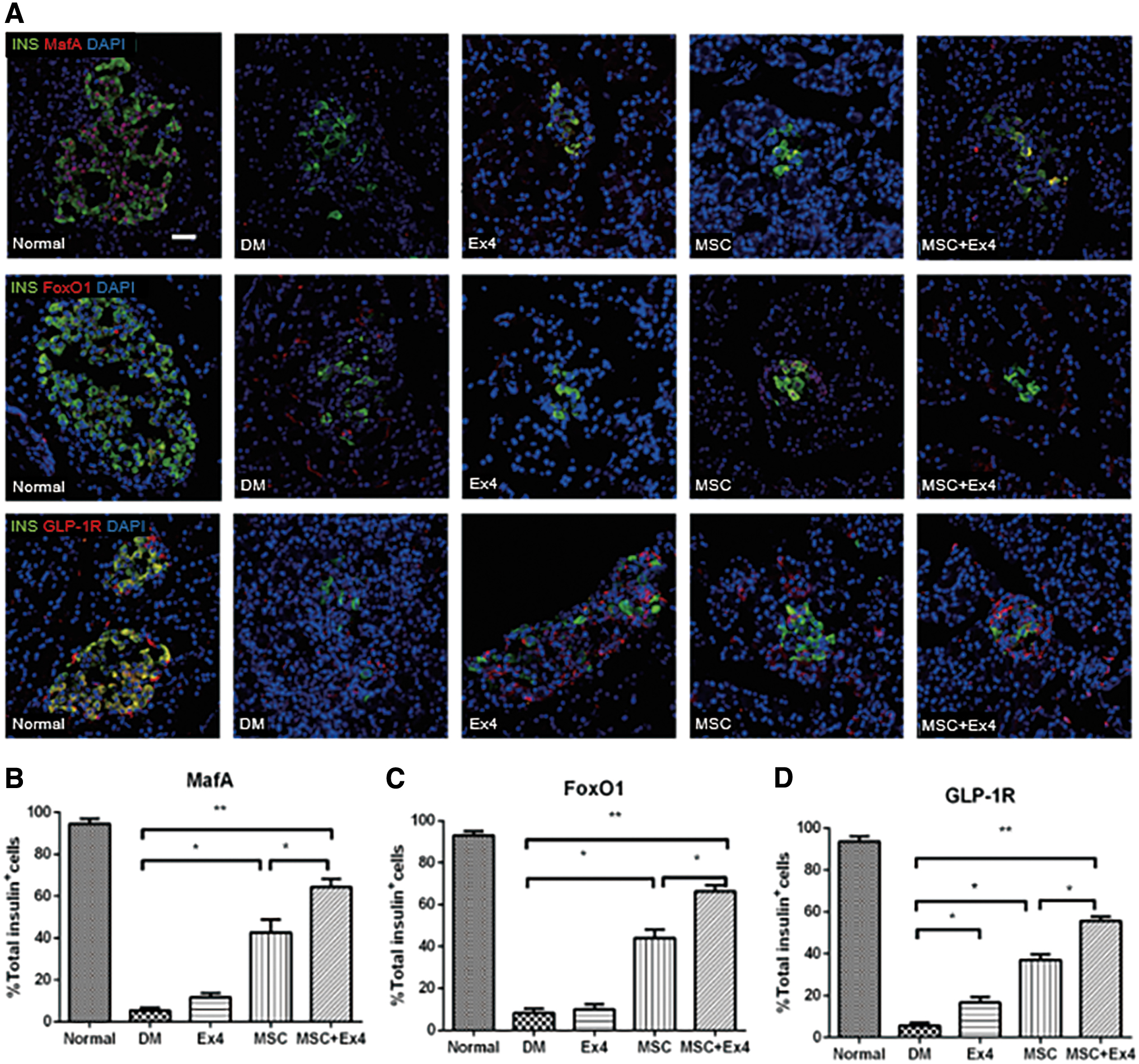
Effect of MSCs and Ex4 on MafA, FoxO1, and GLP-1R.
