
Abstract
Declined function of aged mesenchymal stem cells (MSCs) diminishes the benefits of cell therapy for myocardial infarction (MI). Our previous study has demonstrated that SRT1720, a specific SIRT1 activator, could protect aged human MSCs (hMSCs) against apoptosis. The purpose of the present study was to investigate the role of mitochondria in the antiapoptotic effects of SRT1720. In addition, we established a nonhuman primate MI model to evaluate cell engraftment of SRT1720-pretreated aged hMSCs (SRT1720-OMSCs). A hydrogen peroxide (H2O2)-induced apoptosis model was established in vitro to mimic MI microenvironment. Compared with vehicle-treated aged hMSCs (Vehicle-OMSCs), SRT1720-OMSCs showed alleviated apoptosis level, significantly decreased caspase-3 and caspase-9 activation, and reduced release of cytochrome c when subjected to H2O2 treatment. Mitochondrial contents were compared between young and aged hMSCs and our data showed that aged hMSCs had lower mitochondrial DNA (mtDNA) copy numbers and protein expression levels of components of the mitochondrial electron transport chain (ETC) than young hMSCs. Also, treatment with SRT1720 resulted in enhanced MitoTracker staining, increased mtDNA levels and expression of mitochondrial ETC components in aged hMSCs. Furthermore, SRT1720-OMSCs exhibited elevated mitochondrial respiratory capacity and higher mitochondrial membrane potential. In vivo study demonstrated that SRT1720-OMSCs had higher engraftment rates than Vehicle-OMSCs at 3 days after transplantation into the infarcted nonhuman primate hearts. Taken together, these results suggest that SRT1720 promotes mitochondrial biogenesis and function of aged hMSCs, which is involved in its protective effects against H2O2-induced apoptosis. These findings encourage further exploration of the optimization of aged stem cells function via regulating mitochondrial function.
Introduction
Myocardial infarction (MI) causes high mortality worldwide [1] and transplantation of mesenchymal stem cells (MSCs) has emerged as a promising approach for its treatment. Although growing evidence has shown the safety of MSCs therapies for cardiac repair, the therapeutic effects of MSCs transplantation is unsatisfactory as recent meta-analyses of randomized clinical trials reported the increase in left ventricle (LV) systolic function of MI patients with MSCs transplantation is only 3%–10% [2 –4]. One of the main factors limiting the efficacy of MSCs-based therapy of MI is the poor survival rate of grafted cells in host myocardium with the majority of the cells dying shortly after transplantation [5].
In addition, when aged MSCs are used as a major source of autologous stem cell therapy for elderly people suffering from ischemic heart disease, their stress resistance is significantly decreased, and hence, cardioreparative capabilities are further diminished [6,7]. Therefore, effective methods to improve the survival abilities of aged MSCs are needed.
As the major generators of energy in our cells, mitochondria are plastic and dynamic organelles that maintain their homeostasis and function through quality control events that include mitochondrial biogenesis, mitochondrial dynamics via highly organized fusion and fission process, and mitophagy to clear damaged ones [8,9]. Mitochondrial dysfunction is associated with age-related diseases and stem cell aging [10,11]. During the process of aging, mitochondria undergo morphologic and functional changes, including the appearance of roundish mitochondria, reduction of mitochondrial DNA (mtDNA) but increase of mutations, decreased respiratory chain complex activities, as well as impaired mitochondrial biogenesis [12 –16]. Moreover, mitochondria are pivotal integrators of apoptosis and directly participate in the programmed cell death signaling through the well-defined mitochondrial pathway [17].
The mammalian silent information regulator 2 homolog 1 (SIRT1), an NAD+-dependent protein deacetylase, plays a critical role in regulating longevity, apoptosis, DNA repair, and metabolism [18]. Also, regulating SIRT1 expression and activity could have a significant influence on mitochondrial function [19]. Indeed, SIRT1 activation could stimulate mitochondrial biogenesis in conditions of energy deficiency associated with disease and tissue injury. For example, the putative SIRT1 activator resveratrol has been extensively studied and demonstrated to promote mitochondrial biogenesis and exert protective effects in animal models of cardiovascular disease, neurodegeneration, and metabolic syndrome [20 –25].
In prior studies, we have demonstrated that pretreatment with SRT1720, a selective synthetic SIRT1 activator, could protect aged human MSCs (hMSCs) against cell apoptosis [26]. Given the importance of SIRT1 in the control of mitochondrial function, we here hypothesized that SRT1720 treatment might improve mitochondrial function of aged hMSCs, and mitochondria-mediated apoptosis pathway might play a role in its protective effects. In addition, we established a nonhuman primate MI model to evaluate the efficacy of SRT1720 pretreatment strategy of aged hMSCs transplantation.
Materials and Methods
Ethics statement
All experiments involving live animals were performed in accordance with the Guide for the Care and Use of Laboratory Animals published by the United States National Institutes of Health (NIH Publication No.85-23, revised 1996) and were approved by the Institutional Animal Care and Use Committee of the Second Affiliated Hospital of Zhejiang University School of Medicine. Cynomolgus monkeys (male, 5–6 years old, 4–7 kg body weight) were purchased from Suzhou Xishan Zhongke Laboratory Animal Co. Ltd., which had been certificated by the Association for Assessment and Accreditation of Laboratory Animal Care. Bone marrow-derived hMSCs were obtained from iliac crest aspirates from donors after informed consent was obtained and were used in compliance with the procedures approved by the Human Experimentation and Ethics Committee of the Second Affiliated Hospital of Zhejiang University School of Medicine.
