
Abstract
Periodontal tissues consist of cementum, periodontal ligaments, and alveolar bone, which provide indispensable support for physiological activities involving mastication, swallowing, and pronunciation. The formation of periodontal tissues requires a complex process, during which a close relationship with biomineralization is noticeable. Alveolar bone and cementum are physically hard, both of which are generated from biomineralization and possess the exact mechanical properties resembling other hard tissues. However, when periodontitis, congenital abnormalities, periapical diseases, and other pathological conditions affect the organism, the most common symptom, alveolar bone defect, is always unavoidable, which results in difficulties for current clinical treatment. Thus, exploring effective therapies to improve the prognosis is important. Matrix vesicles (MVs), a special subtype of extracellular vesicles related to histogenesis, are widely produced by the stem cells of developing hard tissues. With the assistance of the enzymes and transporters contained within them, MVs can construct the extracellular matrix and an adequate microenvironment, thus promoting biomineralization and periodontal development. Presently, MVs can be effectively extracted and delivered by scaffolds and generate hard tissues in vitro and in vivo, which are expected to be translated into therapies for alveolar bone defects. In this review, we generalize recent research progress on MV morphology, molecular composition, biological mechanism, and, in particular, the biological functions in periodontal development. In addition to the above unique roles of MVs, we further describe the available MV-related biotechnologies and achievements that make them promising for coping with existing problems and improving the treatment of alveolar bone defects.
Introduction
Periodontal tissues that widely surround the tooth are endowed with intricate structures, special biological features, and physiological significance for humans and rodent animals. The cementum, periodontal ligaments, and alveolar bone are hierarchically arrayed in three layers that manifest as a sandwich-like structure, which together comprise the periodontal tissues [1]. Among them, cementum and alveolar bone have similar mechanical strength and physical properties to those of other hard tissues, and alveolar bone is universally considered to have paramount physiological importance [2 –4]. Alveolar bone can help the tooth resist external forces to maintain stability in mastication, pronunciation, and other movements. In addition, it provides nerve and blood supply to the tooth, securing the physiological health and homeostasis of tooth development [2,4,5] (Fig. 1).
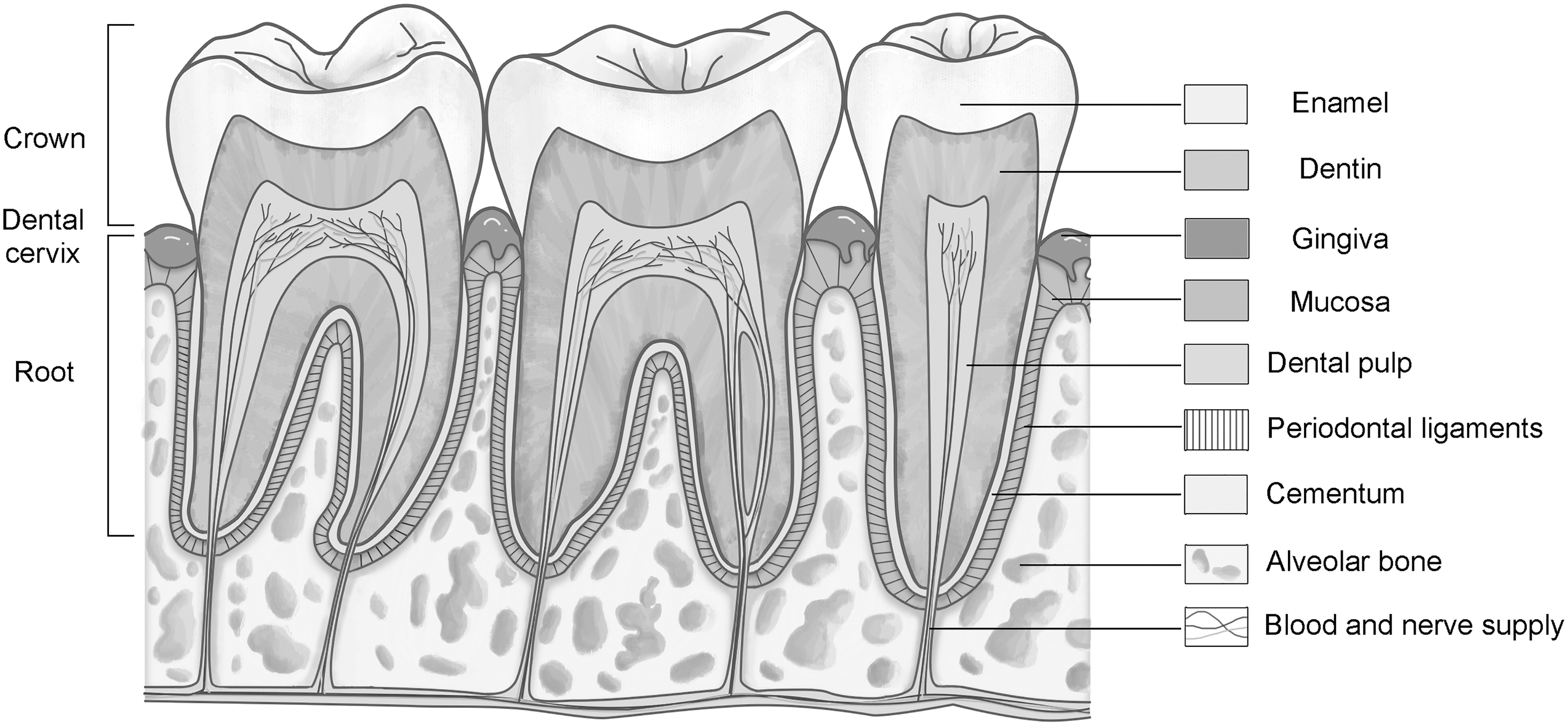
Schematic diagram of teeth and periodontal tissues. This figure presents the anatomy of the right mandibular secondary premolar, first molar, secondary molar, and neighboring periodontal tissues from a longitudinal profile. The dental cervix divides the tooth into two parts: crown and root, which comprise enamel, dentin, and dental pulp. The surrounding structures, such as the gingiva, mucosa, and periodontal tissues (cementum, periodontal ligaments, and alveolar bone) are situated around the tooth. In addition, the affiliated capillaries and nerve endings are dispersed within the alveolar bone, which provide blood and nerve supply for the teeth.
