
Abstract
Cytoreductive protocols are integral both as conditioning regimens for bone marrow (BM) transplantation and as part of therapies for malignancies, but their associated comorbidities represent a long-standing clinical problem. In particular, they cause myeloablation that debilitates the physiological role of mesenchymal stem and precursor cells (MSPCs) in sustaining hematopoiesis. This review addresses the damaging impact of cytoreductive regimens on MSPCs. In addition, it discusses prospects for alleviating the resulting iatrogenic comorbidities. New insights into the structural and functional dynamics of hematopoietic stem cell (HSC) niches reveal the existence of “empty” niches and the ability of the donor-derived healthy HSCs to outcompete the defective HSCs in occupying these niches. These findings support the notion that conditioning regimens, conventionally used to ablate the recipient hematopoiesis to create space for engraftment of the donor-derived HSCs, may not be a necessity for allogeneic BM transplantation. In addition, the capacity of the MSPCs to cross-talk with HSCs, despite major histocompatibility complex disparity, and suppress graft versus host disease indicates the possibility for development of a conditioning-free, MSPCs-enhanced protocol for BM transplantation. The clinical advantage of supplementing cytoreductive protocols with MSPCs to improve autologous hematopoiesis reconstitution and alleviate cytopenia associated with chemo and radiation therapies for cancer is also discussed.
Introduction
Cytoreductive therapies are currently broadly used to treat a variety of malignancies and to induce allogeneic hematopoietic chimerism for treatment of malignant [1,2] and nonmalignant [3 –5] disorders. When this approach is used for cancers, the primary therapeutic objective is control of proliferation in the malignant growths. When this approach is used for nonmalignant diseases, the primary sought-after effect is suppression of immune and autoimmune reactions. Despite the significant therapeutic benefit of cytoreductive therapies for many fatal disorders, they may also result in serious comorbidities affecting the gastrointestinal tract, reproductive organs, pulmonary, urothelial and cardiovascular systems, and hematopoiesis [6,7]. Bone marrow (BM) aplasia and pancytopenia contribute considerably to the adverse side effects of cytoreductive protocols.
With the understanding that cytoreductive regimens damage mesenchymal stem and progenitor cells' (MSPCs) ability to sustain autologous hematopoiesis impaired by anticancer chemo and radiation therapies and engraftment of the allogeneic hematopoietic stem cells (HSC) under conditioning regimens, a search has begun for a means to protect this very important function of BM stroma.
This review provides a synopsis on the current knowledge about different mesenchymal compartment subsets within HSC niches and discusses the prospect for development of the MSPC-based protocols for: (i) supplementation into myeloablative regimens to support autologous hematopoietic reconstitution and (ii) sustaining allogeneic BM engraftment. The rationale for incorporating MSPCs into cytoreductive regimens is based on new insights on the dynamic structure of BM and the critical role of MSPCs within HSC niches under physiologic, pathologic, and therapy-associated conditions [8].
HSC Niches and BM Transplantation Challenges
The unitary theory of blood cells origin from HSCs was postulated by Alexander Maximow [9] and later was confirmed experimentally by Till and McCulloch [10]. HSCs represent a small subpopulation of cells within BM originally identified as CD34+ cells capable of hematopoietic reconstitution. The two basic features of HSCs are their ability to self-renew and to differentiate into mature blood cells. These functionally different HSC subsets are supported by long-term (LT-HSCs) and short-term (ST-HSC) repopulating HSC niches [11 –13].
Bone marrow transplantation (BMT) has been explored for the treatment of a vast variety of disorders; however, comorbidities associated with cytoreductive conditioning regimens and the risk of graft versus host disease (GVHD) limit its broad clinical application [14]. BM is the most sensitive tissue to human leukocyte antigen (HLA) disparity. In addition to major histocompatibility complex (MHC) antigens, numerous minor histocompatibility antigens play a role in allogeneic BM engraftment [15]. HSC engraftment implies that these cells have the ability to migrate into the host's BM microenvironment, known as HSC niches, and undergo self-renewal or differentiation into mature blood cells [16 –18]. This concept led to the conclusion that the host's HSC niches must be ablated to create a space for donor-derived BM to engraft [18 –20].
In conventional BMT protocols, recipient niches are damaged by ablative conditioning regimens resulting in cytopenia [21]. Different conditioning protocols are grouped into total myeloablative conditioning (MAC) [22], nonmyeloablative conditioning (NMC) [23], and reduced-intensity conditioning (RIC) [24]. In addition, monoclonal antibody therapy (MAT) has also been used to facilitate BM engraftment [25,26]. MAT is not conventionally considered as a conditioning regimen; however, it also leads to cytopenia and its associated comorbidities.