MSCs alone and in combination with Ex4 led to numerous insulin and glucagon double-positive cells, as well as GLP-1 and Glucagon double-positive cells
To investigate the role of GLP-1 in promoting the maturation of newly formed β-cells after MSC treatment, immunofluorescent staining of insulin, glucagon, and GLP-1 was performed on islets. The results showed that insulin and glucagon double-positive cells were present on the first day after MSC administration (Fig. 8), and multiple glucagon and GLP-1 double-positive cells were present on the following days after MSC and Ex4 combined treatment (Fig. 9).
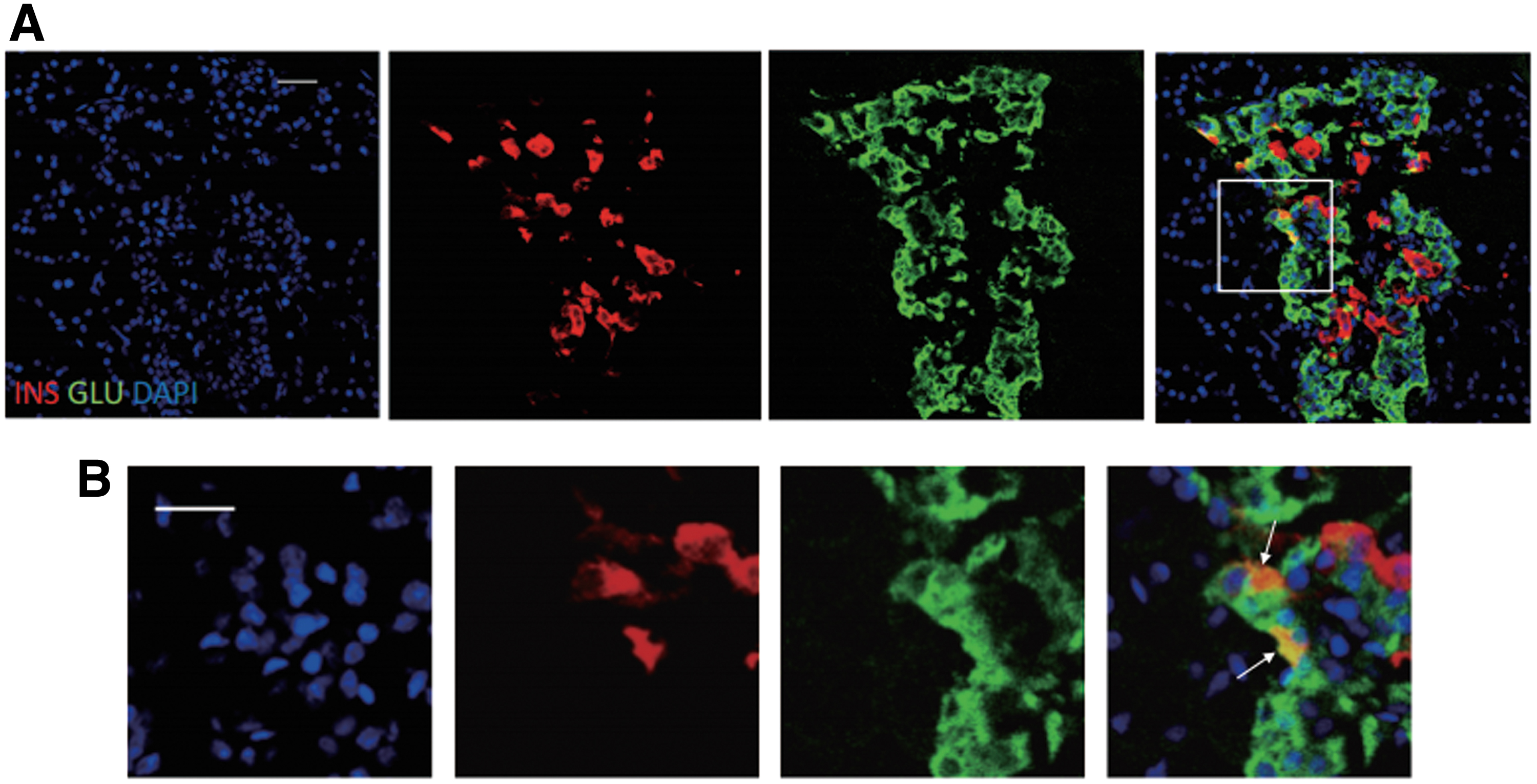
A large number of insulin+ glucagon+ double positive cells appeared 24 h after treatment with MSCs.
