
Abstract
Bronchopulmonary dysplasia (BPD) is a serious chronic lung disease in premature newborns, with high morbidity and mortality rates. Mesenchymal stem cell (MSC) transplantation has developed into a promising approach to alleviate BPD. Small extracellular vesicles, which are an important therapeutic component of MSCs, have been reported to be effective in a mouse model of BPD. However, the affected cell types and detailed underlying mechanisms are unclear. In this study, we found that human umbilical cord mesenchymal stem cell-derived small extracellular vesicles (hucMSC-sEVs) were successfully absorbed by lung tissue after intratracheal administration, and remained in the lungs for at least 72 h. The results showed that hucMSC-sEVs restored alveolar structure and lung function, and ameliorated pulmonary hypertension in a rat model of BPD. The number of Ki-67-positive lung cells were improved, while the number of TUNEL-positive lung cells were reduced in our hucMSC-sEV-treated BPD model. Additionally, SP-C staining (a marker of type II alveolar epithelial cells, TIIAECs) and CD31 staining (a marker of pulmonary vascular endothelial cells, PVECs) were both increased in a hyperoxia-induced BPD model treated with hucMSC-sEVs. In vitro, under hyperoxic conditions, the tube-like structure formation was improved in human umbilical vein endothelial cells, and the proliferation was increased and the apoptosis was attenuated in MLE-12 cells treated with hucMSC-sEVs. Furthermore, we observed downregulated expression of PTEN and cleaved-caspase3, and upregulated expression of p-Akt and vascular endothelial growth factor-A in our hucMSC-sEV-treated BPD model. In conclusion, hucMSC-sEVs improved alveolarization and angiogenesis in a rat BPD model by protecting TIIAECs and PVECs, which were associated with the PTEN/Akt signaling pathway.
Introduction
Bronchopulmonary dysplasia (BPD) is a chronic lung disease in preterm infants that requires mechanical ventilation and oxygen therapy, and it is characterized by impaired alveolar and pulmonary vascular development [1]. Although the survival rate of preterm infants has improved with the application of antenatal glucocorticoids, surfactants, caffeine, vitamin A, and the gentle ventilation strategy [2], the incidence of BPD remains high [3]. Neonates with BPD often manifest poor long-term outcomes, with increased infections, lung function impairment, pulmonary hypertension (PH), and a high mortality rate [4,5]. Treatments for BPD at present are mainly supportive, and further studies are urgently needed to explore novel approaches. Cell-based therapies constitute one such promising approach.
Mesenchymal stem cell (MSC) transplantation has been demonstrated to exert therapeutic effects in both preclinical models and clinical trials of BPD [6 –10]. Accumulating evidence indicates that MSCs repair injured lung tissue primarily through paracrine effect rather than by direct engraftment and differentiation [11,12].
MSC-derived small extracellular vesicles (MSC-sEVs) are considered to be one of the key MSC secretomes. They have been studied for their potential paracrine role and play an important part in cell–cell communication. Compared with live cell-based therapy, sEV-based, cell-free therapy is less immunogenic and more stable; sEVs can also be stored frozen without loss of activity and can cross the blood-brain barrier, creating a potentially safe and effective option [13 –17]. Several studies have found that MSC-sEVs exert protective effects on animal models that mimic BPD [18 –22] as they can block inflammation, improve lung structure and function, and ameliorate PH. However, the affected cellular populations and the related underlying mechanism of action remain unknown.
Hyperoxia is reported to impair alveolarization and angiogenesis [23], In this study, we investigated whether human umbilical cord mesenchymal stem cell-derived sEVs (hucMSC-sEVs) exerted an effect on both type II alveolar epithelial cells (TIIAECs) and pulmonary vascular endothelial cells (PVECs) in a rat model of BPD. Given that PTEN regulates the downstream proteins Akt and vascular endothelial growth factor (VEGF), thus contributing to the regulation of cell survival and angiogenesis [24,25], it is unfortunate that the function of PTEN in lung development in response to hyperoxia has not been fully investigated.
The aim of this study was to analyze the effect of hucMSC-sEVs on a rat model of BPD and investigate its underlying regulatory mechanism. We found that treating hyperoxia-induced injuries with hucMSC-sEVs improved survival and angiogenic effects both in vivo and in vitro, and these findings were associated with the activation of the PTEN/Akt signaling pathway.
Materials and Methods
Animals
Sprague Dawley (SD) rats obtained from the Experimental Animal Center of Chongqing Medical University were maintained at the Animal Laboratory Center of Pediatrics, in the Children's Hospital of Chongqing Medical University. All animal procedures and protocols were approved by the Ethics Committee of Chongqing Medical University (CQLA-2018-0407).
Rat model of BPD
Newborn pups were exposed to either 85% oxygen or room air (RA) from birth (P0) until postnatal day 14 (P14). Newborn rats were divided into three experimental groups: the room air group (RA+PBS, n = 13), hyperoxic group (BPD+PBS, n = 13), and hyperoxia with hucMSC-sEVs group (BPD+sEV, n = 13). The BPD and RA groups contained the same number of rats. Dams were alternated every 48 h from hyperoxia to normoxia to minimize oxygen toxicity. Treatment groups received 40 μL of intratracheally administered MSC-sEVs (20 μg of protein) on P7. This dose was selected based on previous works [15,22,26 –29], and the room air and sham-treated hyperoxic groups received 40 μL of intratracheally administered phosphate-buffered saline (PBS).
Cell culture
The hucMSCs were provided by the Chongqing Stem Cell Engineer Research Center and cultured in DMEM/F12 containing 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin. Cells between passage 4 and passage 6 were used in this study. When hucMSCs had grown to 60%–70% confluency, the medium was replaced with DMEM/F12 containing 10% sEV-free FBS for 48 h. We then collected MSC-conditioned medium (CM) for the isolation of sEVs. sEV-depleted FBS was obtained by overnight ultracentrifugation at 120,000 g at 4°C, and we collected the upper 90% of the supernatant as sEV-free FBS.
Human umbilical vein endothelial cells (HUVECs) and mouse lung epithelial-12 (MLE-12) cells were purchased from the Shanghai Institute of Biochemistry and Cell Biology and maintained in RPMI1640 and DMEM respectively, which were supplemented with 10% FBS and penicillin (100 U/mL)/streptomycin (100 μg/mL).
To stimulate hyperoxic conditions in vitro, HUVECs and MLE-12 cells were placed in 95% O2/5% CO2 at 37°C in a tightly sealed chamber for 24 h.