Despite the improvements in clinical outcomes achieved by reducing the conditioning intensity, the complications caused by these regimens often outweigh their therapeutic benefits. An optimal, clinically safe, and effective protocol for BMT remains to be established. Despite the associated comorbidities, myeloablation remains a standard component of the conditioning regimen required for BMT. This review presents evidence to challenge its necessity, especially as part of treatment for nonmalignant disorders.
Mesenchymal Cell Compartment of HSC Niches During Hematopoietic Homeostasis
HSC niches are complex, functionally dynamic structures sustaining either HSC self-renewal or their differentiation into mature blood cells. These two functionally discrete types of niches populated predominantly by HSCs in the dormant and proliferative status, respectively, have distinct spatial allocations within BM. HSCs migrate between these niches or exit into peripheral circulation [27,28]. They can also switch from dormancy to proliferation during homeostasis and under different stimuli [29]. Ultimately, HSCs' fate (quiescence vs. proliferation, self-renewal vs. differentiation, migration and engraftment) is regulated by their microenvironmental niches [30 –32]. Mesenchymal stem cells (MSCs), originally identified by Friedenstein, represent one of the major cellular compartments within HSC niches [33].
Research on MSC differentiation into different tissues produced a new branch of reparative medicine. Another essential property of MSCs is their immunomodulatory function [34 –36]. The focus of this review is on yet another important role of MSCs, which is in regulating hematopoiesis. The two major types of HSCs niches are endosteal (also known as long term [LT] or periarteriolar owing to their proximity to arterioles) and perivascular (also known as short term [ST] or perisinusoidal) niches, which are located around sinusoids in the inner core of BM [37]. The minority of HSCs are in a quiescent state and reside in endosteal niches, whereas the majority of HSCs, which represent a mixture of dividing and dormant cell populations, are located in perivascular niches [38,39].
Cytopenia results from impaired hematopoiesis, and cytoreductive therapy is one of its causative factors. Understanding the role of damaged mesenchymal cells within HSC niches in pathogenesis of this complication is needed to improve the outcomes of anticancer treatments and BMT-based therapies. HSCs have a vast regenerative potential. For example, replenishment of CD4+ cells, destroyed by human immunodeficiency virus (HIV), is sustained for years. On the contrary, only a few weeks of cytoreductive therapy results in cytopenia [21]. The potential key mechanisms based on compromised function of the mesenchymal stroma cells compartment in the BM, which are responsible for an abrupt impairment of hematopoietic homeostasis under cytoreductive regimens are discussed in this review.
