
Abstract
Appropriate orthopedic force led to bone remodeling of mandibular condyle, while overloaded orthopedic force (OOF) induced condylar bone absorption. Bone absorption is ascribed to the imbalanced activities between osteoclasts (OCs) and osteoblasts (OBs), mechanism of which remains unclear. This study aimed to observe the condylar changes induced by OOF by mandible advancement appliance and to further investigate the role of mammalian target of Rapamycin (mTOR) and RANKL/OPG in osteoclastic differentiation of stem cells in vivo and in vitro. In vivo, the results of micro-CT analysis indicated that condylar bone resorption was induced by OOF through mandibular advancement appliance for 2 weeks and worsened time dependently. Morphologically, cartilage thickness was reduced, subchondral cortical bone line appeared not continuous, and subchondral bone exhibited irregular-shaped and owned uneven surface. The bone mineral density (BMD), bone volume/tissue volume (BV/TV), trabecular number (Tb.N), and trabecular thickness (
Introduction
Class II malocclusion, a common skeletal deformity of craniofacial region, may lead to occlusal disharmony and mandibular retrognathic facial pattern, limiting the airflow and affecting the facial esthetic [1,2]. Functional appliances are the effective procedures for Class II malocclusion through mandibular repositioning to correct jaw discrepancies sagittally and improve facial profile [3]. Clinical studies revealed that mandibular advancement using the bite-jumping orthodontic functional appliances could enhance condylar growth beyond natural growth [4,5]. However, unsuitable advancement of mandibular during orthodontic therapy may induce the iatrogenic temporomandibular joint osteoarthritis, characterized as condylar absorption, pain of the preauricular area or the masticatory muscle, clicking of joint, and limited mandibular movements [6,7]. Therefore, it is of utmost importance to explore the underlying mechanism of condylar absorption under overloaded orthopedic force (OOF) to avoid the condylar destruction and relieve discomfort of temporal-mandibular joint to optimize orthodontic clinical therapy.
Bone is under a highly dynamical balance through the tightly coupled activities of bone resorption by osteoclasts (OCs) and bone formation by osteoblasts (OBs) [8]. Normal bone remodeling is necessary for fracture healing and skeleton adaptation to mechanical stimuli or hormonal levels, while the imbalance of OBs and OCs, especially OC dysfunction, leads to bone lesions [9,10]. OBs, the major component of bone from mesenchymal stem cells (MSCs), secrete matrix proteins and transform mineral into the bone matrix [11]. OCs are large multinucleated cells with more than three nuclei, and its differentiation starts from the commitment of hematopoietic stem cells into the monocyte–macrophage lineage. Subsequently, premature OCs proliferate into mature OCs [12].
OB could affect OC formation, proliferation, differentiation, or apoptosis through several pathways, such as OPG/RANKL/RANK, RANKL/LGR4/RANK, Ephrin2/EphB4, and Fas/FasL pathways [13]. Among the signaling pathways, RANK/RANKL/OPG pathway plays a predominant role in OC differentiation. The fusion of OC precursors and activation of OC resorption were initiated by cells of the mesenchymal lineage, including OBs and osteocytes, due to the expression of receptor activator of NF-κB ligand (RANKL) and osteoprotegerin (OPG), a decoy receptor of RANKL [14,15]. RANKL/OPG was reported to regulate OC differentiation and osteolytic activity directly and modulated OC proliferation through tumor necrosis factor indirectly [16]. Besides, transforming growth factor (TGF), expressed by OBs and located in the bone matrix, is a potent MSC mitogen mediating OC differentiation. In TGF-enhanced mice, RANKL/OPG was activated and OC differentiation was enhanced, leading to an age-associated osteoporotic lesion [17].
The mammalian target of Rapamycin (mTOR) is identified as a cellular key factor in regulating most fundamental cellular functions like growth, proliferation, differentiation, autophagy, and apoptosis; its activity depends on growth factor, hormone, nutrition, and stress signals [18]. Furthermore, the mTOR also plays an important role in determining the bone homeostasis. In vivo, rapamycin, specific inhibitor of mTOR, was reported to significantly decrease endochondral bone growth and decline OC number of young rats [19]. In vitro, suppression of mTOR signaling by rapamycin in OCs could abrogate the OC proliferation from Nf1 heterozygous (Nf1+/−) mice through RANKL/OPG pathway, which is critical in Neurofibromatosis Type 1 treatment [20]. However, some scholars reported that rapamycin stimulated Trap-positive OCs in a dose-dependent manner, indicating that mTOR inhibited the differentiation of OCs [21]. For the lytic bone diseases, rapamycin inhibited the OC proliferation through C/EBPβ [22]. Although mTOR in OCs was reported to modulate OC differentiation themselves, to date little is known about the pivotal role of mTOR of OBs in OC differentiation under OOF.