MSCs isolation and culture
Bone marrow aspirates were obtained during orthopedic surgery involving exposure of the iliac crest from old patients (>65 years old) after informed consent was obtained. hMSCs were harvested and isolated using the direct plating method as described previously [27]. In brief, bone marrow samples were diluted with three equal volumes of growth medium containing Dulbecco's modified Eagle medium (DMEM; Corning, Manassas, VA) supplemented with 10% fetal bovine serum (Life Technologies, Paisley, United Kingdom) and distributed equally across 10 cm dishes. The containers were returned to the 5% CO2 incubator and cultured at 37°C for 48 h. The old medium was aspirated away and then the culture medium was changed every 3–4 days. Small MSCs colonies of attached spindle-shaped cells were visible after 5–7 days. After 12–14 days, the small colonies were easily found and became confluent. At this time point, the cells were subcultured and repeatedly passaged 4–5 times before use in subsequent experiments.
MSCs immunophenotyping
MSCs were analyzed for membrane markers by flow cytometry, which included CD29, CD44, CD90, CD105, CD34, and CD45 (eBioscience, San Diego, CA). After detachment, cells were resuspended at 2 × 106 cells/mL in phosphate-buffered saline (PBS) and fluorescent conjugated antibodies against the cell surface markers were added and incubated at room temperature for 30 min. After that, cells were centrifuged at 800g for 5 min, supernatant was removed to eliminate unbound antibodies, and cells were resuspended in 500 μL PBS. Surface marker expression was evaluated with a BD FACSCanto II Flow Cytometer (BD Biosciences, San Jose, CA).
MSCs differentiation
MSCs multipotency was evaluated by osteogenic, adipogenic, and chondrogenic differentiation experiments. MSCs differentiation induction media were prepared according to previously published protocols [28]. MSCs were seeded onto a six-well plate at a density of 2 × 105 cells per well. Induction media were added and changed every 2 days. Osteogenic, adipogenic, and chondrogenic stimulation were conducted for 21 days. For microscopic evaluation, cells were fixed with 4% paraformaldehyde for 10 min and stained for 20 min at room temperature with the following stainings: matrix calcification, which is present following osteogenesis, was visualized with Alizarin Red (Cyagen, Guangzhou, China); adipogenesis was verified with Oil Red O (Sigma-Aldrich) staining; and finally, chondrogenesis was stained with Toluidine Blue (Sigma-Aldrich). Excess stain was washed away with PBS, and stained cells were visualized under a light microscope.
SRT1720 treatment
MSCs at about 80% confluence were treated with 0.5 μM SRT1720 (Selleck Chemicals, Houston, TX) for 24 h. This pretreatment condition was selected based on our previous experiments, which validated the optimal concentration and incubation time for SRT1720 to exert its protective effects against hydrogen peroxide (H2O2)-induced cell death [26]. SRT1720 was dissolved with dimethyl sulfoxide (DMSO) before MSCs treatment. Also, cells in the control vehicle group were treated with the same final concentration of DMSO.
Apoptosis model
Apoptosis model was induced by H2O2 and serum deprivation as described previously [26]. In brief, the cells were washed with serum-free DMEM and subsequently the culture medium was replaced with serum-free DMEM supplemented with 0.5 mM H2O2 and then the cells were placed at 37°C under normoxic conditions for 3 h.
Annexin V/propidium iodide staining
The binding of annexin V to externalized phosphatidylserine was used as a measurement of cell apoptosis. After H2O2 treatment, MSCs apoptosis was detected with an annexin V-propidium iodide (PI) apoptosis detection kit (LiankeBio, Hangzhou, China) according to the manufacturer's instructions. In brief, 5 × 105 cells were collected in each tube and 500 μL binding buffer was added followed by thorough mixing. Subsequently, 5 μL annexin V and 10 μL PI were added. After mixing, the tube was incubated in the dark at 37°C for 15 min. Then cell apoptosis was evaluated by flow cytometry.
Protein sample preparation and western blotting
Western blot analysis was performed as described previously [29]. MSCs were harvested and lysed in RIPA solution (Beyotime, Shanghai, China) supplemented with a protease inhibitor cocktail (Roche, Base, Switzerland). Mitochondrial and cytosolic fractions were isolated by a Cell Mitochondria Isolation Kit (Beyotime), which was used according to the manufacturer's instructions to evaluate cytosolic cytochrome c levels. Protein concentrations were normalized following protein assays with a BCA Protein Assay Kit (Thermo Scientific, Rockford, IL). Equal amounts of protein were separated by electrophoresis on 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and sequentially transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA). After blocking with 5% (w/v) nonfat dry milk (BD Biosciences, Franklin Lakes, NJ) in Tris-buffered saline with 0.1% Tween20 for 1 h at room temperature, membranes were incubated with respective primary antibodies overnight at 4°C. The blots were then incubated with the appropriate horseradish peroxidase-conjugated secondary antibodies. After washing of the blots, the bands were visualized with an enhanced chemiluminescence system (Millipore, Boston, MA).
The following primary antibodies were used: Caspase-3 (1:1,000; Cell Signaling Technology, Danvers, MA), Cleaved caspase-3 (1:500; Cell Signaling Technology), Caspase-9 (1:1,000; Cell Signaling Technology), Cleaved caspase-9 (1:500; Cell Signaling Technology), Cytochrome c (1:1,000; BD Biosciences, Franklin Lakes, NJ), NDUFB8 (1:1,000; Abcam, Cambridge, MA), SDHB (1:1,000; Abcam), UQCRC2 (1:1,000; Abcam), COXIV (1:1,000; Abcam), ATP5A (1:1,000; Abcam), PGC-1α (1:1,000; Merck Millipore, Darmstadt, Germany), α-tublin (1:3,000; Santa Cruz Biotechnology, Shanghai, China), and β-actin (1:3,000; KANGCHEN, Shanghai, China).