Regulation of the HSC-MSC axis is mediated through either direct cell-to-cell interactions [31] or by secreted molecules [37,38]. The subsets of mesenchymal cells in HSC niches play different roles in hematopoiesis by regulating self-renewal and lineage-specific differentiation of HSCs in homeostasis, pathology, and postconditioning reconstitution by mediating both these mechanisms [39,40]. A brief review of the current understanding of mesenchymal cell subsets and their structural components sustaining LT- and ST-HSC niches is provided hereunder and illustrated in Fig. 1A, B, D.
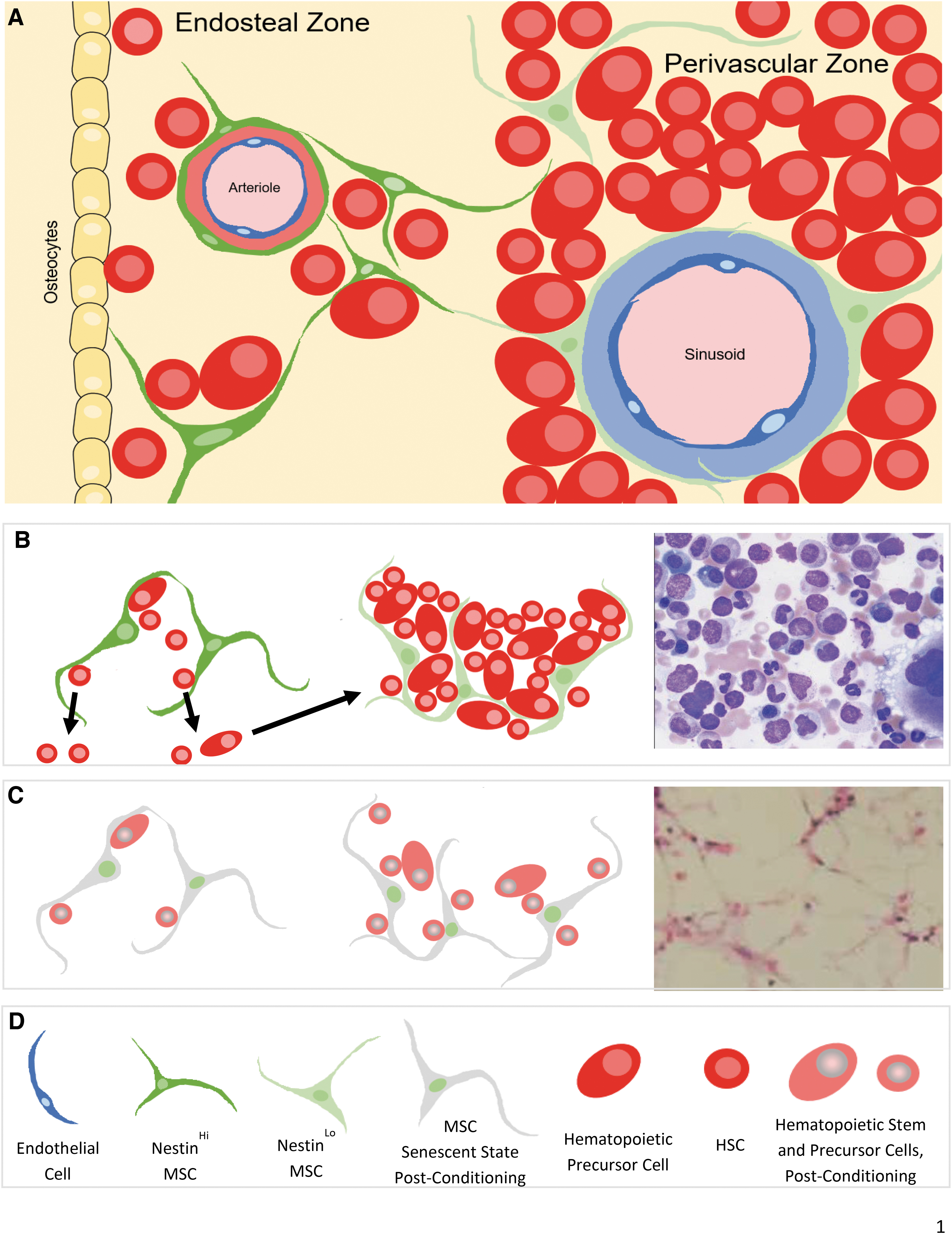
Schematic illustration of hematopoietic stem cell niche types in the bone marrow.
Endosteal HSC niches harbor mesenchymal cells, which are responsible for sustaining LT-HSCs. This includes MSPCs and mature osteoblasts [41,42]. Osteoblasts sustain LT-HSCs subsets' self-renewal ability [38,43], providing “quiescence” signaling to HSCs by direct interaction [44] or by secreting osteopontin [45]. Depending on their stage of differentiation, MSCs promote either myeloid or lymphoid differentiation [46,47]. Osteoclasts are also involved in the regulation of HSC niches and have close interactions with osteoblasts [48]. Nestin and Leptin markers define MSC subsets in both LT- and ST-HSC niches. Nestin+ cells have BM fibroblast colony-forming unit (CFU-F) activity, the ability to direct MSCs into lineage-specific differentiation, and a strong self-renewal potential. Two subtypes of Nestin+ cells have been identified in mouse BM: Nestinperi cells adjacent to arterioles and Nestinretic cells (reticular in shape) associated with sinusoids [39].
Perivascular HSC niches are located in proximity to BM sinusoids and contain a mixture of cells in dormant and proliferative states. The majority of cells in the proliferative state represent a population of lineage-committed progenitors with short-term repopulating potential; these cells are incapable of hematopoietic reconstitution in vivo [49]. CXCL12-abundant reticular (CAR) cells in perivascular niches have a high expression of CXCL12 [50 –52]. LepR is another defining marker of stromal cells supporting ST-HSC niches; this population contributes to bone regeneration after irradiation or fracture [40].
The major molecules regulating the MSC-HSC interactions are NG2, Sca-1, SCF/c-Kit, and CXCL12/CXCR4. They have different levels of expression in endosteal and perivascular niches. NG2+/NestinHi mesenchymal cells in periarteriolar niches promote HSC quiescence, and depletion of NG2+ cells reduces the long-term repopulating potential of HSCs in BM [53]. Sca-1, expressed on mesenchymal cells, plays a role in HSC self-renewal and cell lineage differentiation [17,54]. NestinHi periarteriolar mesenchymal cells are Sca-1Hi [39,55], whereas LepR+ CAR cell lack Sca-1 expression [56].
The SCF/c-Kit axis plays an essential role in HSC self-renewal [57,58]. Mutations or conditional deletion of the SCF or c-Kit locus results in hematopoietic aplasia and anemia [59]. LepR+ CAR cells show a high expression of SCF [58,60]. The CXCL12-CXCR4 axis is important for the homing of HSCs within BM [61]. CXCL12 plays a role in regulating both the quiescent and proliferative states of HSCs in strong correlation with the level of this molecule's expression. CXCL12 is about 10 times higher in CAR cells in perivascular HSC niches compared with osteoblasts in endosteal niches [60].