In this study, we observed the alternations of condylar subchondral bone under OOF. We also explored the association between the activation of mTOR and RANKL/OPG secretion in OBs and osteoclastic differentiation of MSCs in vivo and in vitro. All our efforts were to provide new insights for the clinical treatment of Class II malocclusion.
Materials and Methods
Animals
Male Sprague–Dawley (SD) rats weighing 140–150 g were obtained from Animal Experiment Center of Qingdao University. Animal experiments were performed in accordance with the guidelines of the Animal Use and Care Committee of Qingdao University. All rats were housed under specific conditions: at room temperature (25°C), access to water, and in a 12-h light/12-h dark cycle.
Experimental procedures of rats
In the pre-experiment, 30 rats were randomly divided into three groups wearing a, b, and c appliances, respectively (Fig. 1A). For a and b groups, the mandibular advancement appliance (MAA) was mainly composed of a part of mental tube with the length of 4.5 mm and internal diameter of 3.5 mm. The end of custom metal tube was bent to a 45°labial angle, and another end was bonded to the mandibular incisors with glass sonomer cement (China) or light cure glass sonomer cement (Japan). For c group, the blue enamel resin (China) was stacked as the 45°labial guide plate on the lower incisors. Rats in each group were anesthetized with 2% pentobarbital sodium (45 mg/kg) and were bonded with MAA. To select the ideal MAA for rats, we compared the failure rates of different appliances. Based on the lowest failure rate of customized mental tubes glued with glass sonomer cement of 10%, we chose appliance a as the ideal MAA for rats.
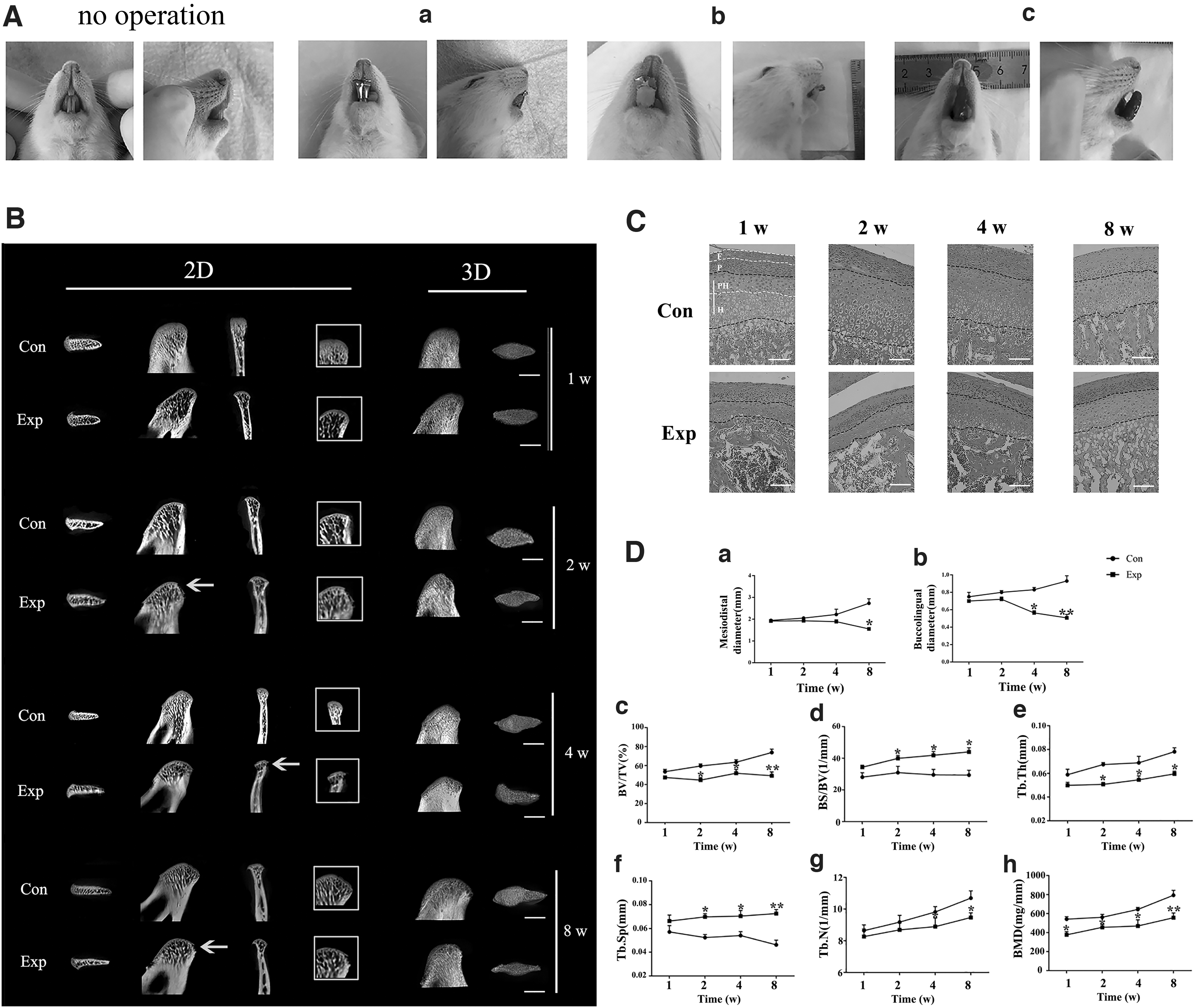
OOF induced condylar bone absorption and cartilage degradation.
In the formal experiment, 48 male SD rats were randomly assigned into experimental group and control group. After 1, 2, 4, and 8 weeks (n = 6), rats were killed, and the condyles were collected for tissue analyses. Throughout the experiment, rats with the devices falling off or that died were excluded from further analysis. The MAA affected the food intake; we crushed the food for rats to take in easier and standardized diet to synchronize the growth and development of rats. Therefore, the changes of condyles were attributed to OOF rather than the differences of growth and development.