Flow cytometry analysis of mitochondrial mass and mitochondrial membrane potential
Mitochondrial mass and membrane potential were analyzed by flow cytometry. Total mitochondrial content and actively respiring mitochondrial content were assessed by flow cytometry using MitoTracker Green (Invitrogen, Carlsbad, CA) staining (independent of mitochondrial inner membrane potential) and MitoTracker Deep Red (Invitrogen) staining (potential-dependent mitochondrial accumulation), respectively. MSCs incubated with culture medium were stained with MitoTracker Green (50 nM) or MitoTracker Deep Red (50 nM) for 30 min at 37°C (protected from light). For mitochondrial membrane potential measurement, MSCs were treated with 100 nM tetramethylrhodamine methyl ester (TMRM; Sigma-Aldrich) for 30 min at 37°C (protected from light). Thereafter, cells were collected after trypsinization and fluorescence was assessed by flow cytometry with a BD FACSCanto II Flow Cytometer (BD Biosciences, San Jose, CA).
Mitochondrial superoxide measurement
Mitochondrial superoxide was measured by flow cytometry following staining with MitoSOX™ Red mitochondrial superoxide indicator (Invitrogen). MSCs incubated with culture medium were stained with 5 μM MitoSOX reagent for 10 min at 37°C (protected from light). Thereafter, cells were collected after trypsinization and fluorescence was assessed by flow cytometry with a BD FACSCanto II Flow Cytometer (BD Biosciences, San Jose, CA).
mtDNA quantification
Genomic nuclear and mtDNA was extracted from MSCs using a Wizard® Genomic DNA Purification Kit (Promega, Madison, WI), according to the manufacturer's instructions. Quantitative polymerase chain reaction (qPCR) was used to measure the copy number of mtDNA encoding 16S rRNA, which was normalized to the copy number of nuclear DNA encoding β2-microglobulin. The reaction mix of DNA, primers, and SYBR Green was run under the conditions as previously described [30], using the ABI 7500 Fast Real-time PCR system (Applied Biosystems, Grand Island, NY). The primers used for 16S rRNA amplification were as follows: 5′-GCCTTCCCCCGTAAATGATA-3′ (forward), 5′-TTATGCGATTACCGGGCTCT-3′ (reverse). For β2-microglobulin, primers were: 5′-TGCTGTCTCCATGTTTGATGTATCT-3′ (forward), 5′-TCTCTGCTCCCCACCTCTAAGT-3′ (reverse).
Measurement of MSCs oxygen consumption rate
Mitochondrial respiration in intact cells was measured by using a high-resolution respirometry system (Oxygraph-2k; OROBOROS Instruments, Innsbruck, Austria) as described previously [31]. In brief, 5 × 105 cells were suspended in MIR05 (respiration medium containing 0.5 mM ethylene glycol tetraacetic acid, 3 mM MgCl2 · 6H2O, 60 mM potassium lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose, and 1 g/L fatty acid-free bovine serum albumin, pH 7.1) and added to the chamber. The basal oxygen consumption rate (OCR) was recorded in assay medium when the respiration was stabilized. Thereafter, cells were exposed to sequential injections of oligomycin (1 μM, ATP synthase complex V inhibitor), carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (2 μM, collapses the proton gradient, disrupts the mitochondrial membrane potential, and uncouples oxygen consumption from ATP production), and rotenone (1 μM, complex I inhibitor) plus antimycin A (1 μM, complex III inhibitor) with three consecutive OCR measurements taken after each injection. OCR measurements were expressed as pmol/min.
Green fluorescent protein lentivirus transfection
MSCs were genetically engineered to express green fluorescent protein (GFP) via lentivirus transfection before cell transplantation. In brief, MSCs were stably transduced [multiplicity of infection (MOI) = 50] with lentiviral vectors encoding GFP (Genechem, Shanghai, China). Lentivirus was added together with 8 μg/mL polybrene (Sigma-Aldrich) in culture medium for 12 h and then the medium was completely refreshed the next day. Transfection efficiency was evaluated by fluorescence microscopy.
Nonhuman primate MI model
Cynomolgus monkeys underwent MI surgeries according to previously described procedures [32,33]. Animals were allocated as three groups: DMEM, Vehicle-OMSCs, and SRT1720-OMSCs. Also, aged hMSCs were genetically engineered to express GFP at 3 days before cell delivery. After immunosuppression treatment, cynomolgus monkeys underwent a left thoracotomy and MI was induced with the left anterior descending coronary artery permanently ligated and verified by substantial ST-segment elevation from electrocardiography monitoring. Thirty minutes after the coronary artery ligation, animals in the Vehicle-OMSCs group received intramyocardial injection of 1 × 107 vehicle-treated aged hMSCs suspended in 1 mL DMEM, whereas animals in the SRT1720-OMSCs group received aged hMSCs that had been pretreated with 0.5 μM SRT1720 for 24 h. Also, animals in the DMEM group were injected with the same volume of DMEM, which served as a cell-free placebo. Five intramyocardial injections were performed for MSCs administration (five injections, of 2 × 106 cells in 0.2 mL sterile DMEM) to the peri-infarct regions with a 29-gauge syringe. A purse-string suture was placed around each injection site to prevent the leakage of injected cells. The chest was closed following a standard operative procedure, and monkeys were monitored carefully until full recovery.