Small extracellular vesicle isolation
sEVs were isolated by serial centrifugation as previously described, with the following slight modifications [30]. In brief, hucMSC-CM was successively centrifuged at 500 g for 10 min, at 2,000 g for 20 min, and at 10,000 g for 30 min to remove dead cells and cellular debris. The supernatant was then ultracentrifuged at 110,000 g for 70 min (SW28; Beckman Coulter). The sEV-enriched pallet was passed through a 0.22 μm filter (Millipore), washed with PBS, and recentrifuged at 110,000 g for 70 min (90Ti; Beckman Coulter). All procedures were performed at 4°C. The purified sEVs were resuspended in 200 μL of PBS and stored at −80°C. To determine the number of hucMSC-sEVs, we quantified the total sEV protein concentration using a BCA protein assay kit (Beyotime) as per the manufacturer's recommendations.
Transmission electron microscopy
sEVs were diluted 50-fold, and a drop of diluent containing sEVs was pipetted onto carbon-coated electron microscopy grids and left to absorb for 30 min at room temperature. Then, we removed all excess fluid, and the samples were negatively dyed with uranyl acetate for several seconds. The morphology of the sEVs was observed using a transmission electron microscope (TEM, Hitachi, S-3000N, Japan).
Nanoparticle tracking analysis
Ten microliters of sEVs was diluted in 1 mL of filtered PBS, and we determined the size distribution using a Zetasizer Nano S90 (Malvern). Videos images of particle movement were collected and analyzed using nanoparticle tracking analysis (NTA) analytical software.
Western blotting analysis
MSCs, hucMSC-sEVs, and lung tissue samples were lysed in RIPA buffer containing phenylmethanesulfonylfluoride and phosphatase inhibitors. The protein concentration was determined using a BCA protein assay kit (Beyotime).
Equal quantities of proteins were resolved using SDS-PAGE and then transferred onto PVDF membranes (Millipore). Membranes were blocked in 5% nonfat milk for 1 h at room temperature and then incubated with the following antibodies: mouse anti-CD63 (1:500, sc5275; Santa Cruz), rabbit anti-Alix (1:500, JM85-31; HuaAn Biotechnology), rabbit anti-PTEN (1:1,000, 9188s; CST), rabbit anti-Akt (1:1,000, 9272s; CST), rabbit anti-p-Akt (1:1,000, 4060p; CST), rabbit anti-cleaved-caspase3 (c-caspase3) (1:1000, 9661s; CST), rabbit anti-VEGF-A (1:1,000, ab46154; Abcam), and rabbit anti-GAPDH (1:1,000, 10494-1-AP; Proteintech) overnight at 4°C. The membranes were then incubated in goat anti-rabbit (1:5,000, SA00001-2; Proteintech) or goat anti-mouse (1:5,000, SA00001-1; Proteintech) antibody at room temperature for 1 h.
The bands of proteins were visualized using ECL Select Western Blotting Detection Reagent. We used ImageJ software to calculate the gray values of the target protein. GAPDH was used as an internal control.
Small extracellular vesicle labeling and tracking in lung tissue and cells
sEVs were labeled with PKH-26 (Sigma-Aldrich) and washed in PBS with centrifugation at 110,000 g for 60 min. In vitro, PKH-26-labeled sEVs were cocultured with MLE-12 cells or HUVECs for 0, 2, 12, and 24 h at a concentration of 80 μg/mL, and then fixed with 4% paraformaldehyde (PFA) for 20 min. Cells were stained with DAPI for 15 min. In vivo, PKH-26-labeled sEVs were intratracheally injected into BPD rats on P7, and we collected lung tissues 0, 2, 12, 24, and 72 h postinjection; each time point contained three rats. Fluorescence was observed by Leica laser confocal microscopy (C2+ system, Nikon, Japan).
Tissue preparation
The animals were anesthetized with an intraperitoneal injection of pentobarbital sodium (40 mg/kg) on P14. After opening the thorax, the right lung was removed and frozen at −80°C, while the left lung was fixed overnight in 4% PFA solution. The hearts were placed in saline.
Immunofluorescence assay
Fixed left lungs were embedded in glue (Sakura, Japan) and cut with a microtome at 10 μm at −20°C (Leica CM1950). Frozen sections and cells were then fixed with 4% PFA and blocked with 10% BSA. Subsequently, the samples were stained with mouse anti-CD31 (1:100, ab64543; Abcam), rabbit anti-SP-C (1:50,10774-1-AP; Proteintech), and rabbit anti-Ki-67 antibodies (1:50, bs-2130R; Bioss, China) overnight at 4°C, and then washed and incubated with Fluor cy3-conjugated secondary antibodies (Proteintech). We used a TUNEL (11684795910; Roche, Switzerland) assay kit and incubated either frozen sections or cells for 1 h at room temperature. Triton-X 100 (0.3%) was used before Ki-67 and TUNEL staining.
Cell nuclei were counter-stained with DAPI for 15 min, and we observed fluorescence using a Leica laser confocal microscope (C2+ system; Nikon, Japan). Five random images were captured in each section at 200 × magnification for quantification of CD31, and at 400 × magnification for quantification of SP-C, Ki-67, and TUNEL. Each group contained at least four animals, and at least three sections were examined from each rat. The number of CD31-positive vessels (<100 μm) was counted for each section, and the number of SP-C, Ki-67, and TUNEL-positive cells per high power field was measured using Image-Pro Plus.
Assessment of alveolar size by mean linear intercept
Fixed left lungs were immersed in 75% ethanol before subsequent processing, embedded in paraffin, sectioned at 5 μm (Leica RM2265), and stained with H&E. Five random photomicrographs of different sections were captured at 200 × magnification. Alveolar size was estimated by measuring the mean linear intercept (MLI) using Image-Pro Plus [21].
PH-induced RVH
To quantify the degree of PH-induced right ventricular hypertrophy (RVH), hearts were excised and divided into right ventricle (RV) and left ventricle (LV) plus interventricular septum (S), and then weighed. The RV to LV + S weight ratio was calculated as Fulton's index (FI).
Lung function
Rats were anesthetized with pentobarbital sodium (40 mg/kg), intubated after tracheostomy, and mechanically ventilated at a rate of 150 breaths/min, with a tidal volume of 8 mL/kg and a positive end-expiratory pressure (PEEP) of 3 cm H2O using a computer-controlled small-animal ventilator (Emka). We measured lung resistance (LR) and dynamic compliance (Cydn) every 5 s.