In summary, endosteal and perivascular HSC niches represent distinct sites preferentially harboring either LT- or ST-HSC niches. This is not a status quo condition: rather, HSCs can change their phenotype, niche type location, and proliferative versus dormancy status. This flexibility contributes to HSC regulation and to maintaining the cellular subsets balance in response to physiologic, pathologic, and therapeutic stimuli [38]. The MSCs in HSC niches, supporting either LT- or ST-HSC populations, have characteristic phenotypes (Table 1).
Mesenchymal Stem Cells with Different Profiles Are Present in the Long-Term Hematopoietic Stem Cell and Short-Term Hematopoietic Stem Cell Niches
Markers characteristic of the quiescent state profile are prevalent in the MSCs residing in the LT-HSC niches, whereas markers characteristic of the proliferative state profile are prevalent in the MSCs residing in the ST-HSC niches.
LT-HSC, long-term hematopoietic stem cell; ST-HSC, short-term hematopoietic stem cell; MSC, mesenchymal stem cell.
Impact of Myeloablation on Function of the MSC/HSC Axis
The chemo and radiation therapies for treating malignancies, the conditioning protocols for BMT, and the immunosuppressive therapies for autoimmune conditions and organ transplantation represent different modalities of cytoreductive regimens. The adverse effects of these regimens are often severe and multifactorial, affecting numerous organs. One of them is the deteriorating effect on hematopoiesis with resulting anemia, thrombocytopenia, and pancytopenia. Hematopoietic homeostasis is sustained through the balance between symmetric and asymmetric division of HSCs. Symmetrical division sustains self-renewal of HSCs. These cells primarily remain in the G0 cell cycle phase under homeostatic condition [62]. Asymmetric division produces one cell with its original properties and another with the potential for lineage-specific differentiation [12,63,64].
Numerous factors regulating these processes are altered by cytoreductive regimens that affect the physiological balance between ST- and LT-HSCs niches [65 –67]. Stromal cells in the HSC niches regulate quiescent, nonproliferative state of the HSCs, which protect them from both direct ionizing radiation (IR)-induced killing and indirect cytokine and free radical-mediated killing [68]. A study employing genetic barcode-based technology demonstrated homogenous differentiation of all engrafted HSC populations in unconditioned mice. By contrast, in mice subjected to conditioning regimes, only a small subset of HSC clones were involved in differentiation [65].
Mesenchymal cell subsets regulating hematopoiesis are damaged by cytoreductive regimens by numerous mechanisms inducing defects at the cellular, protein, and molecular levels. Osteoblasts are reported to respond to radiation by inducing HSC migration toward endosteal regions and eliciting “quiescence” signaling [69]. Anomalies in the CXCL12-CXCR4 axis were demonstrated to contribute to the pathogenesis of pancytopenia in response to myeloablation [51].
Exposure to IR induces long-term persistent senescence status in different cell lines that contributes to reduced functionality of the affected tissues [70]. This was shown to be the case for stromal BM cells as well. It was demonstrated in the mouse model that exposure to the IR triggers expression of the senescence markers and increases expression of p16 (INK4a)/p19(ARF), and that the IR-induced damage to the BM microenvironment can produce an adverse effect on hematopoiesis in an Ink4a/arf-dependent manner [71].
The studies evaluating the effects of radiation on human bone marrow mesenchymal stromal cells also demonstrated induction of the senescence status in these cells when exposed to a low dose of IR [72]. A study by Cameron et al. identified Nrf2 as an important factor in the response of normal tissues to radiation. They showed that Nrf2-mediated gene expression regulates a hematopoietic cell compartment response to radiation, where Nrf2 increases survival of irradiated osteoblasts necessary for LT-HSC niches. The absence therein of Nrf2 elevates stem cell radiosensitivity. Loss of Nrf2 promotes LT-HSCs from a quiescent to a proliferative state, which causes hematopoietic exhaustion and hence lower engraftment after myoablative irradiation [73].
Exposure to high levels of radiation is known to cause an acute radiation syndrome causing BM failure and resulting anemia, thrombocytopenia, and pancytopenia leading to life-threatening infections. The role of damaged MSCs in such conditions is supported by data providing the evidence that coculture of human macrophages with human MSCs or with exosomes isolated from MSCs can significantly enhance survival of mice exposed to lethal irradiation [74]. These and other data support the need to further explore the potential of using the MSCs to treat radiation-induced damage to different tissues [75].
A major target of low dose radiation effects are the stem cells. These cells can accumulate radiation effect during their relatively long lifespan, ultimately affecting their function. However, the common perception about MSCs is that they are relatively radioresistant. This concept can be challenged based on the data demonstrating that the radiation sensitivity of mammalian cells does not always follow a simple linear dose–effect response, with numerous cell types showing hyper-radiosensitivity to very low levels of radiation with no correlation with the cell survival response to the high doses [76,77].