However, periodontitis, periapical periodontitis, cysts, and some congenital abnormality diseases tend to invade alveolar bone, resulting in defects and tooth loss, which are intricate problems for clinical treatment (Fig. 2) [6 –8]. The current therapies for alveolar bone defects mainly include guided tissue regeneration (GTR), autologous bone grafting, and heterogeneous bone filling. These techniques have shown uncertain therapeutic effects and some limitations, and they have not yielded satisfying curative outcomes in many cases. This is because these therapies probably cannot activate intercellular communication, which is imperative to promote tissue regeneration [6,8,9].
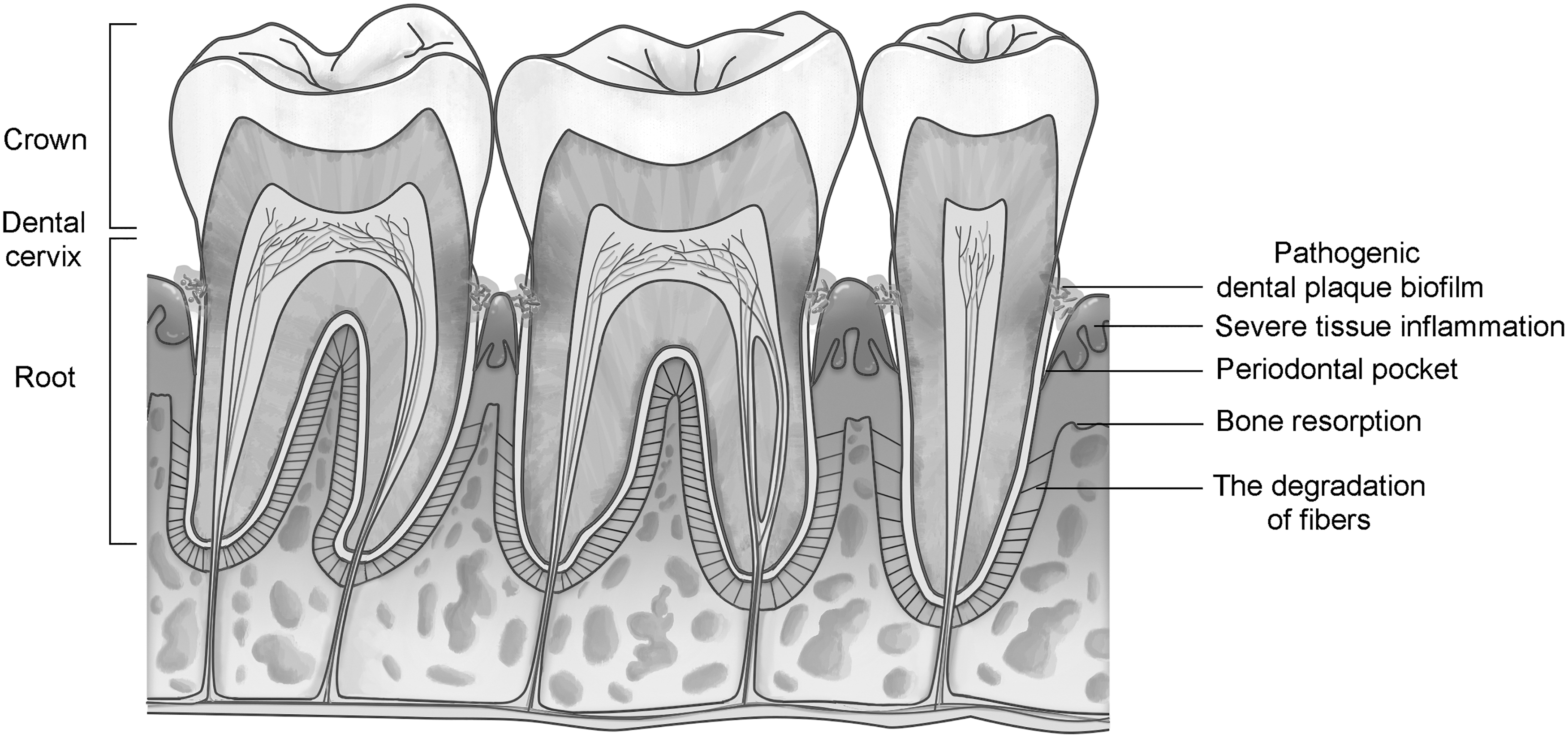
Schematic diagram of chronic periodontitis. This diagram shows the main pathological findings of chronic periodontitis. Pathogenic dental biofilms assemble on the surface of the tooth root and dental cervix, triggering severe inflammation of the gingiva and mucosa. The exaggerated inflammatory response results in the degradation of ligament fibers, bone resorption, and ultimately alveolar bone defects.
Recently, many studies have demonstrated that extracellular vesicles (EVs), acting as a major mediator in intercellular communication, can transfer information between cells and stimulate the self-healing of injured tissues [10,11]. EVs always refer to vesicles (ranging 30–5,000 nm in diameter) that possess enclosed vesicular structures and are distributed throughout the extracellular matrix (ECM) after being secreted by cells [10]. Regarded as important mediators, EVs have been successfully applied in medical diagnosis and treatment as biomarkers, drug carriers, or tissue regenerative promoters [12], and they play an increasingly prominent role in hard tissue formation [11,13,14]. Some biologists have revealed the advantages of orthopedic treatment with the involvement of EVs [15].
In light of their biological differences, EVs are divided into several subtypes, including exosomes, shedding microvesicles, and apoptotic bodies (ApoBDs) [10]. Remarkably, one special subtype, matrix vesicles (MVs), is closely associated with hard tissue formation. MVs (30–500 nm in diameter) are defined as EVs produced by stem cells in developing hard tissues (eg, bone, cartilage, dentin, cementum, and alveolar bone) and adhere to the ECM through chemical bonding or physical traction [16]. MVs are distinctly different from other EVs since they contain diverse enzymes and transporters, and they can build a microenvironment for mineralization and constitute the matrix of hard tissues through the continual uptake of inorganic electrolytes [17 –20].
The discovery of MVs can be traced back to 1967. After injecting human amniotic cells into rat muscles to induce chondrogenesis, Anderson's team observed some vesicle-like clusters under electronic microscopy, which encapsulated needle-shaped minerals and subsequently grew mineralized nodules. Since these vesicles were stably anchored in the matrix, they were naturally termed MVs [21,22]. Henceforth, various interesting findings have emerged in the past decades and have led to a breakthrough in our understanding of hard tissue development.
The existing studies mainly focus on exploring MVs from the aspect of morphology or biological processes and have proven that MVs are imperative for almost all hard tissue formation in the human body. During the physiological development of bone, cartilage, teeth, cementum, and alveolar bone, numerous MVs are produced to form mineralized nodules and constitute the matrix of hard tissues, simultaneously constructing the microenvironment for mineralization to accommodate cell osteogenic or odontogenic differentiation [23 –25].