Tube formation assay
HUVECs were maintained in RPMI1640 and supplemented with 1% FBS and penicillin (100 U/mL)/streptomycin (100 μg/mL) for 24 h on either RA or hyperoxic conditions (HYP). The hucMSC-sEVs (80 μg/mL) were cocultured with HUVECs for 24 h under hyperoxic conditions (HYP+sEV), after which 7.5 × 104 cells per well were plated on 48-well plates coated with growth factor-deprived Matrigel (356230; Corning) to evaluate angiogenesis. Three wells were plated per group, and for each group, the procedure was repeated three times. Five hours later, at least three random images of tube formation were acquired for each well with a light microscope at 200 × magnification, and total tube lengths formed were analyzed using ImageJ.
CCK-8
MLE-12 cells were plated into 96-well culture plates at an initial density of 103 cells per well. Twenty microliters of CCK-8 reagent (KQ749, DOJINDO, Japan) was added to each well after treatment, and the cells were incubated for 2 h at 37°C. We assessed the absorbance of each well at 490 nm using a microplate system (Gen5 GHS 2.0).
Flow cytometry
The apoptotic rate of MLE-12 cells was analyzed by flow cytometric analysis with an Annexin V-APC/7AAD apoptosis detection kit (KGA1026, KeyGEN BioTECH, China) in accordance with the manufacturer's instructions. In brief, MLE-12 cells were digested with EDTA-free trypsin, washed with PBS after treatment, and then resuspended in 200 μL of binding buffer; 5 μL of APC-labeled Annexin V and 5 μL of 7-AAD reagent were then added to the cell suspension. The mixture was incubated in the dark for 15 min at room temperature and then analyzed using flow cytometry (BD Biosciences).
Statistical analysis
We used GraphPad Prism software (Version 5.0, San Diego, CA) for statistical analysis. Data are presented as the mean ± SEM (standard error of the mean). A two-tailed, unpaired Student's t-test was used to compare two groups, and one-way analysis of variance (ANOVA) followed by Tukey's post hoc test was conducted to determine statistically significant differences among more than two groups. Statistical significance was defined as P < 0.05.
Investigators were blinded to experimental groups for histological analysis and physiological measurements.
Results
hucMSC-sEVs ameliorate hyperoxia-induced lung injury in a rat model of BPD
We successfully established our rat model of hyperoxia-induced BPD, and the MLI (which is an indicator of alveolar size) was significantly higher in the BPD group compared with the RA group on P7 (65.30 ± 2.77 vs. 55.70 ± 1.40 μm, P < 0.05), P14 (82.74 ± 3.70 vs. 58.64 ± 1.96 μm, P < 0.01), and P21 (92.08 ± 2.52 vs. 59.28 ± 1.84 μm, P < 0.001), without obvious pathologic changes in BPD on P3 (33.53 ± 1.704 vs. 37.65 ± 0.64 μm, P = 0.06) (Fig. 1A, B).

Morphological changes in lung tissue induced by hyperoxia and characterization of hucMSC-sEVs.
Since hucMSC-sEVs have been reported to be effective components of MSCs, we isolated them from the CM through serial centrifugation as described previously [30]. TEM revealed that hucMSC-sEVs exhibited typical saucer-shaped morphologic features (Fig. 1C) and showed positive expression of the characteristic markers CD63 and Alix as determined by western blot analysis (Fig. 1D). Furthermore, the NTA showed that the diameters of the sEVs ranged from 70 to 300 nm, with a peak size distribution of 175 nm (Fig. 1E).
To determine whether hyperoxia-induced lung injury responded to hucMSC-sEVs, newborn rats were exposed to 85% O2 from P0 to P14, and age-matched control pups were placed on room air. sEVs (20 μg, protein) were then intratracheally administered to the treatment group on P7 when the pathological hallmarks of BPD were established, as defined in our hyperoxia-induced BPD rat model (Fig. 2A). The other groups received intratracheal injections of PBS on P7.
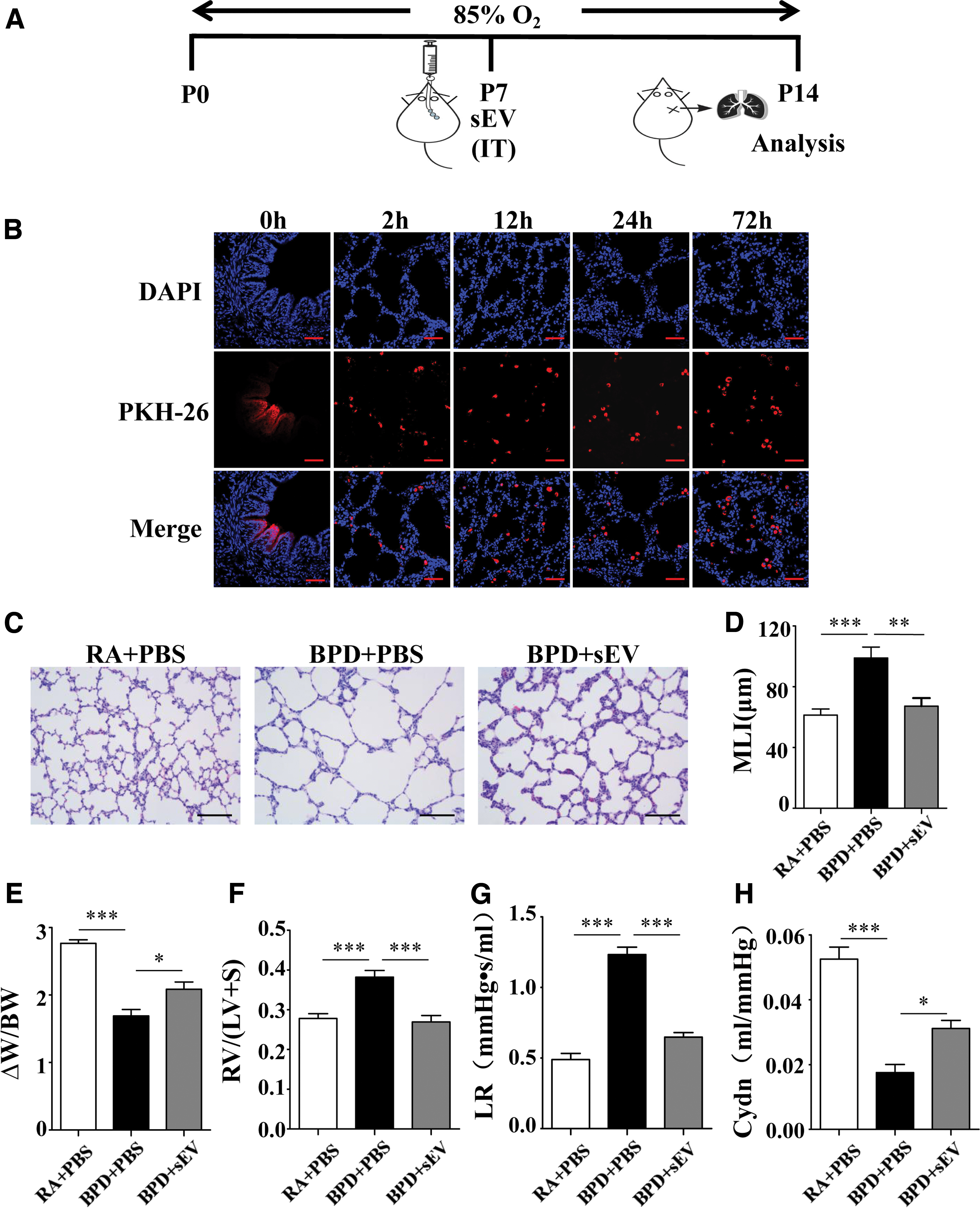
hucMSC-sEVs ameliorate hyperoxia-induced lung injury in a BPD rat model.