Another interesting observation revealed that in the MSC cells, the number of Ki67 + cycling cells did not reduce with an increase of the radiation dose. Also unexpected was the observed lack of direct correlation between dose of radiation and number of the apoptotic cells. It was shown in the proliferative and functional acid-beta-galactosidase assays that a large percentage of the MSCs entered senescence and exhibited radiosensitivity in response to very low dose of radiation (40 mGy) while exhibiting radioresistance in response to higher doses of radiation (up to 2000 mGy) [72].
Unlike discrepancies regarding the role of the mesenchymal components in the HSC niches in compromised hematopoiesis, the fact that radiation exposure impairs hematopoiesis is undeniable. New therapeutic modalities are being developed to protect, reverse, or, preferably, prevent BM damage associated with exposure to radiation [78]. Conventionally, the BM susceptibility to radiation-induced damage was attributed to high proliferative activity of the hematopoietic stem and precursor cells, as cells with high cell division rates are known to be more radiosensitive [79]. Numerous mechanisms reportedly contribute to the HSC damage in response to radiation, such as TP53-mediated apoptosis, telomere dysfunction affecting lineage differentiation, and induction of senescence [80 –82]. Mesenchymal stromal cells, which play a paramount role in sustaining hematopoiesis in the HSC niches in physiologic condition, are reported in some studies to be less radiosensitive compared to HSC [83,84]. Thus MSCs have been suggested as viable candidates for cell-based therapy of cytoreductive regimen-induced BM aplasia [85,86].
It was shown that MSCs can support hematopoiesis in patients with acute radiation syndrome as MSCs promote proliferation of irradiated CD34+ HSCs, rescue these cells from radiation-induced apoptosis, and support hematopoietic reconstitution after coculture [87]. Other studies have demonstrated that radioprotection can be delivered through MSCs-induced increased secretion of the hematopoietic growth factor Flt3L [88]. Preclinical and clinical trials show the effectiveness of the MSCs in sustaining the HSCs engraftment in BMT patients [89,90]. The mechanisms of action are suggested to be the secretion of cytokines and growth factors that stimulate hematopoiesis, combined with inhibition of T and dendritic cell-based responses [91,92].
The studies exploring the potential therapeutic applications of the donor-derived MSCs are evolving from the utilization of direct MSC-based therapeutic protocols to the utilization of MSC-educated macrophages and MSC-exosome educated macrophages as adoptive cell therapies for radiation-induced malfunction of mesenchymal stromal cells in the HSC niches in the BM [93].
Chemotherapy is another iatrogenic tool, which, like radiation, has a cytoreductive effect with potential to cause BM aplasia and associated comorbidities. Hematopoietic recovery after chemotherapy can be one of the most challenging aspects of cancer patient management. One of the potential pathogenic mechanisms of flawed hematopoiesis under this therapy is chemotherapy-induced damage of the stromal components of the BM. It was shown over 20 years ago by estimation of the frequency of the CFU-F in the BM of the recipients of BMT that the BM stromal microenvironment is seriously and irreversibly damaged after BMT (based on 12-year-long follow-up) [94].
Kemp et al. compared the phenotype and functional properties of BM-derived MSCs from untreated patients with hematological malignancy and those treated with high-dose chemotherapy (HDC). This study demonstrated a significant reduction in MSC expansion and MSC CD44 expression by MSCs derived from patients receiving HDC regimens, thus implicating disadvantage in the use of the autologous MSCs in chemotherapeutically treated patients [95]. Another study by this group demonstrated that the exposure of MSCs to the chemotherapeutic agents cyclophosphamide and melphalan has strong negative effect on MSC expansion and CD44 expression. In addition, the functional injury of the MSCs was reflected by their decreased ability to support hematopoietic cell migration and repopulation [96].
The commonly used chemotherapy drugs cytarabine and daunorubicin were tested for their effect on BM-residing MSCs and drug-induced alteration of hematopoiesis-supportive function of these cells. The chemotherapeutic drugs induced altered ability of the MSCs to support hematopoietic reconstitution was attributed to increased intracellular level of reactive oxygen species and induction of cell apoptosis [97]. Somaiah et al. evaluated the effect of the chemotherapeutic drugs cytarabine, daunorubicin, and vincristine on differentiation, phenotypic and gene expression changes in MSCs, and tested the anticancer effects of drug-treated MSC on leukemia cells. The findings in this study demonstrated that the assayed drugs altered the phenotype and the osteogenic and adipogenic differentiation of MSCs and rendered leukemic cells in these patients more chemo resistant [98].
Thus, the majority of studies are in agreement that the chemotherapeutic drugs have negative effects on MSCs' various functions. However, the recent in vitro study by Lopez Perez et al. demonstrated that human BM-derived MSCs are relatively resistant to treatment with the widely used anti-metabolite drugs, 5-FU and gemcitabine. In this study the expression of the MSC surface markers and their multilineage differentiation potential was shown to be irrespective of 5-FU or gemcitabine treatment. The high expression levels of enzymes involved in DNA metabolism and multidrug resistance transporters in the MSCs addressing contributing factors for the observed readouts. However, the authors did not address in this study the ability of the MSCs to support hematopoiesis [99].