Immunosuppression
All cynomolgus monkeys, cell treated and DMEM treated, received cyclosporine A and methylprednisolone as an immunosuppressive therapy to prevent rejection of human stem cell transplants. Five days before cell/DMEM delivery, cynomolgus monkeys were given cyclosporine A orally at a dose of 45 mg/kg/day for the experimental duration and the plasma concentration was maintained at a level of 100–250 ng/mL. Methylprednisolone was administered starting from 1 day before cell/DMEM injection in all animals with a loading-dose of 500 mg per animal, which was then continued with a dose of 1 mg/kg daily until the end of the study.
Immunofluorescence staining
Cynomolgus monkeys were euthanized at 3 days after MI surgery and heart tissues were collected and prepared as described previously [32]. Engraftment of GFP-positive hMSCs in monkey hearts was assessed by immunofluorescence staining. Tissue sections were frozen and processed for immunofluorescence staining. Slides were fixed in 4% formalin for 10 min, permeabilized with 0.2% triton-X100 for 20 min and then blocked with 3% bovine serum for 1 h and incubated with the anti-GFP antibody (1:200; Abcam) overnight at 4°C. Thereafter, the slides were incubated with the corresponding secondary antibodies for 1 h at room temperature and mounted in VECTASHIELD with 4′-6-diamidino-2-phenylindole Mounting Medium (Vector Laboratories, Burlingame, CA). Photographs were taken at a magnification of 200 × , and five random high-power fields from the border zone of each heart sample were analyzed.
Quantitative analysis of cell retention
MSCs engraftment levels in the heart were estimated by quantifying GFP DNA levels via qPCR as described previously [34]. Heart apex samples were collected and submitted to DNA extraction with a Wizard Genomic DNA Purification Kit (Promega). The GFP DNA gene sequence was amplified by qPCR using SYBR Premix Ex Taq (Takara Biotechnology Co., Dalian, China). The primers used were as follows: 5′-GCGAGAAGATCATCCCCGTG-3′ (forward), 5′-ACTTCTGGTTCTTGGCGTCG-3′ (reverse). The engraftment of GFP-positive hMSCs in the recipient monkeys was calculated against a standard curve obtained by mixing monkey DNA prepared from normal hearts with DNA isolated from a known number of hMSCs (diluting the genomic DNA of GFP-positive MSCs in normal monkey heart tissue genomic DNA at a ratio from 1:1 to 1:500). Cell engraftment was quantified as the number of GFP-positive cells per 100 heart cells in apex region.
Cardiac magnetic resonance imaging acquisition and analysis
Cardiac magnetic resonance imaging (CMRI) was conducted on a 1.5 T clinical scanner (Siemens Medical Systems, Erlangen, Germany) with a Syngo magnetic resonance software. Animals underwent CMRI at 3 days and 3 months after MI induction and treatment. After intramuscular injection of the ketamine (10 mg/kg) and xylazine (1 mg/kg) anesthetic mixture, the animal was placed in a supine position within the scanner and a four-element phased array coil was placed around the animal's chest. The CMRI protocol included cine imaging for assessment of the LV geometry, global LV contractile function, regional systolic LV wall thickening, and delayed gadolinium-enhanced imaging for assessment of the infarct size. The CMRI sequence parameters used were slice thickness: 8 mm, gap thickness: 1.2 mm, repetition time: 29.43 ms, flip angle: 50°, and field of view: 180 mm × 147 mm. All CMRI scans were analyzed by a reviewer who was blinded as to treatment allocation and timing of the evaluations. The scar size was calculated from the short-axis delayed enhancement images covering from the apex to the mitral valve plane.
Statistical analysis
All data are expressed as mean ± standard error of the mean. The differences between two independent experimental groups were determined using an unpaired two-tailed Student's t test, while differences between four groups were compared using one-way analysis of variance followed by Tukey's post hoc test. Kruskal-Wallis with Dunn's multiple comparisons test was performed for in vivo cardiac function analysis. Also, within-group comparisons of changes from baseline to the study endpoint were carried out using a paired Student's t test. All statistical analyses were performed using the GraphPad Prism software, and the threshold for significance level was set at P < 0.05.
Results
Isolation and characterization of aged hMSCs
Aged hMSCs were isolated and characterized by cell surface antigen expression analysis with flow cytometry. It turned out that aged hMSCs expressed typical MSC markers, including CD29, CD44, CD90, and CD105, but no hematopoietic lineage markers such as CD34 and CD45 (Supplementary Fig. S1A). Also, these aged hMSCs remained their pluripotency, which was confirmed by their capacity of osteogenic, adipogenic, and chondrogenic differentiation in vitro (Supplementary Fig. S1B).
SRT1720 protects aged hMSCs against H2O2-induced apoptosis by inhibiting the mitochondria-mediated pathway
To verify the antiapoptosis effects of SRT1720, aged hMSCs were pretreated with SRT1720 and then exposed to H2O2 as described in our earlier article [26], and cell apoptosis rate was measured by annexin V and PI double-labeling staining. While SRT1720 had no effect on apoptosis under normal culture condition, SRT1720-OMSCs showed decreased percentage of early apoptotic cells (annexin V positive and PI negative) as well as late apoptotic cells (annexin V positive and PI positive) following H2O2 exposure, compared with the Vehicle-OMSCs group (Fig. 1A). To investigate the involvement of mitochondria in the antiapoptotic effects of SRT1720, we examined caspase-9 activation and release of cytochrome c from mitochondria into the cytosol. Caspase-9 is the initiator proteolytic enzyme activated during mitochondria-mediated apoptotic events, and we found that SRT1720-OMSCs had significantly lower cleaved caspase-9 expression compared with Vehicle-OMSCs following H2O2 exposure, which was consistent with attenuated caspase-3 activation (Fig. 1B–D). Furthermore, the amount of cytochrome c was markedly decreased in the cytosolic fraction of SRT1720-OMSCs compared with that of Vehicle-OMSCs under H2O2 treatment (Fig. 1E, F). These results demonstrate that improved mitochondrial integrity is involved in the protective effects of SRT1720 on SRT1720-OMSCs against H2O2-induced apoptosis.