To determine how hucMSC-sEVs were internalized by pulmonary cells, sEVs labeled with PKH-26 were intratracheally injected into the BPD group on P7. We observed that the sEVs remained in the airways at 0 h, they were internalized into lung cells after 2 h, and this status was maintained for at least until 72 h. sEVs were primarily localized in the cytoplasm (Fig. 2B).
We subsequently observed that the hyperoxia-induced increase in MLI (98.93 ± 7.27 vs. 61.34 ± 3.99 μm, P < 0.001) was significantly ameliorated by hucMSC-sEV treatment (67.26 ± 5.32 μm, P < 0.01 vs. BPD) (Fig. 2C, D). The body weights of neonatal rats were measured at birth (BW) and on P14 (W), and the W-BW to BW ratio was calculated as a weight-gain rate, which was significantly decreased in the BPD group compared with the RA group (1.69 ± 0.09 vs. 2.77 ± 0.05, P < 0.001); administration of sEVs significantly improved the weight-gain rate relative to the BPD group (2.09 ± 0.11, P < 0.05 vs. BPD) (Fig. 2E).
Because RV hypertrophy suggests PH, we evaluated the degree of hyperoxia-induced PH, and found that FI was significantly elevated in the BPD group compared with the RA group (0.38 ± 0.02 vs. 0.28 ± 0.01, P < 0.001), and that hucMSC-sEV treatment attenuated the FI elevation compared with the BPD group (0.27 ± 0.16, P < 0.001 vs. BPD) (Fig. 2F), indicating that a reversal of PH occurred.
Pulmonary function testing was performed to determine the functional impact of altered lung structure. LR was significantly increased (1.23 ± 0.05 vs. 0.49 ± 0.04, P < 0.001) and Cdyn was significantly decreased (0.02 ± 0.002 vs. 0.05 ± 0.004, P < 0.001) in the BPD group compared with the RA group, and intratracheal administration of hucMSC-EVs significantly reversed the changes in LR and Cdyn compared with the BPD group (0.65 ± 0.03, P < 0.001, and 0.03 ± 0.002, P < 0.05 vs. BPD respectively) (Fig. 2G, H).
hucMSC-sEVs improve cell survival and angiogenesis following hyperoxia-induced lung injury
We used Ki-67 and TUNEL staining to determine the cell survival of different groups (Fig. 3A, C), and observed that the hyperoxia-induced diminution in the number of Ki-67-positive cells (10.92 ± 0.46 vs. 22.20 ± 3.27, P < 0.01) in rat lung tissue was significantly improved by hucMSC-sEV treatment (19.69 ± 1.24, P < 0.05 vs. BPD) (Fig. 3B). Moreover, the hyperoxia-induced increase in the number of TUNEL-positive cells in the lungs (2.91 ± 0.34 vs. 0.19 ± 0.06, P < 0.001) was significantly ameliorated by hucMSC-sEV treatment (1.31 ± 0.25, P < 0.05 vs. BPD) (Fig. 3D), indicating that the administration of hucMSC-sEVs improved proliferation and prevented apoptosis in hyperoxia-induced lung injury.