An additional confirmation for the observation that MSCs in the senescent state have decreased ability to support hematopoiesis came from a study by Yuan et al. This study showed that HIV protein p55-gag could induce senescence of MSCs, resulting in their reduced capacity to support expansion of HSCs in vitro [100].
Another correlation between senescent state in the MSCs and their impaired ability to sustain hematopoiesis was demonstrated in patients with myelodysplastic syndromes (MDS). MDS-derived MSCs were found to have significantly reduced osteogenic differentiation and altered expression of key molecules involved in the interaction with HSCs, such as Osteopontin, Jagged1, Kit-ligand, and Angiopoietin, and several chemokines. In addition, the MDS-derived MSCs had significantly diminished ability to support CD34+ HSCs in long-term culture [101]. In line with this finding, deficient hematopoiesis was detected in myeloma BM-derived MSCs, which also was shown in association with their senescent state. Specifically, it was shown that myeloma BM mesenchymal stromal cells have an increased expression of senescence-associated β-galactosidase, increased cell size, reduced proliferation capacity, and characteristic expression of senescence-associated secretory profile members [102].
BM aplasia associated with cytoreductive regimen-induced senescence status in the mesenchymal stromal cells is given in Fig. 1C and D.
Challenging the Niche-Creation Dogma for Allogeneic HSC Engraftment: Is Myeloablation Required for Allogeneic HSC Engraftment?
A long-held perception is that ablation of autologous hematopoiesis is required for a successful allogeneic BMT [32,65]. To minimize the adverse effects, conditioning regimens have shifted from MAC to NMC and RIC. Although outcomes have improved, the complications remain significant [65,66]. BMT conditioning protocols damage HSC niches including their mesenchymal cell components resulting in different levels of pancytopenia [103,104].
A question arises as to whether BM aplasia and pancytopenia resulting from cytoreductive therapy are owing to the impaired function of MSPCs sustaining hematopoiesis. Could damage to MSPCs be the key factor responsible for both (i) impaired autologous hematopoietic reconstitution in case of chemo and radiation therapies for malignances and (ii) hindered engraftment of donor HSCs in case of BMT? Does myeloablation damage the niche-stromal components and impair their ability to support hematopoiesis rather than clearing the HSC niche space for donor BM engraftment? We would like to address these queries by discussing two intertwined questions: what is the current view on cellular and molecular level impacts of myeloablation on hematopoiesis, and what are the ablation requirements for HSC engraftment? In addition, a question about the role of MSPCs in the prevention of GVHD is discussed.
Conditioning, as a requirement for BMT, was reexamined after evidence emerged of its damaging effect on the mesenchymal compartment, which has the vital function for supporting hematopoiesis in HSC niches [65,105]. Numerous studies demonstrated the possibility of induction of hematopoietic chimerism without host myeloablation [106 –109] and the possibility that dietary valine restriction may reduce iatrogenic complications in HSC transplantation [110]. It was shown that HSC niches are dynamic, rather than invariable, structures [28,111]. Healthy donor-derived HSCs have the ability to outcompete diseased host HSCs for niche space during engraftment [107,112]. In addition, empty HSC niches appear to be available for engraftment of donor-derived HSCs [113].
Administration of massive doses of BM cells was shown to secure hematopoietic engraftment without conditioning [106,111,114]. The stem/precursor cell ratio and genetic composition of transplanted BM were also shown to be significant factors in promoting allogeneic engraftment. A competitive repopulation assays demonstrated that low numbers of HSCs successfully compete with the greater numbers of cotransplanted short-term precursor cells [115]. In addition, wild-type (healthy) donor HSCs can replace genetically defective HSCs in the recipients BM [112]. Unconditioned BMT transplantation was shown to minimally perturb natural hematopoiesis [65]. Taken together, these data support the rational for further exploration of protocols for allogeneic HSCs transplantation without conditioning.
Potential Advantage in Adaptation of the MSPCs into Therapeutic Protocols
MSPC-based protocols to support autologous hematopoiesis under cytoreductive therapy
Is it possible to avoid pancytopenia associated with cytoreductive regimens for treatment of malignancies? MSCs play a key role in LT- and ST-HSC niche integrity [46,53,116,117]. Radiation exposure, chemotherapeutic protocols, and other cytoreductive regimens can induce the senescent state and thus play a significant role in impaired ability of MSCs to sustain HSC niche integrity and resulting pancytopenia [71,118]. Incorporating MSPCs into protocols of the cytoreductive regimens may, therefore, be a key factor for treatment and prevention of cytopenia. Further research is needed to determine which subset of the mesenchymal cells or their combination would be optimal for such therapies.