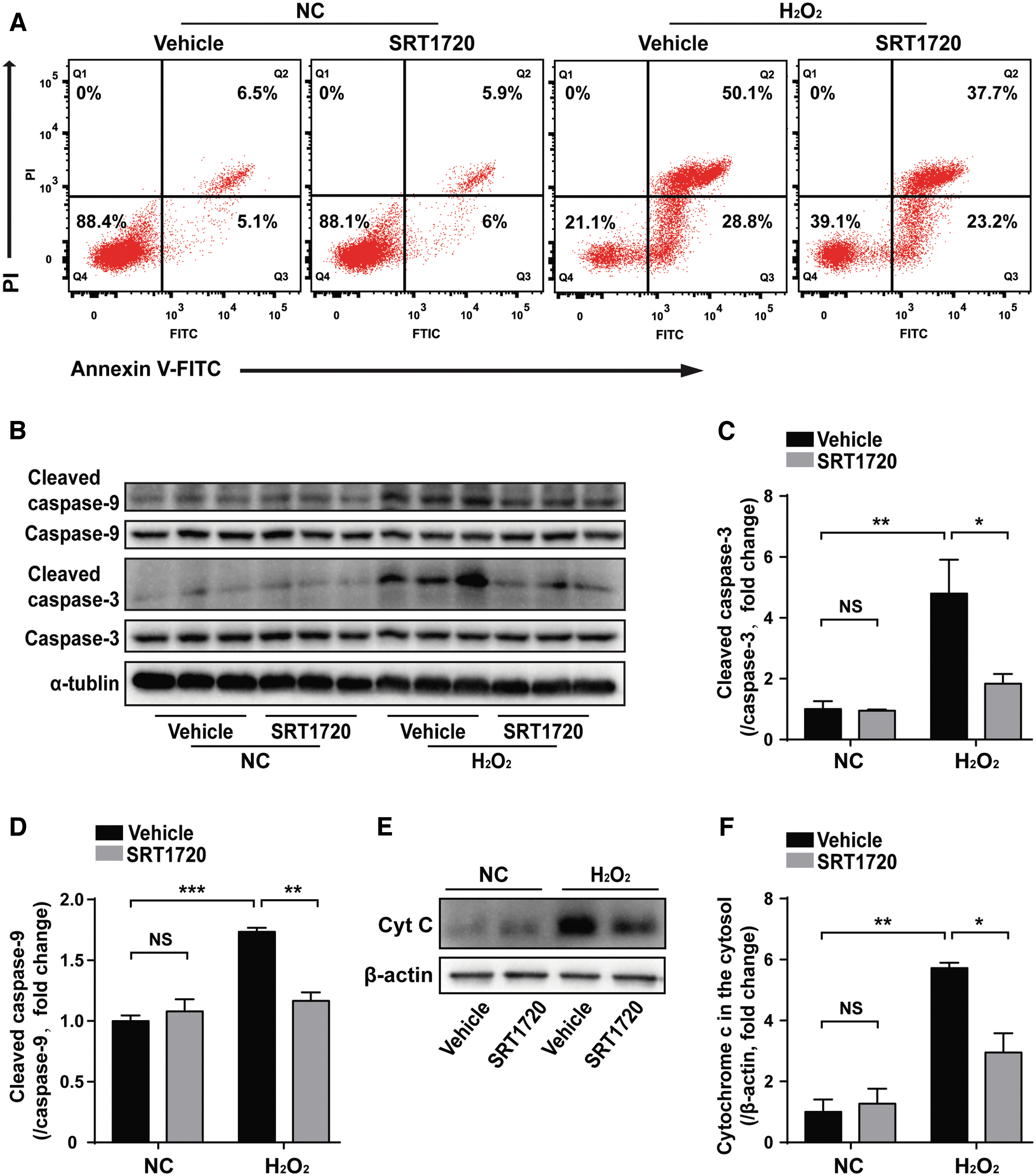
SRT1720 pretreatment attenuated aged hMSCs apoptosis following H2O2 exposure in vitro.
SRT1720 promotes mitochondrial biogenesis of aged hMSCs
There was evidence that mitochondrial content was reduced in hMSCs of aging individuals [35]. We evaluated mtDNA levels in young and aged hMSCs. Quantification of mtDNA copy numbers showed that aged hMSCs had decreased mtDNA content compared with young hMSCs (Fig. 2A). We further analyzed the protein expression of representative components of the mitochondrial electron transport chain (ETC), and we found that expression of NDUFB8, SDHB, and UQCRC2 was significantly reduced in aged hMSCs (Fig. 2B, C). In addition, the protein expression levels of PGC1α, the master regulator of mitochondrial biogenesis, were also decreased in aged hMSCs (Fig. 2D). These data suggest that age-related decrease in mitochondrial content is likely due to impaired mitochondrial biogenesis of aged hMSCs.