Micro-CT analysis
After 1, 2, 4, and 8 weeks, the condyles of rats were dissected and fixed in 4% paraformaldehyde for micro-computed tomography (micro-CT) scanning and then were prepared for the nanodynamic mechanical analysis test.
Scan condition was set to 70 kV, the electrical flow is 60 μA, the exposure time was set to 4,000 ms, and the resolution was set to 18 μm. Mimics software was used for reconstruction. Two cubes (0.5 × 0.5 × 0.5 mm) were selected from the condyle tissue in a blinded manner to calculate the relevant bone parameters, including bone volume/tissue volume (BV/TV), bone surface/tissue volume (BS/TV), bone trabecular thickness (
Enzyme-linked immunosorbent assay
The levels of RANKL and OPG in condylar subchondral bone were assayed by rat RANKL Enzyme-Linked Immunosorbent Assay (ELISA) Kit and rat OPG ELISA Kit (Elabscience Biotechnology Company, China) based on the manufacturer's instructions. The optical density was measured by a microplate reader set to 450 nm with wavelength by a spectrophotometer.
Hematoxylin and eosin, immunohistochemistry staining, and immunofluorescence staining
The mandibular condyles of rats were dissected and fixed in 4% paraformaldehyde and then decalcified with 4% EDTA for 5 weeks, dehydrated in ethanol, embedded in paraffin, and cut sagittally into 5 μm-thick serial sections. The most central sagittal sections of each condylar were selected randomly for hematoxylin-eosin (HE) staining and immunohistochemical (IHC) detection with antibodies for RANKL (2 μg/mL, ab19347; Abcam) and OPG (2 μg/mL, ab7150; Abcam). For control group, nonimmune goat serum was substituted for the primary antibody. For immunofluorescence (IF) staining, sections of 8-week samples were incubated with antibodies of Osterix (μg/mL, ab22552; Abcam) and Trap (μg/mL, abab2721; Abcam). Sections of samples of rats for 4 time points were incubated with antibodies of Osterix and mTOR (μg/mL, 66888–1). Fluorescence intensity was evaluated using Image-Pro Plus 6.0 (Media Cybernetics, Rockville, MD).