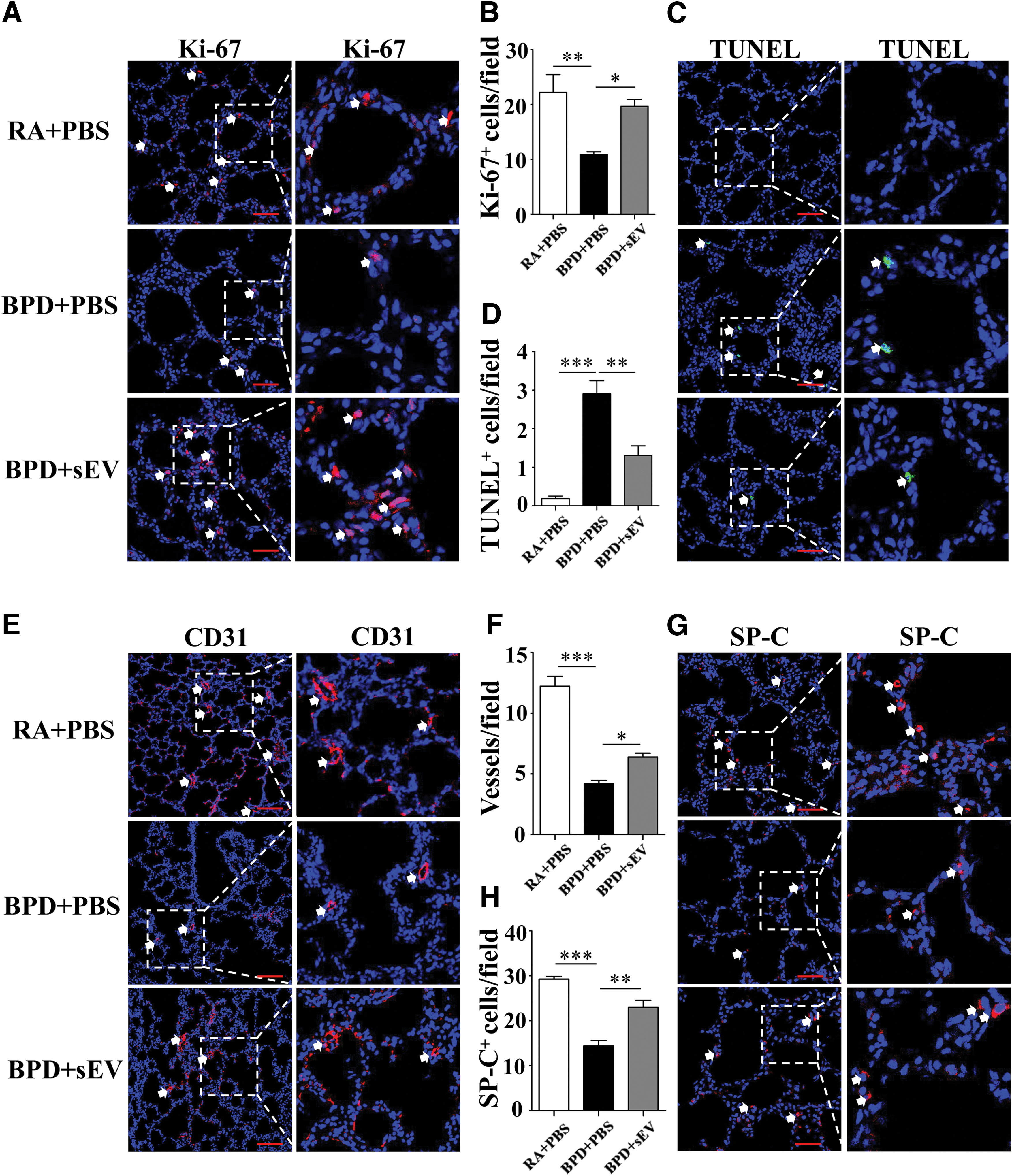
hucMSC-sEVs improve cell survival in lung tissue following hyperoxia-induced lung injury.
We performed lung section staining with CD31(a marker of PVECs) and SP-C (a marker of TIIAECs) to determine whether hucMSC-sEVs exerted an effect on these two types of cells (Fig. 3E, G). The BPD group manifested a significant loss of small vessels (<100 μm) compared with the RA group (4.20 ± 0.27 vs. 12.23 ± 0.81, P < 0.001), indicating impaired angiogenesis, whereas hucMSC-sEV treatment restored these vessels (6.39 ± 0.32, P < 0.05 vs. BPD) (Fig. 3F). The decreased number of SP-C-positive cells (14.38 ± 1.24 vs. 29.25 ± 0.61, P < 0.001) in rat lung tissue that resulted from hyperoxia was also significantly improved by intratracheal transplantation of hucMSC-sEVs (23.03 ± 1.50, P < 0.05 vs. BPD), indicating that the repair ability of the alveoli improved (Fig. 3H).
hucMSC-sEVs mediate angiogenesis and cell survival after hyperoxia-induced injury in HUVECs and MLE-12 cells
To better understand how hucMSC-sEVs interact with PVECs and TIIAECs, sEVs labeled with PKH-26 (red) were incubated with HUVECs and MLE-12 cells in 24-well plates for 0, 2, 12, and 24 h at 37°C. We noted that sEVs were internalized into both cell types after 2 h, they were primarily localized to the cytoplasm within 12 h, and at 24 h some sEVs aggregated at the nuclear membrane (Fig. 4A).
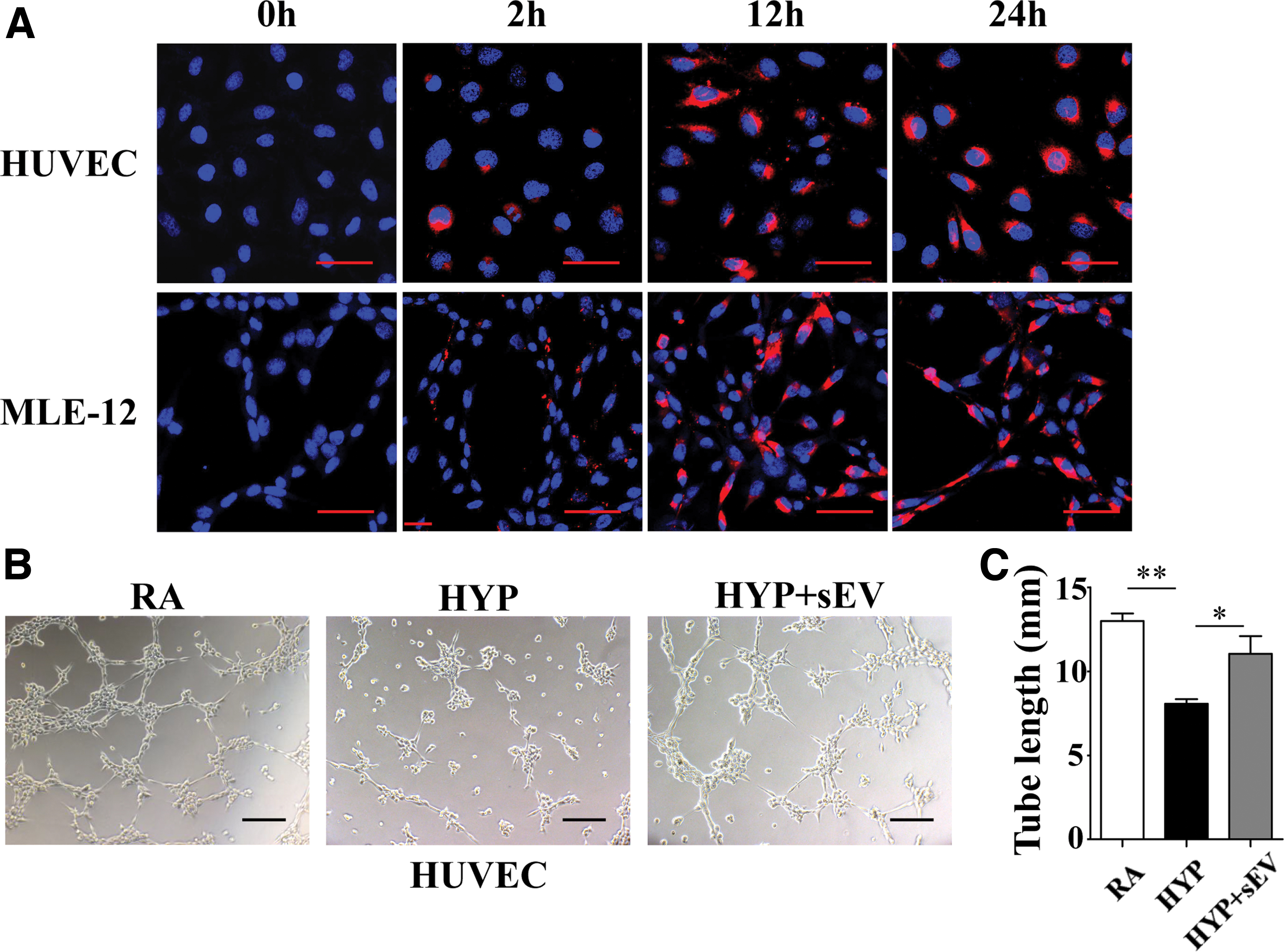
hucMSC-sEVs mediate angiogenesis following hyperoxia-induced injury in HUVECs.