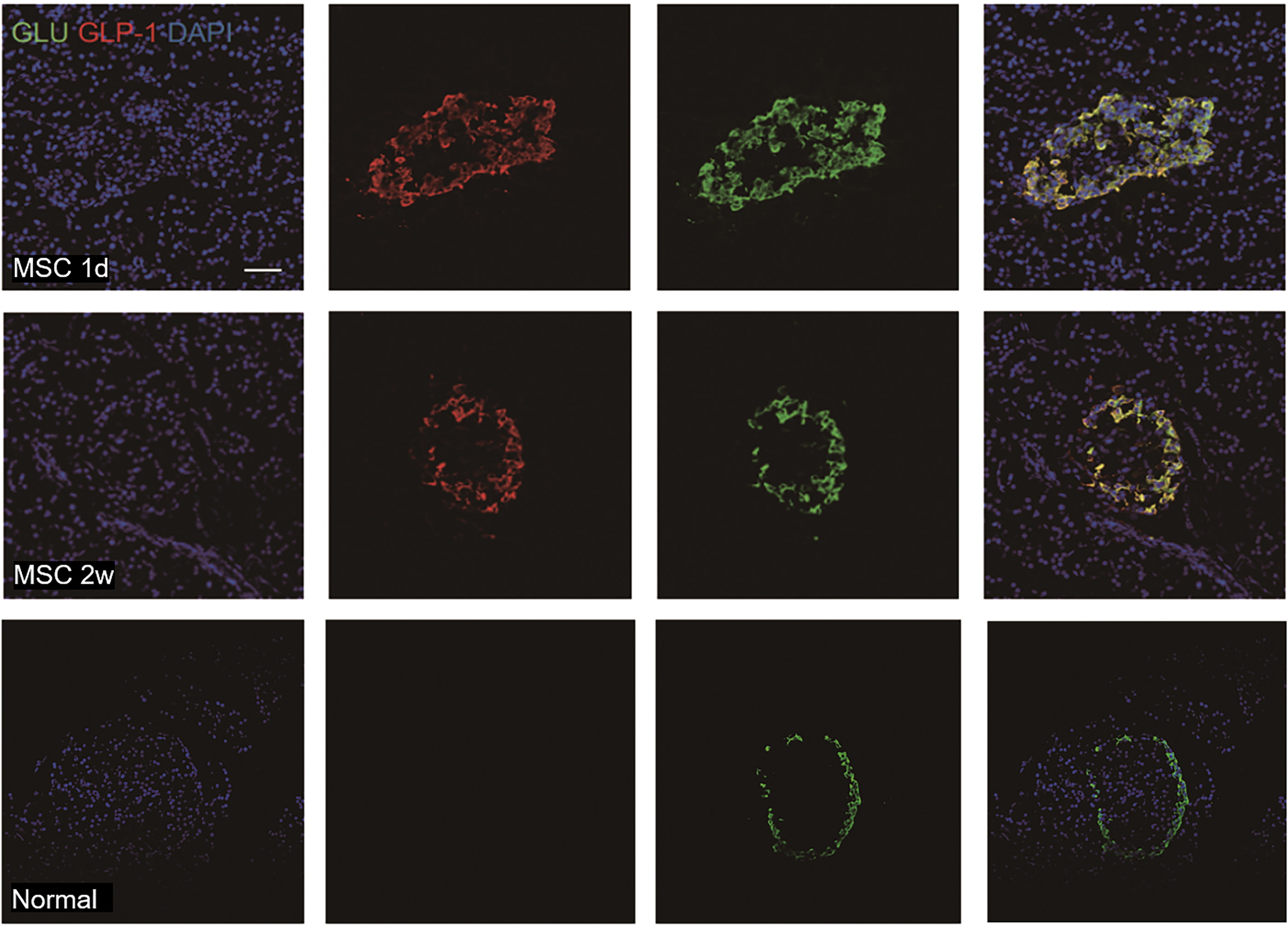
A large number of GLP-1+ glucagon+ double positive cells appeared 2 weeks after MSC treatment. GLP-1+ (red fluorescent) glucagon+ (green fluorescence) (bar = 50 μm). GLP-1, glucagon-like peptide-1. Color images are available online.