Isolation and culture of rat BMSCs and the osteogenic induction
Bone mesenchymal stem cells (BMSCs) were harvested by flushing bone marrow cells from the femurs of male Sprague–Dawley rats. The bone cavity was flushed with complete cell culture medium, which comprised α-minimum essential medium (Hyclone, Beijing, China) supplemented with 1% penicillin/streptomycin (Hyclone) and 10% fetal bovine serum (Gibco, Shanghai, China). Cellular material was centrifuged, the supernatant was discarded, and the pellet was mixed with complete cell culture medium. Cells were plated into a culture flask and incubated in a humidified atmosphere containing 5% CO2 at 37°C. The medium was changed for fresh medium every 3 days. Cells were subcultured when cell growth reached 80% confluency. After that, the BMSCs were identified through flow cytometry detected by CD34, CD29, CD45, CD90 (Supplementary Fig. S1). Then, BMSCs were induced into OBs, cultured with osteogenic induction medium, containing 0.01% dexamethasone, 50 g/mL vitamin C, and 1% glutamine. The OBs were identified by Alizarin Red Staining and CCK8.
Application of mechanical stretch on cells in vivo
The cells were stretched as described in our previous study [23]. OBs were cultured on collagen coated six-well BioFlex plates at a density of 3 × 105 cells/well and randomly divided into three groups: stretched group, stretch+rapa group, and control group. OBs from stretch+rapa group were pretreated with 100 nM rapamycin (Selleck) for 24 h before stretch application. Mechanical deformation was induced by flexible strain cell, which was composed of vacuum manifold controlled by solenoid valve and controlled by computer time program. The computer-controlled flexible strain device delivered mechanical stretch for 6 s per cycle with the elongation of 20%. OBs from stretched group and stretch+rapa group were subjected to mechanical stretch for 24 h. Unstretched OBs grown on BioFlex plates under identical conditions were served as static controls.
Real-time polymerase chain reaction
For cell experiments, after stretch loading, the total RNA was isolated from OBs using the TRIzol. After extraction, RNA was used for synthesis of complementary DNA. The PCR results of samples were standardized to the GAPDH expression as an internal reference. The primers applied for the PCR amplifications were listed as follows: mTOR forward: ACACCGAATCAATCATCAG, reverse: TCCTCTTCATCAGCAAGT; OPG forward: AATGTGGAATAGATGTTACC, reverse: TCTACCAAGACACTAAGC; GAPDH forward: TGAAGGTCGGTGTGAACGG, reverse: GTGAGTGGAGTCATACTGGAA. Expression of target genes, relative to that of GAPDH, was calculated by the 2−▵▵Ct method.
Western blot
After mechanical stretch, the OBs were washed with ice-cold phosphate buffer saline (PBS) and lysed by the RIPA buffer. The cell lysate was centrifuged, and the supernatants were detached at 14,000 g and 4°C for 10 min. The bicinchoninic acid (BCA) protein assay was applied to certain the actual protein amount. Equivalent amounts of proteins were separated on sodium salt-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto a polyvinylidene fluoride (PVDF) membrane. The membranes were blocked with 5% skim milk for 1 h and incubated at 4°C overnight with the primary antibodies. Then, the membranes were incubated with a secondary antibody for 1 h and observed by electrochemical luminescence. Densitometry analysis was executed by ImageJ software to quantify the relative protein expressions. Antibodies used were p-mTOR (μg/mL, ab8440; Abcam), mTOR (μg/mL, ab2732; Abcam), RANKL (μg/mL, 239617; Abcam), and OPG (μg/mL, ab73400; Abcam).
Tartrate-resistant acid phosphatase staining (Trap Staining)
BMSCs were cultured and induced into the OCs by OC inducers (recombinant protein RANKL 50 ng/mL) to promote osteoclastic differentiation of BMSCs for 7 days. When BMSCs were induced into the late stage of OC differentiation, the loading experiment was repeated on OBs, and the supernate of three groups of OBs were collected and added to the culture medium of BMSCs in late differentiation. The cells were then cultured for 24 h. Subsequently, cells were collected and washed twice with PBS and lysed with 0.2% Triton-100. Then cells were reacted with TRAP at 37°C for 30 min. The reaction was terminated by adding 50 mL NaOH solution (0.1 N). Five sights of sections were randomly selected to calculate multinucleated cells. The typical mature OC owns giant cell, multiple nuclei, irregular shape, and Trap+.