Mitochondrial content was decreased in aged hMSCs compared with young hMSCs.
While SIRT1 activation has been shown to increase mitochondrial content in multiple cell types [36,37], we tested if SRT1720 administration could improve mitochondrial biogenesis of aged hMSCs. We found that SRT1720 treatment resulted in increased intensity of MitoTracker green fluorescence in aged hMSCs, which reflected an increase in mitochondrial mass, compared with the vehicle group (Fig. 3A). Flow cytometry analysis of MitoTracker Deep Red staining, which identified intact and functional mitochondria, showed increased fluorescence intensity in the SRT1720 group similarly (Fig. 3B). The increase in mitochondrial mass in response to SRT1720 treatment was further confirmed by an increase in mtDNA content (Fig. 3C). The western blot analysis demonstrated that SRT1720 treatment elevated the protein expression levels of NDUFB8, SDHB, UQCRC2, COXIV, and ATP5A, components of the mitochondrial ETC in aged hMSCs (Fig. 3D, E). Taken together, these data show that SRT1720 treatment promotes mitochondrial biogenesis of aged hMSCs.

SRT1720 promoted mitochondrial biogenesis of aged hMSCs.
SRT1720 enhances mitochondrial function of aged hMSCs
To investigate whether mitochondrial function is affected by SRT1720 treatment, we measured OCR of aged hMSCs as indicators of mitochondrial respiration function (Fig. 4A). Metabolic profiling analysis demonstrated that aged hMSCs treated with SRT1720 had a modest increase in basal oxidative respiration (Fig. 4B), while treatment with the mitochondrial uncoupler FCCP resulted in a dramatically higher maximal respiration in SRT1720-treated cells compared with the vehicle treatment group (Fig. 4C). Also, SRT1720-treated aged hMSCs also displayed significantly increased TMRM fluorescence intensity compared with vehicle-treated cells, which indicated higher mitochondrial membrane potential levels (Fig. 4D). As mitochondria are the main source of reactive oxygen species (ROS) in mammalian cells, we evaluated the effects of SRT1720 on the mitochondrial ROS by flow cytometry following staining with MitoSOX Red mitochondrial superoxide indicator.
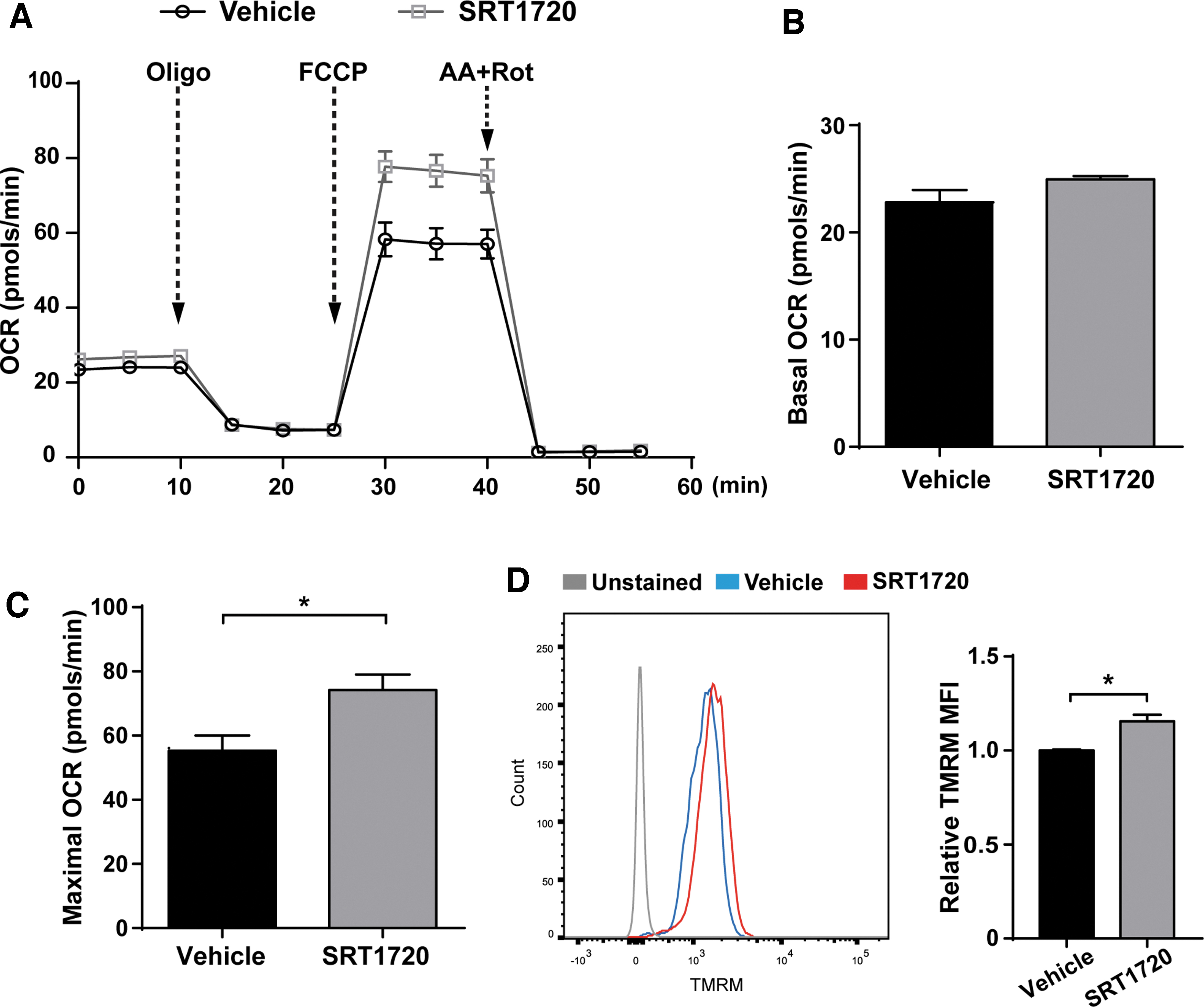
SRT1720 enhanced mitochondrial function of aged hMSCs.