We treated HUVECs and MLE-12 cells with 80 μg/mL of hucMSC-sEVs to assess the angiogenesis and cell survival induced by hyperoxia, respectively, and the results indicated that hyperoxia impaired the ability to form capillary tubes in HUVECs (8.07 ± 0.27 vs. 13.00 ± 0.44 mm, P < 0.01), while the total capillary tube length was significantly improved after treatment with hucMSCS-sEVs (11.04 ± 1.05 mm, P < 0.05 vs. HYP). This suggested a proangiogenic role for sEVs in vitro (Fig. 4B, C).
We next explored the cell survival status of hyperoxic MLE-12 cells treated with hucMSC-sEVs. Compared with the HYP group, MLE-12 cells treated with hucMSC-sEVs showed significantly increased Ki-67 staining (6.00 ± 0.51 vs. 2.50 ± 0.29, P < 0.01) (Fig. 5A, B) and improved viability (86.69% ± 5.12% vs. 70.58% ± 2.83%, P < 0.05) (Fig. 5C). Furthermore, hucMSC-sEVs significantly ameliorated TUNEL staining (0.44 ± 0.06 vs. 2.21 ± 0.34, P < 0.01) (Fig. 5D, E), and also suppressed the hyperoxia-triggered apoptotic rate of MLE-12 cells (8.93% ± 1.05% vs. 12.12% ± 0.32%, P < 0.05) (Fig. 5F, G). These results indicated that hucMSC-sEVs improved proliferation and attenuated apoptosis in response to hyperoxia in vitro.
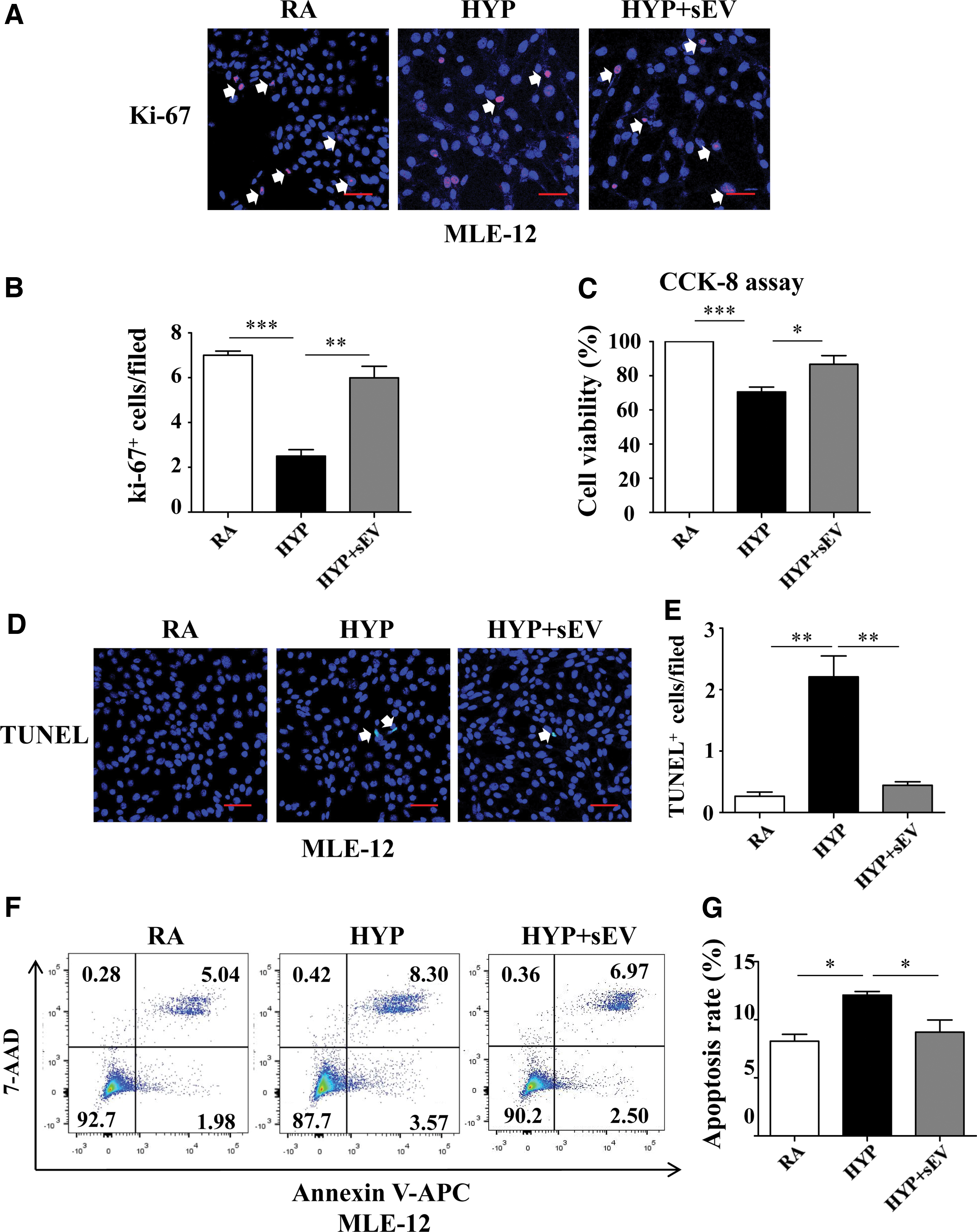
hucMSC-sEVs mediate cell survival following hyperoxia-induced injury in MLE-12 cells.