Statistical analysis
Data are expressed as the mean ± standard deviation (SD) from at least three independent experiments. Differences between experimental groups were analyzed using the Student's t-test. Values of P < 0.05 are defined as statistically significant.
Results
OOF induced condylar bone absorption and cartilage degradation
Three-dimensional (3D) image, 2D image, and bone parameters of condylar subchondral bone were detected by micro-CT. Morphologically, the condylar head of experimental rats seemed slim and irregularly shaped, and the condyle surface looked rough and uneven, especially for the 8-week rats. As Fig. 1B showed, the condyle in experimental group for 2, 4, and 8 weeks exhibited the discontinuous bone cortex line, which was a crucial indicator of bone destruction in bones of trunk and limbs (yellow arrow). Notably, the discontinuous bone cortex line was reported for the first time on the mandibular condylar bone resorption research. In contrast, the condyles of control group exhibited the plump shape, smooth surface, and continuous cortical lines of bone (Fig. 1B). Normally, the length and width of condyle increased related to normal growth age dependently. However, under the OOF originating from MAA, the condylar volume was reduced obviously for 2 weeks, with the considerable reduction of both length and width, worsening at 8 weeks (Fig. 1D). BMD of the condyles increased gradually with time in both control group and experimental group. And the BMD of the mandibular condyles in experimental group was higher than the matched control group at 2, 4, and 8 weeks, confirming that the OOF caused osseous destruction of condyles from the early stages. Consistent with the results of BMD, a significant loss occurred in the subchondral bone of mandibular condyles in experimental group. As indicated in Fig. 1C, BV/TV in experimental group was markedly lower than that in control group, respectively, especially at 2, 4, and 8 weeks. In comparison to the control group, the decreased trabecular number (Tb.N) and trabecular thickness (
Histologically, the condylar cartilage normally was composed of fibrous, proliferative, prehypertrophic, and hypertrophic layers and owned an abundant cartilage matrix. For experimental condylar cartilage, irregular cellular arrangements and marked cartilage loss in prehypertrophic and hypertrophic zones were visually detected, indicating the degradation of condylar cartilage induced by OOF (Fig. 1D).
OOF disturbed the OC-OB balance, leading to condylar bone absorption
Based on 8-week experiment rats representing the peak of condyle bone absorption under OOF, we selected 8-week samples as our research objectives.
To assess the OC and OB activity in condylar bone resorption, we chose the 8-week samples and labeled bone tissue section by double-labeling IF with Trap and Osterix, bonding specifically to OCs and OBs. As shown in Fig. 2A, positive areas and fluorescence intensity of Trap and Osterix in experimental rats were visually larger than those of control rats, indicating that both osteoclastogenesis and osteogenesis were enhanced in condyle bone absorption. To further compare the difference of OC activity and OB activity under OOF, we quantitatively analyzed the fluorescence intensity of Trap and Osterix. As shown in Fig. 2B, the OCs were activated prominently than OBs, illustrating the osteoclastogenesis prevailing over osteogenesis, which accounted for condylar bone absorption.