In the past few years, EV-centered tissue regenerative technologies have become a promising approach for reconstructing and restoring damaged tissues, including alveolar bone defects [26,27]. Nevertheless, it is still difficult to completely regenerate the damaged alveolar bone by conventional EVs, probably because they lack the mineralization activity, connective stability, and functional specificity to induce hard tissue formation [14,28].
Compared to other EVs, MVs deserve more attention and should be considered a special subtype of EVs that can overcome these difficulties. These intrinsic vesicles from hard tissues contain abundant enzymes and can tightly adhere to the ECM to promote tissue remodeling and biomineralization, which may render them superior in tissue regeneration [19,25]. Furthermore, proteomic studies have revealed that MVs contain various functional proteins and growth factors, resulting in the stimulation of cell proliferation, construction of hard tissues, and suppression of tissue inflammation [29 –31]. The latest studies have reported that MVs prepared by chemical bionic technology displayed outstanding bone regeneration performance in calvarial defect models [32]. Under the microenvironment constructed by MVs, the damaged tissue upregulated the expression of proteins associated with hard tissue formation, and new bones were generally formed to fill the defect. Although naturally derived MVs have not been directly used in alveolar bone regeneration, the advanced MV extraction technologies and scaffold materials with predominant properties will make MV-centered tissue regenerative technologies a preferred option in the regeneration of alveolar bone in future studies, which is expected to provide a new reliable clinical therapy.
Herein, we provide an overview of the recent research progress on MVs from the aspects of morphology, molecular composition, and biological mechanism and highlight their biological functions in periodontal formation. Moreover, we summarize the present MV-related biotechnologies and conclude that MVs can act as a new promising curative approach in scientific research and clinical therapies for refractory alveolar bone defects.
MV Structure and Composition
MVs manifest a typical three-dimensional vesicular structure organized with three relatively individual portions: from the exterior to the interior, a vesicle membrane, membrane-binding proteins, and luminal contents.
Vesicle membrane
The vesicle membrane usually has a similar specificity to other biomembranes, as it is composed of a phospholipid bilayer that is rich in phosphatidylserine (PS), phosphatidylinositol, sphingolipids, and other lipids. As has been widely demonstrated, the vesicle membrane of MVs practically guarantees the stability and tightness of the vesicle and simultaneously provides attachment sites for membrane-binding proteins [17,33,34].
Membrane-binding proteins
This category includes a large variety of proteins with different functions and mainly consists of EV markers, enzymes, and cytoskeletal proteins that settle on the vesicle membrane [35]. According to many proteomic studies, most EV markers (such as CD9, CD81, heat shock protein [HSP], Rab, SNAREs, and annexins [Anx]) are expressed on the MV membrane [36] and participate in each stage of events during MV formation, such as the process of cargo assembly and vesicle release. The existence of EV markers confirms that MVs belong to EVs from the aspect of biological composition biogenesis [17,37]. Notably, enzymes and ion channel proteins are also attached to the membrane and fundamentally ensure the biological activity of MVs in biomineralization by initiating multiple enzymatic reactions. They principally include tissue nonspecific alkaline phosphatase (TNAP), ecto-nucleotide pyrophosphatase/phosphodiesterase 1 (ENPP1), ankylosis protein (ANK), sodium/phosphate cotransporter type III (Pit1), and calcium channel proteins (AnxA2 and AnxA6) [25,38,39]. Moreover, the proper execution of biological functions is closely related to complex physical movements. Cytoskeletal proteins, such as actin, fibronectin, and tubulin, are other key MV membrane proteins. From the biogenesis to the maturity of MVs, cytoskeletal proteins function by maintaining and supporting the multiple movements of MVs, including contraction, torsion, and traction [40]. Furthermore, the presence of some signaling proteins, regulatory proteins, and transcription-related proteins further increases the diversity and coordination of the molecular composition of the membrane [35].
Luminal contents
Distinctively, luminal contents encapsulated in MVs, with only a finite group of free proteins present, are mainly inorganic electrolytes and crystals, which show an obvious compositional heterogeneity compared with other vesicular components. As Sauer's chemical analysis revealed, Ca2+ and phosphate (PO4 3−, HPO4 2−, and H2PO4 −) are the main inorganic electrolytes. When a certain density is reached, the electrolytes will precipitate to form crystals in the luminal zone [41,42]. Thereafter, through diffraction spectrum tests, scholars have found more precisely that the crystals inside of immature MVs are composed primarily of amorphous calcium phosphate (ACP), as well as a small amount of octacalcium phosphates (OCP). However, with the consecutive accumulation of inorganic electrolytes and the maturation of MVs, the crystals are eventually reconstructed into ordered hydroxyapatite (HAP) [43,44]. Some free proteins, such as phosphoethanolamine/phosphocholine phosphatase-1 (PHOSPHO1), without binding to the membrane, actually promote the intake of inorganic electrolytes in MVs by transforming the electrolyte state [45].
In summary, in the MV structure, the vesicle membrane rich in lipids provides proper adhesion sites for a series of functional proteins and synchronously encompasses the contents that include calcium phosphorus and electrolytes. Accordingly, highly organized vesicles have been constructed in three-dimensional space, and the morphological features of MVs confirm that they are a specific EV subtype.
Nevertheless, it is still difficult to distinguish MVs from other EVs by current morphological or proteomic tests. For instance, MVs and exosomes share a large degree of similarity in vesicular structures and markers, which results in indistinguishable differences in the results of electron microscopy observations, particle diameter analysis, flow cytometry, Western blotting, and other tests [37]. Admittedly, there are obvious differences in content composition. Exosomes contain abundant mRNA and miRNA, while MVs contain numerous inorganic electrolytes and crystals, but are nearly lacking in nucleic acids [35,46]. Therefore, the application of energy spectral diffraction or exploitation of more effective content-based tests will help to discern MVs from other EVs in future studies.
Biological Process of MVs
Similar to other bioactive molecules, MVs undergo a complex biological process, which is tightly associated with hard tissue development. The first stage is biogenesis. When stimulated by signals of biomineralization, stem cells in hard tissues start to secrete substantial MVs through budding-cleavage or possibly the endocytosis-exocytosis pathway. Next, the produced MVs begin to perform biological functions automatically and initiate biomineralization. Generally, biomineralization involves two phases: MV-mediated mineralization and collagen mineralization. MV-mediated mineralization manifests as electrolyte internalization and mineral accumulation within the contents of MVs. The expansion of interior contents results in the rupture of the vesicle membrane of MVs, and MVs evolve into mineralized nodules. Then, biomineralization is followed by the phase of collagen mineralization. Driven by the collagen fibrils and the microenvironment for mineralization, mineralized nodules can become enlarged and enriched and eventually constitute the matrix components of hard tissues.