Discussion
The present study shows that combined treatment with MSC and Ex4 can effectively lower the blood glucose level than monotherapy, especially the fasting blood glucose, and promote the restoration of the number and function of pancreatic β-cells.
Diabetes is characterized by partial or complete damage to β-cell mass and results in decreased insulin secretion. Hence, targeting β-cell mass restoration and reconstruction is an important strategy in diabetic therapy.
MSCs are considered to be ideal tools for the regeneration of damaged tissues and the treatment of multiple diseases [3]. We previously reported that MSC treatment nearly normalized fasting blood glucose level by restoring newly formed β-cells in STZ-induced diabetic rats, but not by restoring random blood glucose level. In addition, considering that <3% of MSCs can differentiate into fully functional β-cells in vivo, we hypothesize that a reagent that can not only promote the growth of newly formed β-cells but also differentiate them into mature insulin-generating cells would have great therapeutic significance.
GLP-1 is able to induce the synthesis and release of insulin from pancreatic β-cells [30]. In addition, GLP-1 exerts various effects on β-cells, such as suppressing cytokine-mediated apoptosis, improving glucose-induced insulin release, enhancing neogenesis, and stimulating of cell replication and the differentiation of β-cell progenitors [23,26,32,33]. However, GLP-1 is easily degraded due to its very short half-life [34]. Hence, we focused on Ex4, an agonist of the GLP-1 receptor with slow degradation and a prolonged plasma half-life. Ex4 has been reported to be more potent than natural GLP-1 in stimulating the pancreas to synthesize insulin [35]. Therefore, Ex4 was used to explore the role of GLP-1 activation in the development and maturation of newly formed β-cells in STZ-induced diabetic rats treated with MSCs.
In the present study, STZ-induced acute and extensive β-cell loss was followed by the restoration of β-cell volume after MSCs were administered alone or in combination with Ex4, and no differences in the total numbers of β-cells were observed between the two groups. After 2 weeks treatment with MSCs and Ex4, body weight, the number of insulin-positive cells, and the serum insulin concentration were significantly increased in diabetic rats challenged with glucose, and these findings were in accordance with previous studies on the stimulatory effects of GLP-1 and Ex4 in the differentiation of rodent pancreas [18,19,25,36].
Interestingly, the level of nonfasting blood glucose in MSC+Ex4 treated rats was significantly lower than that in MSC-treated rats, while the fasting blood glucose level in the two groups was comparable. Since the total numbers of β-cells in MSC-treated rats were similar to those in MSC+Ex4-treated rats, it seemed that defects in β-cell processing or insulin generation were not the primary reason for these effects. In contrast, the extremely slow increase and reduced peak insulin concentration following glucose stimulation in the IPGTT demonstrated that MSC-treated rats were burdened with serious deficiencies in insulin secretion when challenged by normal homeostatic stimulation, suggesting that the postprandial hyperglycemia observed in MSC-treated rats was attributed to suboptimal and slow insulin secretion from β-cells after glucose stimulation.
In addition, we found that the combination of MSCs and Ex4 could better restore the function of newly formed islet β-cells than just increase the number of β-cells. These results suggested that prolonged Ex4 administration was capable of promoting insulin release and the functional restoration of newly formed β-cells in MSC-treated diabetic rats. Despite these findings, the specific mechanism by which Ex4 plays a beneficial role in the growth and differentiation of newly formed β-cells remains unknown.
Ex4 has been confirmed to be beneficial in the pancreas by maintaining the optimal quality and function of β-cells through the upregulation of gene expressions, thus inducing the neogenesis and proliferation of β-cells and suppressing the apoptosis of islet cells [37].
To clarify whether Ex4 stimulated insulin secretion by enhancing the proliferation and/or decreasing the apoptosis of newly formed β-cells, which further increased the number of newly formed β-cells, we conducted several experiments. We found no significant change in the ratio of α-cells to β-cells (α-β-cell ratio) in the islets of MSC-treated rats or MSC+Ex4-treated rats. β-cell proliferation was examined by Ki67 staining, apoptosis was measured by TUNEL staining, and the results showed that the effect of Ex4 on the proliferation and apoptosis of newly formed β-cells could not fully explain its effect on glucose-challenged insulin secretion. Thus, these findings suggested that Ex4 improved glucose-induced insulin release primarily by stimulating the function of newly formed β-cells rather than altering the number of cells.