MSPC-based protocol to sustain allogeneic HSC engraftment in a conditioning-free BMT
Mesenchymal cells represent a strong candidate for sustaining a conditioning-free allogeneic HSC engraftment without GVHD. As a major cellular component of HSC niches, MSPCs sustain hematopoiesis under homeostatic conditions; when they are damaged by conditioning regimens, their ability to sustain engraftment of allogeneic HSCs is impaired. The ability of MSPCs to cross-talk with HSCs despite allogeneic disparity [119 –121] allows these donor-derived MSPCs to resume the impaired hematopoiesis-supporting function of the stromal component in the HSC niches and additionally enables the use of the high doses of MSPCs from unrelated donors or commercial sources [122].
Amelioration and prevention of GVHD
Despite the accumulating evidence supporting the prospect of a clinically successful BMT protocol without conditioning [108,123], its clinical adaptation is hindered by the potential complication of GVHD [124,125]. Transplantation of purified HSCs was attempted as a venue to avoid GVHD. However, it significantly reduced HSC engraftment and induction of allogeneic chimerism [126]. To improve engraftment of allogeneic HSCs, while avoiding GVHD, a large cohort of hematopoietic [127,128] and nonhematopoietic cells were studied [52,56,129 –131], including osteoblasts, CAR cells, and MSPC populations. However, an adverse-effect-free protocol for induction of allogeneic hematopoietic chimerism remains to be developed.
The concept that MSCs can talk with HSCs across the MHC disparity was first shown by A. J. Friedenstein over 50 years ago when he identified MSCs in the bone marrow [33]. In 1982, T.M. Dexter foreseeably asked: “Is it possible that successful transplantation requires transfusion not only of haematopoietic stem cells but also transfusion and engraftment of stromal cell precursors? We will no doubt know the answer in the near future” [132].
Since then, the insight on phenotype and functions of the MSCs has advanced significantly. It was confirmed that MSCs can be used irrespective of HLA compatibility, as they do not trigger the immunologic reactions. For a while, the absence of the antigenicity of the MSCs was explained by the limited HLA class I expression and lack of expression of the HLA class II and costimulatory molecules CD40, CD40 ligand, CD80, and CD86 [133 –135]. It was suggested that, in the absence of costimulatory molecules, the secondary signal would not be activated, leading to induction of the T cell anergy [136 –138]. It was demonstrated that MSCs do not elicit proliferative responses from allogeneic T cells in culture [139] and that allogeneic MSCs transplantation is feasible and safe in various animal models [140 –144] and in humans [134,145].
Numerous clinical trials [146,147 –149] and veterinary practice studies [140,144,150] exploring the therapeutic adaptation of the MSCs in a variety of malignant [151,152] and nonmalignant [134,146,153,154] disorders prompted further studies aiming at elucidating the mechanisms underlying their toleranogenic properties. Klyushnenkova et al. showed that immunotoleranogenic properties of the MSCs are not owing to the lack of the costimulatory molecules expression, but to their ability to suppress T cell proliferation [9]. Treatment of MSCs in this study with interferon (INF), or transduction with B7-1 or B7-2, did not enhance the ability of these cells to stimulate alloreactive T cell proliferation. The ability of the T cells, removed from the culture with MSCs, to proliferate in response to stimulation with allogeneic peripheral blood mononuclear cells supported the conclusion that MSCs have immunosuppressive property irrelevant to the lack of the costimulatory molecule expression.
Suppression of alloreactivity by MSCs has been reported in humans [136] and in baboon [155] and mouse models [156]. In addition, it was demonstrated that MSCs can be immunoprivileged and immunosuppressive. They are incapable of inducing allogeneic lymphocytes proliferation in mixed lymphocyte culture, have inhibitory effect on lymphocyte proliferation, and secrete anti-inflammatory cytokines IL-10 and TGF-β [157].
Further studies of the immunomodulatory mechanisms of the MSCs revealed that these cells have potent immunoregulatory abilities by interacting with cells of the adaptive and innate immune system. They inhibit the differentiation of T cells into T helper 17 (Th17) cells and repress their proliferation while promoting generation of T regulatory (Treg) cells. The bi-directional immunoregulatory dialog between MSCs and Th17 cells is mediated by soluble factors, extracellular vesicles (EVs), mRNAs, miRNAs, cell-to-cell contact, and the transfer of organelles [158].
Incorporation of MSPCs into BMT protocol presents additional significant clinical advantage owing to their capacity to suppress GVHD [159 –161]. GVHD is a potentially life-threatening complication of allogeneic BMT, and is one of the major factors limiting the broad application of this therapeutic modality for hematological malignancies [162] and nonmalignant disorders [163 –165]. GVHD is classified into acute and chronic GVHD (aGVHD and cGVHD, respectively) [166].