Biogenesis
Theoretically, each specific subtype of EVs takes a specific pathway of formation. The biogenesis of exosomes requires cell endocytosis and exocytosis, during which exogenous molecules are absorbed intracellularly to form endosomes and then released after integrating with the membrane. Shedding microvesicles are produced by budding and cleavage of the regional plasma membrane without cell endocytosis. ApoBDs, consisting of cell debris and degenerative organelles, are produced during cell apoptosis [47].
Regarding MVs, budding-cleavage is a classic pathway and is commonly accepted [14,48,49], which is universal in most cases and initiated from special domains of the cell membrane [48,50,51]. Triggered by signals of biomineralization, the cells hydrolyze substantial adenosine triphosphate (ATP) to acquire sufficient energy, which drives the redistribution of intracellular Ca2+ and inorganic molecules that have congregated in specific domains. Selective redistribution of Ca2+ further activates aminophospholipid translocases (flippases and floppases), scramblases, and calpain, causing physical bending of the plasma membrane, which appears as outward budding [10,46]. Then, these lipids, enzymes, proteins, and other molecules that will comprise the MVs gradually assemble in the curved region through the affinity of some targets, such as lipid rafts, and the anchoring force of other membrane molecules (for example, arrestin domain-containing protein-1 [ARRDC1] recruits endosomal sorting complex for transport [ESCRT] family proteins, which in turn regulate the distribution and delivery of intracellular substances [52]), inducing the evolution of physical bending into membrane protrusions. Meanwhile, mitochondria may transfer some necessary mineral precursors into the domains through the force of membrane potential variation [53]. As more molecules accumulate, the curvature of the cell protrusion is intensively increased, which results in domain cleavage and the formation of enclosed vesicles, namely, MVs [54]. Recent studies have proven that a small guanosine triphosphate hydrolase (GTPase), adenosine diphosphate (ADP)-ribosylation factor 6 (ARF6), plays an important regulatory role in this pathway. ARF6 can activate phospholipase D (PLD) and extracellular signal-regulated kinase (ERK) in succession through chemical combination and improve the activity of myosin light chain kinase (MLCK) at the budding site [55]. Accordingly, the movement of actin, myosin, microvilli, and other structures in this region can be significantly enhanced, thus promoting the formation of MVs.
Moreover, the endocytosis-exocytosis process may be an underlying pathway as some scholars recently disclosed [17,37,56] based on a series of phenomena and proteomic evidence. Originally, Yang et al. discovered two types of MVs in the bone biomineralization of zebrafish, which were shaped like multivesicular bodies (MVBs) and monovesicles [57], and this phenomenon was even observed in mammals such as mice [49]. In addition, extracellular minerals, such as iron, can be transported into cells and participate in MV construction with the cooperation of endosomes [53]. Since MVBs and endosomes are regarded as necessary for the endocytosis process, the formation of MVs may be more complicated than ordinarily imagined. Considering the many compositional similarities between exosomes and MVs, a possible endocytosis-exocytosis pathway for MV biogenesis was eventually inferred by Shapiro et al. [17].
The underlying endocytosis-exocytosis process is similar to that which produces exosomes [17,37,56]. On the membrane of those cells that are going to secrete MVs, highly active domains exist, which mediate the cellular physiological processes of endocytosis and metabolism. When endocytosis is initiated by exogenous molecular stimuli, ESCRT complexes located on these domains begin to recognize and transport the molecules, synchronously activating GTPase family proteins, including RAB11 and RAB35 [37]. GTPase complexes then degrade the domains and stimulate membrane invagination. As a consequence, early endosome vesicles are formed intracellularly and act as carriers for those molecules [58,59]. Subsequently, the Golgi complex modifies early endosomes by synthesizing TNAP, ENPP1, and other associated proteins and then transports these proteins into endosomes in small vesicle form, which ensures the constituent integrity of endosomes destined to become MVs. Then, under the regulation of several signaling molecular families, such as ESCRT, lipids, and transmembrane proteins, some special membrane domains of the early endosomes gradually curl up and encapsulate a portion of the contents to form intraluminal vesicles (ILVs), namely, inward budding [10,46]. Notably, when these ILVs are produced to some extent, the early endosomes mature and transform into multivesicular endosomes (MVEs) with multiple vesicles intraluminally [46,58]. However, a large proportion of MVEs will be targeted to lysosomes and undergo a degradation course due to their incomplete biological composition; only others with constituent integrity will fuse with the plasma membrane and release ILVs to the ECM to become MVs [60]. During the release process, Rab family proteins (including Rab27a, Rab27b, Rab11, and Rab35) are key regulators. They can directly connect the plasma membrane with MVEs, target MVEs to the destination, and ultimately promote MV release by increasing the permeability of the plasma membrane [61,62] (Fig. 3).

Biogenesis of MVs in osteoblasts. The classic budding-cleavage pathway and the underlying endocytosis-exocytosis process are involved. The right side of the cell membrane shows the budding process. Initially, diverse intracellular cargoes are enriched in special domains of the cell membrane and are probably modified by mitochondria. The expanded cargoes lead to the protrusion and subsequent cleavage of the domains, which form enclosed vesicles and MVs outside the cell. The budding-cleavage process mainly relies on ARF6 and cytoskeletal proteins. The potential endocytosis-exocytosis route starts from the upper left side of this diagram. Extracellular cargoes are sorted into receptor-ligand complexes and dissociative molecules by the cell membrane. Then, they are internalized from the cell surface to become early endosomes, relying on some regulators (such as Rab family proteins). With the enrichment of diverse cargoes, early endosomes evolve into MVEs. This course is assisted by several mediators (ESCRT, lipids, and tetraspanins) and the modification of the Golgi complex (by adding some endogenous molecules). Those individual vesicles budding inside MVEs are termed ILVs. Subsequently, some ILVs are degraded by hydrolases in the lysosome, while the others are targeted to integrate with the cell membrane and form MVs, which finally reside extracellularly to take effect. ESCRT, endosomal sorting complex for transport; ILV, intraluminal vesicle; MV, matrix vesicle; MVE, multivesicular endosome.
MV-mediated mineralization
Once produced, MVs are prone to adhere to the collagen fibers and proteoglycans in the ECM to obtain adequate stability, by which they can absorb inorganic electrolytes and mediate biomineralization [20]. This process mainly involves the transformation and transportation of phosphate and Ca2+ and relies on the various enzymes and ion channels within MVs (Fig. 4).