Administration of hucMSC-sEVs improves alveolarization and angiogenesis associated with the PTEN/Akt pathway
To investigate the mechanism by which hucMSC-sEV administration improves the impaired pulmonary alveolarization and angiogenesis induced by hyperoxia in a BPD rat model, we measured the levels of PTEN and Akt, in addition to downstream targets, such as caspase3 and VEGF-A, in whole rat lung tissue. The results of western blotting analysis showed that the expressions of PTEN and c-caspase3 were significantly suppressed (1.034 ± 0.14 vs. 1.80 ± 0.20, P < 0.05, and 0.69 ± 0.04 vs. 1.08 ± 0.11, P < 0.01, respectively), while the expressions of p-Akt and VEGF-A were significantly increased in the hyperoxic group that received hucMSC-sEV treatment (0.49 ± 0.03 vs. 0.33 ± 0.04, P < 0.05, and 0.48 ± 0.05 vs. 0.20 ± 0.03, P < 0.05, respectively) (Fig. 6A–E).
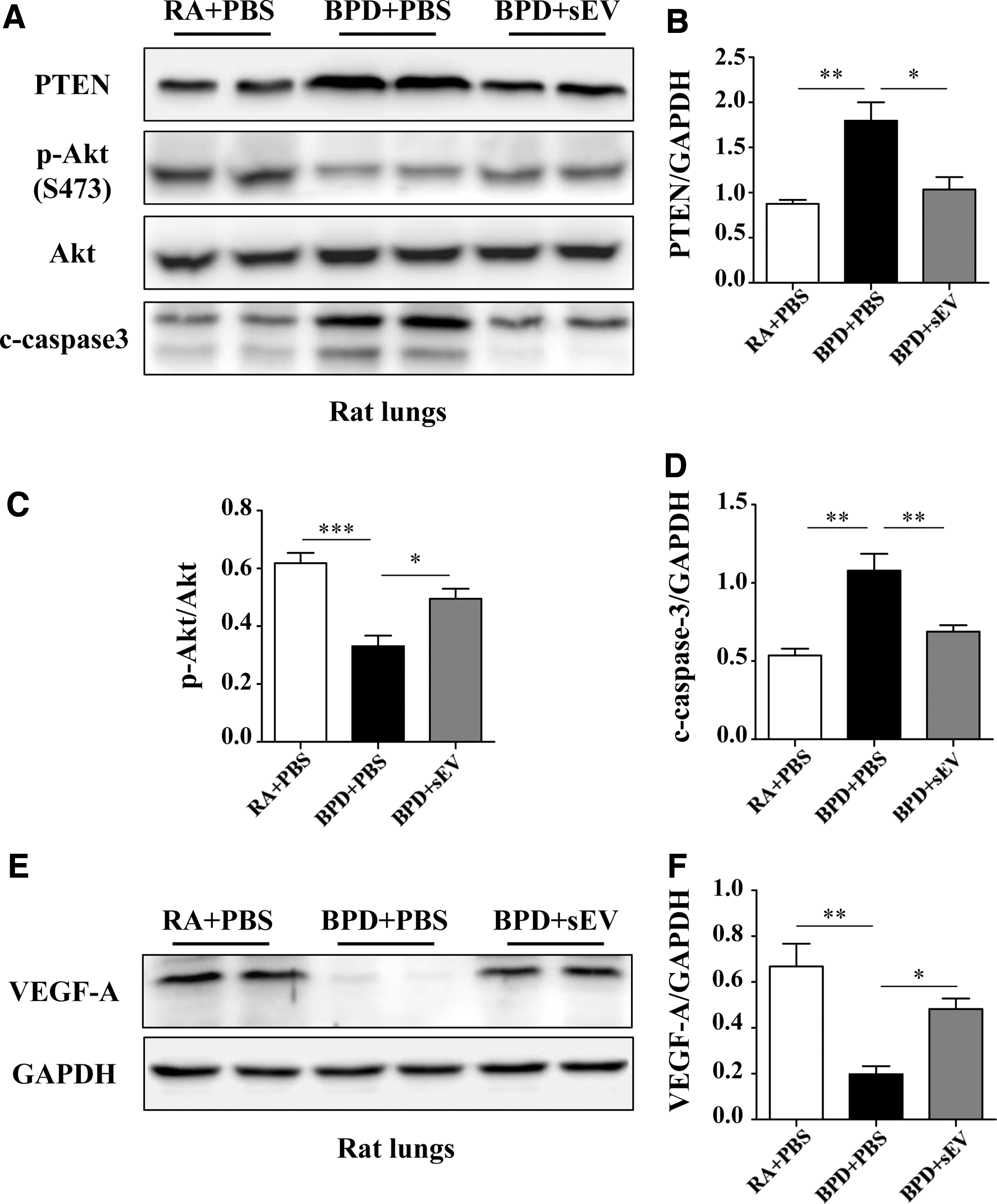
hucMSC-sEVs preserve lung alveolarization and angiogenesis in hyperoxia-induced lung injury associated with the PTEN/Akt-signaling pathway.
Collectively, these data suggested that hucMSC-sEVs improve pulmonary alveolarization and angiogenesis in a mechanism associated with augmented Akt activity and inhibition of PTEN.
Discussion
MSCs have provided new hope in the field of BPD treatment, and these are known to principally regulate injured tissues in a paracrine manner [11,12]. In addition to a large number of cytokines and growth factors secreted by MSCs, sEVs constitute a key secretome that has been studied to clarify their potential paracrine function, with great potential for the repair of damaged lung tissue [13 –17].
The optimal route and dose of treatment with respect to the use of sEVs for transplantation remain unclear. However, previous studies have shown that intratracheal, intravenous, and intraperitoneal administration of sEVs are all effective in a hyperoxia-induced BPD model [18 –22]. Takahashi et al. has shown that sEVs disappeared from serum within 30 min of intravenous administration [31], and that only a few labeled sEVs remained in the lung, while most accumulated in the liver and spleen [32,33]. We performed intratracheal administration of PKH-26-labeled hucMSC-sEVs and found obvious signs of aggregation in the lungs 2 h after injection, and that the aggregation lasted at least 72 h.