Adaptation of the MSCs has been reported to be an effective therapeutic tool for the treatment, amelioration, and even prevention of GVHD. The mechanisms of the immunomodulatory effects of the MSCs mediating these clinically beneficial outcomes are under intensive research [149,167 –169]. It was shown that amelioration of the inflammatory reactions in GVHD by MSCs is associated with increased Treg cell populations and decreased secretion of the proinflammatory cytokines. The suggested mediators of these outcomes are MSC-secreted TSG6, IL-10, PGE2, vascular endothelial growth factor, and TGF-β1 [160].
Further modifications of the MSC-based protocols for amelioration of GVHD revealed the potential for substitution of the cellular inoculum of the MSCs by BM-MSC-derived EVs, administration of which recapitulated the therapeutic effects of the cell-based protocols. Systemic infusion of human BM-MSC-derived EVs was shown to prolong the survival of mice with aGVHD in association with reduced multiorgan damage, preservation of Treg cell population, and suppression of the naive T cells differentiation from naive to effector cell phenotype [170].
It was also shown that MSCs have the ability to enhance hematopoietic engraftment and hematological recovery after both autologous [171] and allogenic [151,172] BMT. This property may become crucial in the cytoreductive regimens, specifically not only in improving the allogeneic BM engraftment, but also in supporting autologous hematopoiesis reconstitution after its damage by chemo and radiation therapies for cancers. Koc et al. were the first to report improvement of hematopoietic engraftment when autologous BM-MSCs were cotransplanted with HSCs [171]. These findings were confirmed by results from the recent preclinical studies where MSCs were able to delay the onset of GVHD [173,174].
Newly revealed additional mechanisms contributing to the preventive effect of MSCs on GVHD provide an explanation for an existing paradox of the observed immunomodulatory effect after MSC infusion in association with failure of their long-term engraftment. The proposed explanation is that MSC immunomodulation depends on interaction with cytotoxic NK and T cells of the recipient. The apoptosis of the transplanted MSCs triggered by these immune cells is, according to this study, a requirement for the immunosuppressive outcome of MSC transplantation. Additional confirmation for this theory is the finding that the cytotoxicity of the host's cells and resulting apoptosis of the transfuse MSCs directly correlate with the immunomodulatory effect of these cells [173].
In disagreement with the numerous reports regarding the effectiveness of MSCs in ameliorating GVHD, a recently published analysis of over two-dozen large randomized controlled trials with enrollment of patients with hematologic malignancies concluded that MSC-based therapy to ameliorate aGVHD was not effective [175]. Indeed, outcomes are variable in different institutions and among the patients subjected to MSC-based protocols for therapy of GVHD. To eliminate these discrepancies and improve consistency in therapeutic outcomes, new clinical protocols need to be developed that define the optimal origin of the MSCs, the dose for their administration, and the cellular composition based on research that elucidates the mechanisms underlying their GVHD suppressive effects [176].
Summary and Conclusions
MSPCs have wide therapeutic potential. This review discussed their role in sustaining autologous and allogeneic hematopoiesis in physiologic condition and under cytoreductive therapies. MSPCs have been shown to play a critical role in hematopoietic homeostasis. Cytoreductive therapies appear to damage their ability to sustain hematopoiesis and contribute to BM aplasia and the onset of cytopenia [53,62,104,160]. These data suggest that incorporating MSPCs into chemo, radiation, and immunosuppressive therapies could sustain autologous hematopoietic reconstitution and alleviate cytopenia, comorbidity almost unavoidable in association with these therapeutic regimens.
MSPCs also hold therapeutic potential for improving outcomes of allogeneic hematopoietic chimerism. HSC niches are dynamic structures. Empty niches that self- and donor-derived HSCs can occupy are available within BM [114]. Healthy hematopoietic donor cells can outcompete the diseased recipient's HSCs to occupy recipient HSC niches [107,113]. These findings support the notion that the need for “clearing space” for BM engraftment is a misconceived paradigm and that conditioning can be eliminated from BMT procedure. The ability of mesenchymal cells to: (i) sustain allogeneic hematopoiesis across MHC disparity and (ii) prevent GVHD make them a strong candidate for the development of an MSPC-supplemented, conditioning-free BMT protocol [6].
The mesenchymal cell subsets, based on their phenotype, stage of maturation, and source of origin (eg, BM, umbilical cord, adipose tissue) as well as the cell dosage and MSPC/HSC ratio for supplementation into cytoreductive regimens is yet to be identified. Further exploration of the MSPC potential to compensate the function of mesenchymal stroma damaged by cytoreductive regimens could lead to new MSPC-supplemented therapeutic regimens for malignancies. Development of the MSPC-based conditioning-free protocol for BMT could open a new venue for treatment of nonmalignant disorders.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Acknowledgments
The author thanks Paul Hunter for help with bibliography and Jennifer Wilson and Dmitri Zorine with preparation of the article.
Funding Information
No funding was received for this work.