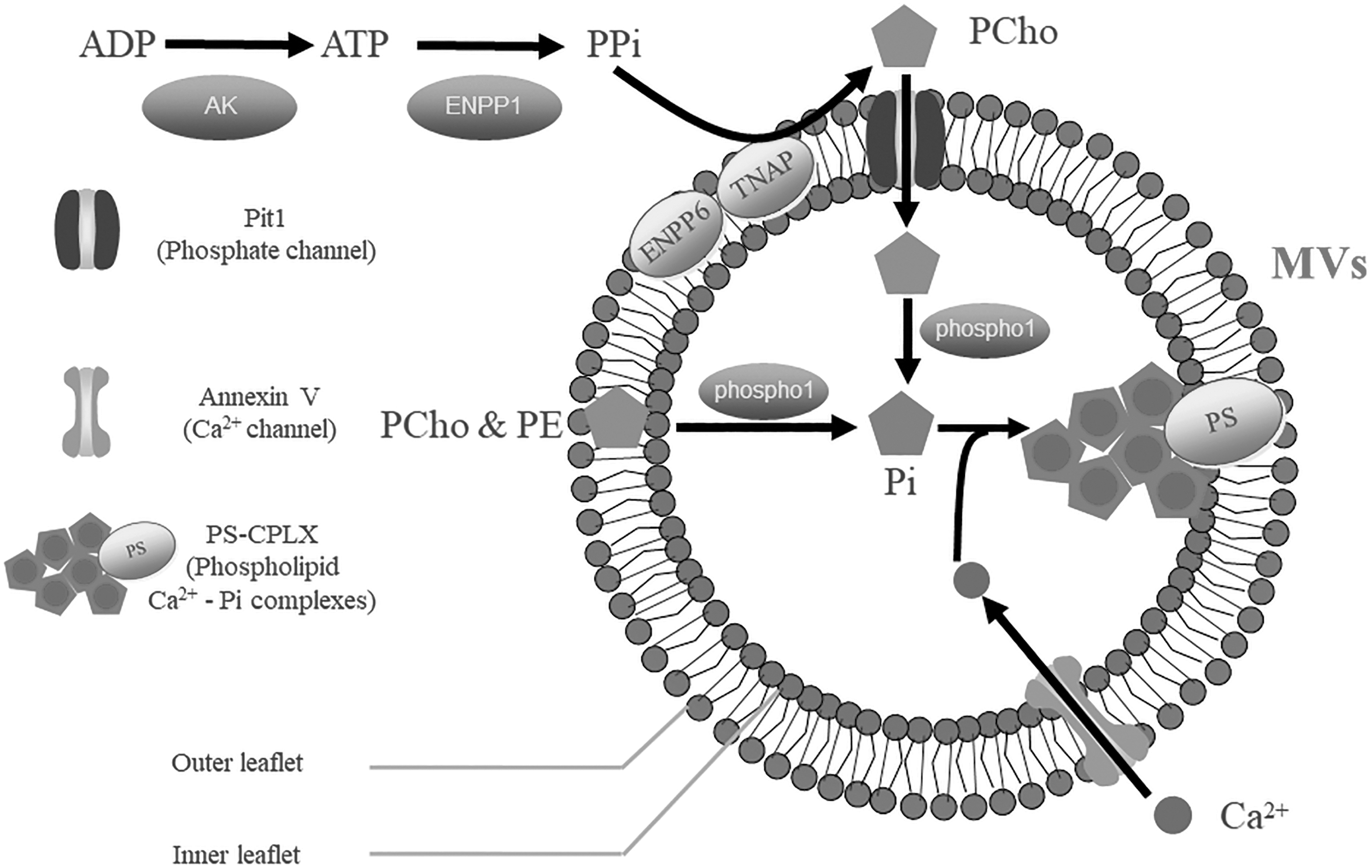
Schematic representation of MV-mediated mineralization. The vesicle membrane of MVs is composed of a phospholipid bilayer, which accommodates multiple enzymes, membrane transporters, and other molecules associated with biomineralization. This process is mainly characterized by the influx of phosphate and Ca2+. Initially, outside the vesicle, ADP is used to synthesize ATP by activated AK. Then, ENPP1 hydrolyzes ATP and produces PPi, a major and original source of exogenous phosphate. Vesicular membrane proteins (ENPP6 and TNAP) can recognize PPi and convert it into exogenous PCho, which later passes through the phosphate channel, Pit1, and is delivered into MVs. Subsequently, the exogenous PCho and the endogenous organic phosphate (including PCho or PE from the vesicle membrane) are hydrolyzed by phospho1 and then form Pi, the active state of phosphate. Meanwhile, external Ca2+ is delivered into MVs with the assistance of annexin V. Ultimately, the cumulative Pi, Ca2+, and PS that are located on the inner leaflet of the vesicle membrane bind chemically to form PS—CPLX, a group of minerals in the early phase of biomineralization. ENPP6, ecto-nucleotide pyrophosphatase/phosphodiesterase 6; PCho, phosphocholine; PE, phosphoethanolamine; Pi, inorganic phosphate; PPi, inorganic pyrophosphate; PS, phosphatidylserine; TNAP, tissue nonspecific alkaline phosphatase.
According to many previous studies, phosphate is affiliated with complex formation and transportation processes. Under stimuli from the microenvironment, extracellular adenylate kinase (AK) is activated to react with ADP and then synthesizes ATP, the major energy source for biological activities [63]. Then, ATP begins to release chemical energy within a hydrolysis process induced by ENPP1. During this period, inorganic pyrophosphate (PPi), a major source of exogenous phosphate, also forms [64]. TNAP located on the MV membrane will later recognize and convert PPi into phosphocholine (PCho), and PCho can effectively flow into MVs through the Pit1 channel [25,65]. Furthermore, the abundant membrane lipids have also been proven to be another essential source of inorganic phosphate (Pi), known as the endogenous source. Enzymology-related studies have demonstrated that phospholipase C (PLA C) and ENPP6 can transform MV membrane lipids into phosphoethanolamine (PE) and PCho [45]. Eventually, organic phosphate from either of these sources will be transformed into Pi within the MVs by PHOSPHO1, and Pi is the exact active state in biomineralization.