Flow cytometric data showed that treatment with SRT1720 markedly decreased mitochondrial superoxide levels in aged hMSCs (Supplementary Fig. S2). These findings indicate that, in addition to promoting mitochondrial biogenesis, SRT1720 treatment also results in enhanced mitochondrial function in aged hMSCs.
SRT1720 pretreatment improves aged hMSCs engraftment in infarcted hearts of nonhuman primates
Next, we tested the effects of SRT1720 pretreatment on the cell engraftment in a nonhuman primate MI model. Six male cynomolgus monkeys underwent MI surgery and received Vehicle-OMSCs or SRT1720-OMSCs transplantation (n = 3 for each group). All monkeys were euthanized and heart tissues were obtained at 3 days after MI and representative macroscopies of short-axis cardiac slices of one vehicle-OMSCs and one SRT1720-OMSCs-treated cynomolgus monkey are shown in Supplementary Fig. S3A. As aged hMSCs were genetically engineered to express GFP before transplantation, cell engraftment was determined by immunofluorescence staining for GFP expression and quantifying GFP DNA levels via qPCR. Clusters of GFP-positive aged hMSCs were identified in the peri-infarct areas of heart tissues, and the number of cells retained was significantly increased in the SRT1720-OMSCs group (Fig. 5A, B). qPCR results showed that the engraftment of aged hMSCs was significantly improved in the SRT1720-OMSCs group compared with the Vehicle-OMSCs group at day 3 post-MI (Fig. 5C). These results suggest that SRT1720 pretreatment increases cell engraftment of aged hMSCs at day 3 in the infarcted hearts of cynomolgus monkeys.

SRT1720 pretreatment improved engraftment of aged hMSCs.
SRT1720-OMSCs transplantation improves left ventricular ejection fraction recovery post-MI
We next explored the possible effects of SRT1720-OMSCs transplantation on the recovery of cardiac function following MI in cynomolgus monkeys. A 3-month study was implemented to evaluate changes of cardiac function in nonhuman primates and animals were allocated as three groups: DMEM, Vehicle-OMSCs, and SRT1720-OMSCs (DMEM group: n = 4; Vehicle-OMSCs group: n = 4; SRT1720-OMSCs group: n = 4). To investigate cardiac function, we performed CMRI at 3 days (baseline) and 3 months after MI injury and cell transplantation. CMRI-derived cardiac function data are presented in Supplementary Table S1.
The left ventricular ejection fraction (LVEF) assessed by CMRI did not significantly differ between three groups at baseline. However, when comparing the evolution of LVEF within each group over time after 3 months, in both DMEM and Vehicle-OMSCs groups, the LVEF tended to decrease, whereas in the SRT1720-OMSCs group, the LVEF improved significantly (Fig. 6A). The strikingly different pattern of changes in LV function can be better delineated when the absolute change in LVEF was calculated between baseline and 3-month (Δ LVEF): with both DMEM and Vehicle-OMSCs groups showing a decrease of −3.37% ± 1.86% and −3.10% ± 2.43%, respectively, whereas the SRT1720-OMSCs group demonstrating an improved Δ LVEF (3.78% ± 0.46%; P < 0.05 vs. DMEM) (Fig. 6B). Measurements of cardiac chamber volume showed that left ventricular end-systolic volume were increased over time in the DMEM and Vehicle-OMSCs group, but did not differ significantly in the SRT1720-OMSCs group (Supplementary Table S1). Analysis of late gadolinium-enhanced images by CMRI revealed that infarct size did not differ significantly over time within each group (DMEM group: baseline post-MI, 4.10 cm2 ± 0.37 cm2 vs. 3 months post-MI, 3.67 cm2 ± 0.27 cm2, P = 0.311; Vehicle-OMSCs group: baseline post-MI, 4.68 cm2 ± 0.39 cm2 vs. 3 months post-MI, 3.99 cm2 ± 0.22 cm2, P = 0.158), however, a statistical trend of decrease in infarct size was observed in the SRT1720-OMSCs-treated hearts at 3 months (3.41 cm2 ± 0.56 cm2) compared with baseline right after MI (4.43 cm2 ± 0.31 cm2; P = 0.055) (Fig. 6C and Supplementary Fig. S3B). Thus, analysis of infarct size suggested an improved infarct shrinkage in the SRT1720-OMSCs group even though intergroup analysis showed no significant differences in absolute changes of infarct size between three treatment groups (Fig. 6D). These findings indicate that SRT1720-OMSCs transplantation improved global LV systolic function post-MI.

Assessment of cardiac function at 3 days and 3 months post-MI by cardiac magnetic resonance imaging.
Discussion
Increasing age is correlated with reductions in functional characteristics of MSCs, including proliferation, proangiogenic potency, as well as antiapoptotic capabilities [38 –40]. A previous study using MI models in rats found that aged hMSCs exhibited worse survival rate and conferred less functional improvement than young hMSCs [41]. In the present study, we demonstrate that pretreating aged hMSCs with SRT1720 enhances their resistance to H2O2-induced apoptosis via inhibiting the mitochondria-mediated pathway and augments engraftment substantially in the infarcted myocardium of cynomolgus monkeys.