Different doses of STZ cause different numbers of beta cell apoptosis [38]. In the diabetic rat model induced by low-dose continuous injection of STZ, the residual beta cell proliferation level was higher compared with the nondiabetic control group. However, in a high-dose single-injection diabetes model as we used, it can be seen that the beta cells have been almost completely destroyed in the untreated diabetic control group, and the remaining beta cell proliferation level is too low to be ignored.
An increase in the ratio of glucagon+ cells on total islet cells can be the result of two different events: the absolute number of alpha cells has increased or the absolute number of beta cells has decreased, or a combination of both. In this high-dose STZ treated animals the number of beta cells significantly reduced; thus, the ratio of alpha cells/(alpha+beta cells) increased correspondingly.
The eventual differentiation and maturation of β-cells require several crucial transcription factors, including PDX-1, MafA, FoxO1, and GLP-1R, which ultimately potentiate insulin release after glucose stimulation.
PDX-1 has been confirmed to be extensively involved in maintaining the normal function of β-cells and regulating glucose-induced insulin gene expression [39 –42]. In animal models, the enhanced β-cell mass is accompanied by the expression of PDX-1 in ductal cells during pancreatic proliferation [43,44].
In the present study, combined MSCs and Ex4 administration upregulated the gene expression of PDX-1 in MSC-treated rats compared to either untreated rats or MSC-treated rats, as shown by the elevated PDX-1 positive cell numbers within islets. In all islets from patients with diabetes, as well as in animal models, PDX-1 protein was localized in the cytoplasm, whereas in the nondiabetic islets, PDX-1 was in the nucleus [45 –47]. For PDX-1 to exert its transcriptional activity, it must translocate from the cytoplasm to the nucleus. In this study, PDX-1 in the MSC group and MSC+Ex4 group was more expressed in the nucleus of β cells than the DM group, indicating that its islet function has been greatly improved.
It is likely that the expression of MafA was significantly enhanced in the MSC+Ex4 group compared with the other groups. Moreover, it has been reported that FoxO1 ablation causes the loss of β-cell mass by promoting the dedifferentiation of β-cells into α-cells under metabolic stress [48]. Our study showed that the expression of FoxO1 in newly formed β-cells was increased, suggesting that Ex4 promoted the maturation of newly formed β-cells by reducing their dedifferentiation.
When GLP-1 binds to receptors on pancreatic progenitor cells, propagated proliferation and cell signal transduction are induced [49]. In GLP-1R−/− mice, islet cell proliferation was blocked after the administration of Ex4 [50]. Hence, GLP-1R in pancreatic β-cells is essential for DM treatment. In the present study, we found an increased number of GLP-1R+ cells in MSC+Ex4-treated rats compared to MSC-treated rats.
Collectively, these findings revealed that Ex4 exerts significant protective and beneficial effects on the pancreas in diabetic rats, ultimately leading to efficient and optimal insulin synthesis and secretion. However, more research is needed to further clarify the specific mechanisms by which the combined treatment of MSCs and Ex4 affects the reproduction and maturation of pancreatic β-cells.
Conclusion
In summary, the present study showed that the combined treatment of MSCs and Ex4 significantly attenuated fasting blood glucose and random blood glucose by promoting the expression of PDX-1, MafA, FoxO1, and GLP-1R in newly formed β-cells, thus contributing to the functional restoration of newly formed β-cells. Although further studies are required, the role of MSCs and Ex4 combined therapy in enhancing the regeneration and maturation of newly formed islet β-cells might provide new insights into therapeutic options for diabetes.
Footnotes
Acknowledgments
The authors sincerely acknowledge technical assistance from X.S. and Q.H. They also thank members of Y.M. and W.H. for all of their discussion over the experiment.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was financially supported by the National Basic Science and Development Program (2012CB518103) and the 863 Projects of Ministry of Science and Technology of China (2012AA020502 and 2013AA020105).