The transportation of Ca2+ seems to be relatively simple, possibly because Ca2+ is generally adapted to inorganic forms. Before of the secretion of MVs, the permeability of the mitochondrial membrane changes dramatically, and numerous Ca2+ ions leak into MVs [25]. After being secreted into the ECM, MVs are capable of accumulating Ca2+ from the ECM with the assistance of a chemical gradient, which causes Ca2+ to successfully enter the calcium channel AnxA5 [51]. With the augmentation of Ca2+ and Pi, crystals consisting of ACP and OCP are generated from the MV contents. These crystals begin to construct covalent bonds with PS on the internal membrane and, hence, evolve into stable acidic phospholipid Ca2+—Pi complexes (PS—CPLX) [35].
Thereafter, OCP, ACP, and other crystals will be reconstructed and gradually replaced by HAP, with minerals of PS—CPLX expanding to fill the entire intraluminal space, destroying the vesicle membrane of MVs, and ultimately growing mineralized nodules. Thus, MV-mediated mineralization is completed [19,66].
Collagen mineralization
Mineralized nodules derived from MVs are always universally anchored in adjacent collagen fibrils and appear to be a necklace-like structure. However, there are still broad unmineralized areas among each mineralized nodule or different collagen fibril bundles. How the biomineralization process of these zones proceeds is a question to be addressed in mechanistic studies and is known as collagen mineralization. To date, there are at least two widely accepted pathways in collagen mineralization, which are termed hole zone theory and collagen fibril superhelix theory [19,25,67].
From the view of hole zone theory, the microenvironment for mineralization activates enzymes in the tissue fluid, leading to proteoglycan degradation and unmineralized gap shrinkage. Inorganic electrolytes thus start to accumulate, which ultimately form crystals to fill the gap [68]. Furthermore, collagen fibril bundles are also capable of narrowing the unmineralized area by transforming themselves into superhelix structures, which strengthens the anchoring force to mineralized nodules. As a result, mineralized nodules can utilize various enzymes in the ECM effectively and enlarge in size, fuse, and gradually fill the unmineralized gap [39,69]. Under the biological activity of hole zone and collagen fibrils, the process of collagen mineralization is completed. Ultimately, hard tissues are completely mineralized and grow to maturity.
MVs in Periodontal Development and Alveolar Bone Regeneration
The developmental process of periodontal tissues
Periodontal tissues that spread over the tooth are regarded as important connective tissues for the human body. They can provide the necessary mechanical retention force for teeth (regardless of whether they are natural teeth, dental implants, or dentures) and promote tooth development through blood and nerve supply [1,2,4,5]. To our current knowledge, periodontal tissues include three elementary components: periodontal ligaments, cementum, and alveolar bone, all of which are derived from the dental follicle tissues of tooth germ [1,70,71]. Furthermore, cementum and alveolar bone are physically hard and possess a similar developmental process resembling that of other hard tissues [2 –5]. Admittedly, this process is closely associated with biomineralization, which relies on MVs [24,72] (Fig. 5).
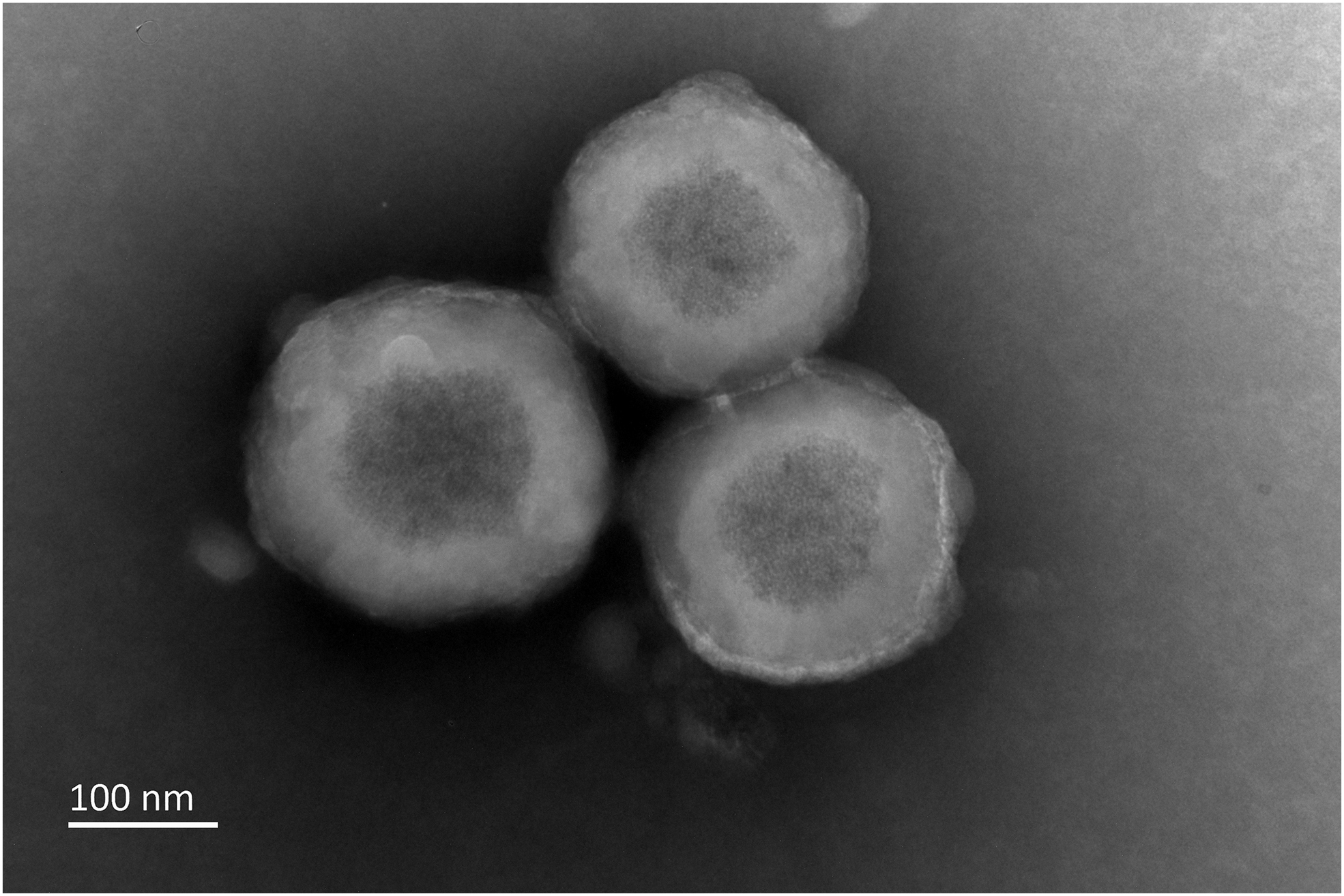
Ultrastructure of MVs produced by DFCs. This figure was captured by TEM. DFCs are the precursor cells in periodontal development. After an osteogenic induction of DFCs for a period, MVs were isolated according to the modified MV isolation protocol. DFC-derived MVs have a typical vesicular structure (∼200 nm in diameter). Furthermore, the vesicle membrane is relatively thick, and some needle-like minerals of high density can be found in the contents. DFC, dental follicle cell; TEM, transmission electronic microscopy.