Mitochondria serve as the power house of the cell, generating ATP and providing essential intermediate metabolites to sustain cell function. Recent research suggested that personal characters such as chronological age and disease conditions altered mitochondrial function and metabolic profile of hMSCs. For example, Pietila et al. found that hMSCs from aged donors exhibited dramatically lower mitochondrial-to-cytoplasm area and reduced manganese superoxide dismutase levels compared with cells derived from young donors [35]. Consistent with their findings, our results showed that aged hMSCs had decreased mitochondrial mass compared with young hMSCs as manifested by mtDNA content. In addition, the protein expression levels of PGC1α, the master regulator of mitochondrial biogenesis, were also decreased in aged hMSCs, implying that the decrease in the abundance of mitochondria might be partly due to diminished mitochondrial biogenesis. Furthermore, a recent study indicated hMSCs from subjects with atherosclerosis exhibited impaired mitochondrial function that contributed to higher intercellular ROS levels, compromised immunomodulatory capacity, and increased susceptibility to cell death compared to hMSCs from individuals without atherosclerosis [42]. Obesity also induces mitochondrial abnormalities and dysfunction, leading to elevated ROS production [43,44]. Taken together, these studies provide evidence that mitochondrial alterations due to donor age or morbidity possibly account for donor-related variability in cell quality and thus the mitochondrion is emerging as a plausible pharmacological target to improve the therapeutic efficacy of hMSCs.
Since its discovery as a conserved NAD+-dependent deacetylase, SIRT1 has received much attention on its role in regulating metabolic health [45]. Previous studies showed that SIRT1 upregulated mitochondrial biogenesis and enhanced oxidative metabolism in numerous tissues, including liver, skeletal muscle, and adipose tissue [19]. Also, recent efforts toward rejuvenating adult stem cells suggested the possibility of the restoration of aged stem cell function by improving mitochondrial function via supplementing NAD+ precursor. For example, treating aged mice with the NAD+ precursor nicotinamide riboside exerted a potent rejuvenating effect on muscle stem cells by enhancing mitochondrial function, which was mediated by SIRT1 protein activation [46]. However, to our knowledge, the role of SIRT1-associated nuclear/mitochondrial axis in aged hMSCs has not been investigated yet. Here, we found that SRT1720 pretreatment could protect aged hMSCs against mitochondria-mediated apoptotic cell death. SRT1720 treatment promotes mitochondrial biogenesis and mitochondrial function of aged hMSCs. These results are consistent with previous reports that pharmacological activation of mitochondrial biogenesis can block apoptotic cell death in other in vitro and in vivo models [47 –51].
Although MSCs have been widely used for cardiac repair, the vast majority of transplanted MSCs are lost due to apoptosis or cell death after grafting into injured hearts, significantly offsetting their long-term efficacy [52 –54]. The advanced aging issues inherent to the donors, on the contrary, diminishes the homing abilities of stem cells, further hampering their cardioreparative effects and restricting their clinical application. Up to date, many methods have been studied and developed to improve the fates of transplanted stem cells, such as hypoxia preconditioning, Bcl-2 and Akt gene overexpression [33,55,56]. Nevertheless, few of these studies have been focused on the capacity of aged stem cells to treat MI. Herein, we explored for the first time the potential effects of SRT1720 pretreatment using MSCs derived from elderly people in the treatment of MI in nonhuman primates. The present experiments showed that the retention of aged hMSCs in the infarcted myocardium of cynomolgus monkeys was significantly augmented by SRT1720 pretreatment. With regard to changes in cardiac function, our results demonstrated that SRT1720-OMSCs were superior to Vehicle-OMSCs in improving global LVEF at 3 months post-MI. Also, a borderline statistically significant decrease in infarct size over time was observed in the SRT1720-OMSCs group. Thus, SRT1720 pretreatment represents as a novel strategy for the optimization of aged hMSCs.
There are several limitations to this study. First, our experimental settings cannot fully mimic the clinical scenario. To avoid potential xenogeneic rejection risk of hMSCs, we used immunosuppressive treatment regimens on cynomolgus monkeys, while it was unnecessary for patients choosing autologous stem cell therapy. Second, because it is a primate study, the group size is smaller than is typical for a rodent study. Although we reported statistically significant data for LV systolic function improvement, some parameters of cardiac function showed trends which would require larger groups to attain statistical significance.
Conclusions
In conclusion, for the first time, we reported that SRT1720 pretreatment enhanced engraftment of aged hMSCs in a nonhuman primate MI model. It was related with increased mitochondrial biogenesis and function following SRT1720 treatment. Our findings pointed to the prosurvival effects of SRT1720 and raised the possibility of regulating aged hMSCs function via modulating mitochondrial functions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key R&D Program of China (2019YFA0110400, 2016YFC1301204 for Jian'an Wang and 2017YFA0103700 for Xinyang Hu), the grants from National Natural Science Foundation of China (No. 81870292 for Jian'an Wang, 81622006, 81670261 for Xinyang Hu, 81570233, 81770252 for Xianbao Liu), The Key R&D projects of Zhejiang Province (No. 2015C03028 for Jian'an Wang, 2018C03014 for Xinyang Hu), the Fundamental Research Funds for the Central Universities (2019XZZX005-4-03 for Zhu Wei, 2020FZZX003-02-11 for Rongrong Wu).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.