In addition, BPD occurs in preterm infants, who often have immature organs and immune deficiency. Intravenous administration of sEVs containing immunosuppressive compounds might have unknown systemic effects, and intratracheal administration may provide a safer manner for clinical use. A previous study also showed that intratracheal administration of MSCs was more effective than intravenous administration in protecting against hyperoxia-induced lung injury [34]. However, further analyses are needed to compare the effective dose range between different routes of administration of MSC-sEVs, and to determine their potential systemic effects.
Since quantifying sEVs themselves remains difficult, it is controversial to normalize the amount of MSC-sEVs used for comparative functional studies. At present, the therapeutic dose of sEVs is primarily based on the total amount of protein, particle counts, and the initial number of secreting cells [35]. Our results indicated that a single-dose intratracheal injection of hucMSC-sEVs (20 μg per rat) could control the adverse effects of hyperoxia on lung structure and function. Our protein concentration is consistent with doses (about 2 μg/g) used in previous studies of MSC-sEV therapy [15,22,26 –29], and further supports the idea that total protein concentration could be used as a quantification method for sEVs.
Nevertheless, other investigators preferred to use particle number for quantification, and suggested that both protein measurement and initial number of secreting cells were inaccurate and influenced by the isolation procedure [36]. Accurate particle number measurement may be possible only within a certain concentration and size range that varies by platform, and absolute values differ between different software [37]. However, none of these values are perfectly correlated with absolute sEV number, measuring multiple parameters at once and performing a dose–response experiment may be a good option for further studies seeking to normalize the amount of sEVs used in treatment.
The particle size and morphology of hucMSC-derived extracellular vesicles (hucMSC-EVs) in our study showed that the EVs in our preparation were MSC-sEVs [38]. The term “MSC-sEVs” in this study is synonymous with “MSC-derived exosomes,” as per recent recommendations [35,39].
Isolation of sEVs is mainly based on their size and density, but these factors cannot distinguish among different EV types, whose physical parameters of biogenesis overlap. Endosome-origin exosomes and plasma membrane-derived microvesicles are not practical to distinguish because they lack a specific biomarker [40]. Therefore, all MSC-sEVs described so far are probably mixtures of different EV types whose biogenesis is unclear. Thus, the standardized methods of isolation, purification, and characterization of sEVs require further refinement to facilitate the comparison of results obtained at different laboratories.
MSC-sEVs were first reported to mitigate hyperoxia-induced lung injury by modulating the lung macrophage phenotype [21]. Another study supported the contention that MSC-sEVs exerted therapeutic effects by regulating macrophages [20]. VEGF was reported to mediate the therapeutic efficacy of MSC-sEVs against hyperoxic lung injury [19,22], and Chaubey et al. [18] found a key role for sEV-associated factor TSG-6 in the treatment of a mouse BPD model. Little is known regarding the affected populations in MSC-sEV treatment of BPD.
Angiogenesis is necessary for normal alveolarization during the postnatal period of lung growth [41], and endothelial and epithelial cells are both important in pulmonary alveologenesis [42], TIIAECs is a type of alveolar epithelial cell that can participate in the repair of various lung injuries. Our study found that MSC-sEVs have a protective effect on TIIAECs and PVECs under conditions of hyperoxia-induced lung injury in rats, providing new insight about MSC-sEV therapy. Ameliorating loss of TIIAECs and PVECs suggested that improved angiogenesis and alveolarization occurred after MSC-sEV treatment.
Intriguingly, two previous studies confirming our findings indicated that MSC-sEVs exerted a significant effect in preventing the hyperoxia-induced reduction in pulmonary vessels [19,20]. However, two other investigations showed that sEVs increased small vessel density, but these results were not statistically significant [21,22]. The first two studies supporting our results used immunofluorescence technology as well, while the last two used immunohistochemical technology, and thus, the detection ability of the two methods for small pulmonary vessels may be different. In addition, because there are individual differences in animals between different laboratories, more experiments are needed to determine the effects of sEVs on pulmonary vessels in BPD.
Recent studies have shown that sEVs derived from MSCs play regulatory roles in tissue regeneration, apoptosis, angiogenesis, and immune regulation [20,21,28,43]. However, the specific molecular pathway by which sEVs treat the BPD model has not been reported. Our results suggested that hucMSC-sEVs improved cell survival and angiogenesis both in vivo, for lung injuries in the BPD model, and in vitro, for MLE-12 cells and HUVECs after hyperoxic exposure.
The prolonged exposure to hyperoxia is known to disrupt normal lung development, which is linked to abnormal expression of Akt and VEGF [44,45]. VEGF is a key component of pulmonary angiogenesis, the levels of which are attenuated in the lungs of infants with BPD [46], and recombinant VEGF administration has been recognized as preventing the occurrence of BPD in rats [47]. PTEN is the upstream gene of Akt, and the PTEN/Akt-signaling pathway was reported to regulate cellular proliferation, apoptosis, and angiogenesis [24,25,48].
However, its functional role in hyperoxia-induced lung injuries has not been elucidated. Our data showed that the expression levels of p-Akt and VEGF-A were significantly increased, and that the expression of PTEN and c-caspase3 were negatively correlated with p-Akt in the lungs after MSC-sEV treatment. This indicated that hucMSC-sEVs improve pulmonary alveolarization and angiogenesis through a mechanism associated with the activation of PTEN/Akt.
This study has several limitations that need to be addressed. Based on previous literature, we used only one dose of MSC-sEV and there is no dose–response data. Further studies are needed to identify a range of doses along with the functional assays to standardize exosome dosing, and to investigate different routes of administration. We primarily focused on TIIAECs and PVECs, but MSC-sEVs may affect other lung cells, including TIAECs, fibroblasts, and immune cells, under hyperoxic conditions. We also did not explore the cause–effect relationship between the treatment and molecular pathways. More thorough investigations are required to elucidate the complex mechanisms associated with MSC-sEV therapy, such as the other affected cell types and the specific components of sEVs that function in protection in the BPD model.
Conclusions
In summary, hucMSC-sEVs demonstrated a therapeutic effect on hyperoxia-induced injuries, both in lung tissue, MLE-12 cells, and HUVECs. In a hyperoxia-induced rat model of BPD, intratracheal administration of hucMSC-sEVs constitute a promising approach for the preservation of disrupted alveolarization and angiogenesis, as hucMSC-sEVs protect TIIAECs and PVECs, in a mechanism associated with the PTEN/Akt signaling pathway.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Chongqing Science and Technology Commission and Chongqing Health Commission (no. Cstc2014yykfC10003 and no. 2018ZDXM041).