Once periodontal tissues start to germinate, dental follicle cells gradually differentiate into fibroblasts, cementoblasts, and osteoblasts, which are located in the central region, proximal tooth, and proximal bone side, respectively [73 –75]. Periodontal ligaments are usually built in the early stage [76 –78], and this process is initiated by fibroblasts. In the central region, several Sharpey's fibers are created by fibroblasts, connecting the tooth root to the bone. Subsequently, fibroblasts continue to secrete matrix to multiply the quantity of the fibers, which simultaneously boost dental follicle tissue remodeling and evolve into periodontal ligaments with abundant and orderly arranged fibers [5,70,79]. In the meantime, cementoblasts cross the epithelial barrier, surround the tooth root, and secrete a considerable amount of organic matrix, including cementum protein-1 (CEMP1), cementum attachment protein (CAP), and cementum growth factors [5,80 –82]. While the organic components become saturated to a certain extent, cementoblasts are triggered to produce batches of MVs to induce the degradation of cementum-related proteins on the proximal tooth side and facilitate the biomineralization process [24,83,84]. As a result, mature cementum tissue is generated and tightly adheres to the tooth root [71,74,78]. Likewise, studies have reported that the biogenesis of alveolar bone has a strong correlation with MVs [24,85]. After undergoing osteogenic differentiation, osteoblasts located on the proximal bone side can secrete some proteins, such as type I collagen (COL1), bone morphogenetic protein (BMP), and periostin. In this special condition, MVs are produced for further biomineralization. They mediate biomineralization, fuse into mineralized nodules, and gradually construct the matrix of alveolar bone. Accompanied by the thorough completion of biomineralization, periodontal tissues are finally constructed, and the secretion of MVs ends [24,74].
Current therapies for alveolar bone defects
Despite the stable structure of hard tissue, alveolar bone is susceptible to being affected by certain prevalent diseases, such as periodontitis, periapical inflammation, and cysts, which corrode the bone and cause defects. Worse still, alveolar bone defects are likely to result in tooth instability and tooth loss, which severely interferes with the ordinary physiological movements (such as chewing and pronunciation) of the organism [6 –9]. In this case, how to repair the alveolar bone defects effectively and maintain tooth stability to secure physiological activity is a real challenge for clinicians.
To date, the clinical therapeutic techniques for alveolar bone defects mainly include GTR, autologous bone grafting, and heterogeneous bone filling. Although these therapies have resulted in good outcomes in some patients upon clinical application, many limitations still exist. The therapeutic indication of GTR is always too narrow, which makes it difficult to address some complex cases (such as two-wall defects), and high technical sensitivity is required for surgical operation [9]. Although autologous bone grafting has been acknowledged as an effective method, it causes secondary trauma to normal tissues [8]. The heterogeneous bone filling technique lacks osteogenic and osteoinductive potential, so its ability to promote self-healing is not comprehensive [86]. In addition, the current treatments for alveolar bone defects present a universal problem: their potential to regenerate hard tissues is finite, while the therapeutic period is relatively long. To improve the prognosis, it is urgent to exploit therapies that are more effective [6,87].
Prospects of MVs for alveolar bone regeneration
As naturally derived particles from developing hard tissues, MVs are inherently endowed with ideal biocompatibility and biological effects, as we discussed. Although sufficient therapeutic research on treating alveolar bone defects with MVs has been lacking to date, continued advances in MV-related biotechnologies provide favorable conditions for the successful application of MVs, and some desired positive effects of hard tissue regeneration can be found in the various published studies.
Previously, Boskey's group proved that materials made from hydrogels can be used as ideal carriers of MVs. By securing the stability of MVs, hydrogels could enhance the activity of enzymes in MVs and ultimately promote biomineralization [88]. However, due to the deficiency of isolation techniques at that time, MVs were often limited in yield and unlikely to be used on a large scale. Researchers were enthusiastic to explore new ways to increase MV production, and Balcerzak et al. designed the modified MV isolation protocol in 2008, which not only upgraded the yield and purity of MVs but also maintained the activity of MV proteins [29]. This method has been confirmed to be valid and was regarded as the standard protocol in the following studies. After employing and improving the above MV-related biotechnologies, Kunitomi et al. utilized collagen hydrogel materials to load osteoblast-sourced MVs in vitro [89], and the results showed that numerous MVs had accumulated sufficient crystals interiorly and been transformed into mineralized nodules. A significant number of mineralized nodules then fused into a wide range of mineralized belts and eventually formed recognizable hard tissues, which strongly boosted the subsequent application of MVs. Notably, based on the biological structure of MVs, Wang et al. created bioinspired MVs through chemical synthesis techniques. These particles, consisting of the polyglycolic acid copolymer and black phosphorus, were prepared by evaporation and exhibited good stability and biocompatibility. Moreover, in the C57 mouse model, the bioinspired MVs effectively activated osteoblasts in situ and reconstructed the calvarial defects with a considerable amount of new bones. Although these vesicles were not derived from organisms, they have been proven to be effective in hard tissue repair. It is promising that naturally derived MVs may act as powerful stimulators in hard tissue regeneration [32]. More recently, Bolean et al. examined AnxA5, which settled on MV membranes, and were found to be a good medium for MV adhesion, and materials with a good binding of AnxA5 are likely to provide a satisfying loading efficiency for MVs, which can direct the invention of new scaffolds [51].
The repair and regeneration of alveolar bone have both common (such as a long formation period and inaccurate efficacy) and exclusive characteristics compared with those of other hard tissues. For example, alveolar bone defects are always caused by periodontitis, and the moderate control of inflammation becomes an imperative prerequisite for tissue regeneration. Previously, scientists investigated the feasibility of treating periodontitis with EVs from adipose-derived stem cells, and the outcome reflected that the EV group was effective in immunoregulation and alleviated neutrophil infiltration to some extent [26]. Afterward, Chew et al. used EVs derived from periodontal stem cells to test their therapeutic potential in tissue regeneration. In the rat model of mesial alveolar defects, some newborn tissues emerged in the injured region, but failed to fully repair the defects [27]. Compared with other EV subtypes, MVs have more potential in controlling tissue inflammation [29]. For instance, Bessueille et al. reported that TNAP, a major component of MVs, could accelerate the process of ATP assimilation and upgrade anti-inflammatory cytokine expression in a transgenic mouse model [31]. Furthermore, by inhibiting the receptor activator of the nuclear factor κ-B ligand (RANKL) pathway, MVs could suppress osteoclastogenesis and effectively alleviate pathological bone absorption [90]. Moreover, components, including numerous enzymes, can secure the ability of MVs to mediate biomineralization, and the positive cases in hard tissue regeneration of MVs that have been demonstrated are prospectively linked to more remarkable therapeutic results in alveolar bone regeneration. In future studies, attention should be focused on optimizing MV-related biotechnologies comprising isolation, storage, and delivery approaches and confirming their efficiency in a wide range of models, which is a prerequisite to maximize the biological potency and promote the general application of MVs.
Accordingly, unlike other EVs, MVs are expected to play a unique role in treating larger, more complex, and different types of alveolar bone defects and in promoting tissue regeneration (Fig. 6).
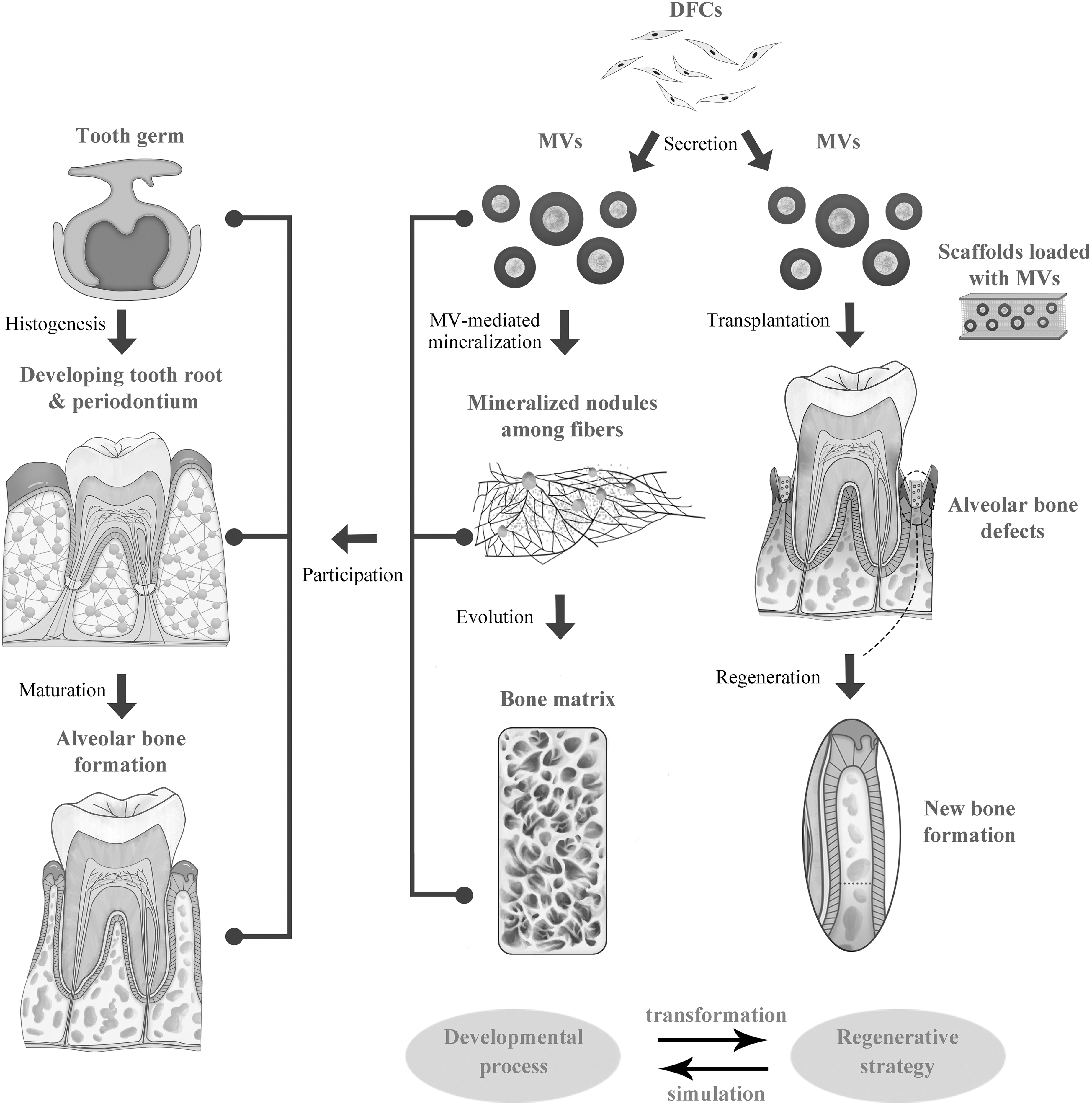
The role of MVs in alveolar bone development and regeneration. The diagram illustrates the theme of this review, which focuses on transforming the development process of alveolar bone into a regenerative strategy. Since periodontium originates from the dental follicle, in alveolar bone formation, a variety of MVs are generated by DFCs and then adhere to collagen fibers. Then, MVs can mediate biomineralization and evolve into mineralized nodules. In this case, the bone matrix is constructed. Notably, these biological activities are sustained throughout germination, histogenesis, and maturation of teeth and alveolar bone. Based on this knowledge, we propose MV-centered therapies for alveolar bone regeneration. MVs can be collected from DFCs and then transplanted into the alveolar bone defects combined with scaffolds (for periodontitis, periodontal scaling is necessary beforehand). The scaffolds are mainly bone replacement materials, and MVs are regarded as stimulators to enhance the therapeutic efficacy. As a result, the regeneration of new bone can be achieved, which promotes the healing of alveolar bone defects.
Conclusion
As discussed above, MVs present many unique features with respect to morphology, molecular composition, biological mechanisms, and, especially, biological functions in periodontal development. The entire vesicle structure is supported by a highly organized phospholipid bilayer that provides a sufficient capacity to accommodate MV contents, including multifunctional proteins, electrolytes, and minerals. Relying on these compositional features, MVs can promote biomineralization and construct cementum and alveolar bone. Moreover, MV-related biotechnologies have been utilized in some in vitro and in vivo cases and manifested positive effects in regenerating hard tissues and anti-inflammation. Based on this evidence, we propose that an MV-centered tissue regenerative strategy is promising for treating refractory alveolar bone defects and improving the clinical prognosis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key Research and Development Program of China (No. 2017YFA0104800).