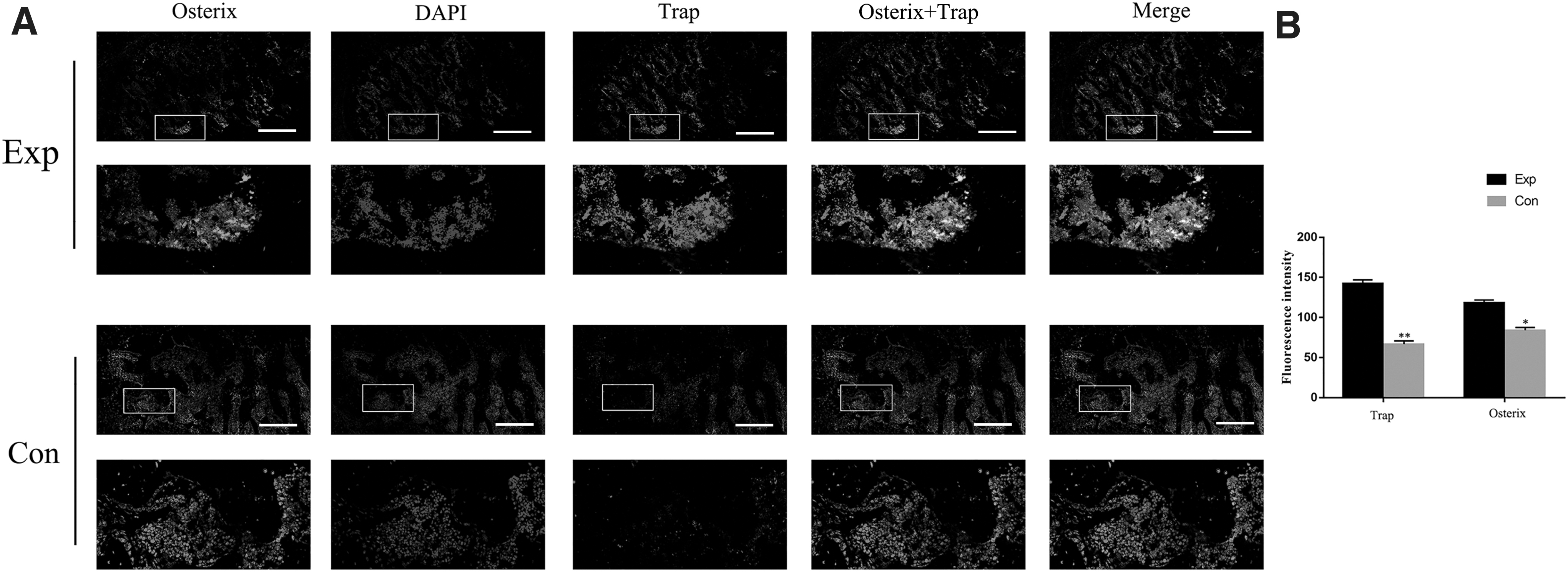
OOF activated osteoclastogenesis prevailing over osteogenesis.
The mTOR and RANKL/OPG of OBs were activated by OOF during condylar bone absorption
To verify the potential association of mTOR and OBs, we opted immunofluorescent labeling of the Osterix, mTOR, and 4,6-diamino-2-phenylindole (DAPI). As shown in Fig. 3A, under OOF, the expression of mTOR was increased in condylar subchondral bone of experimental rats compared with that of control rat. The OBs could be labeled by Osterix and DAPI dye. More importantly, it is observed that mTOR-positive areas and OB area in experiment mice condyles partially overlapped, implying that the mTOR of OBs in condylar subchondral bone was activated by OOF.
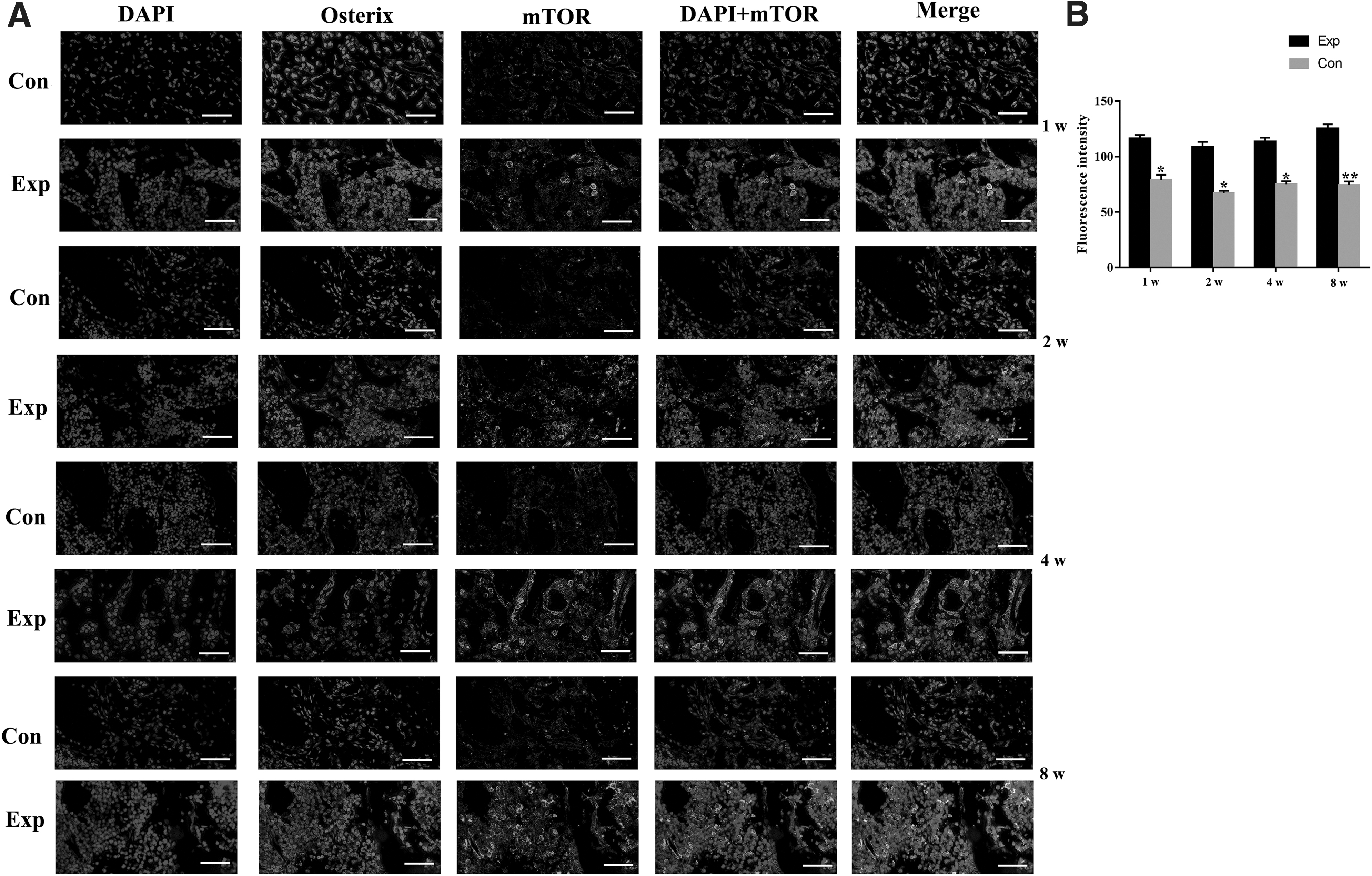
OOF activated the mTOR in osteoblasts in subchondral bone.
The results of IHC staining indicated that the expression of RANKL was significantly increased in the experimental group compared with those in control group over 1 week (Fig. 4A, B). The ratio of RANKL/OPG was effectively increased, indicating the activation of RANKL/OPG. In addition, ELISA was performed to measure the concentrations of RANKL and OPG in condylar subchondral bone collected from different rat groups. Compared with the control rats, OOF obviously increased the levels of RANKL at 1, 2, 4, and 8 weeks. However, the stimulation of OOF exerted a milder effect on OPG expression in experimental rats, without increasing the OPG level to a large extent. The ratio of RANKL/OPG was significantly elevated (Fig. 4C).

OOF altered the RANKL/OPG ratio in subchondral bone.
Overloaded mechanical stretch altered the expression of mTOR, RANKL/OPG ratio in OBs in vitro, and the supernatant of OBs promoted MSCs differentiating into OCs
We further detected the association between mTOR and RANKL/OPG in vitro. First, we applied the overloaded stretch on OBs to simulate the force microenvironment originating from MAAs. Mechanical stretch remarkably increased the mRNA levels of RANKL and mTOR. The ratio of RANKL/OPG was increased significantly. Meanwhile, stretch loading obviously increased phosphorylation of mTOR, without altering the overall mTOR protein levels, indicating that mechanical stretch indeed activated protein of p-mTOR (Fig. 5A). In addition, rapamycin effectively abrogated the rising trend of p-mTOR in OBs induced by mechanical stretch. After confirming that the activation of mTOR was suppressed successfully, we repeated the stretch loading experiments. As shown in Fig. 5B, the increased ratio of RANKL/OPG was effectively decreased by the inhibition of p-mTOR. The results of PCR exhibited the similar trend. As observed before, the activation of mTOR increased ratio of RANKL/OPG genetically and in protein.
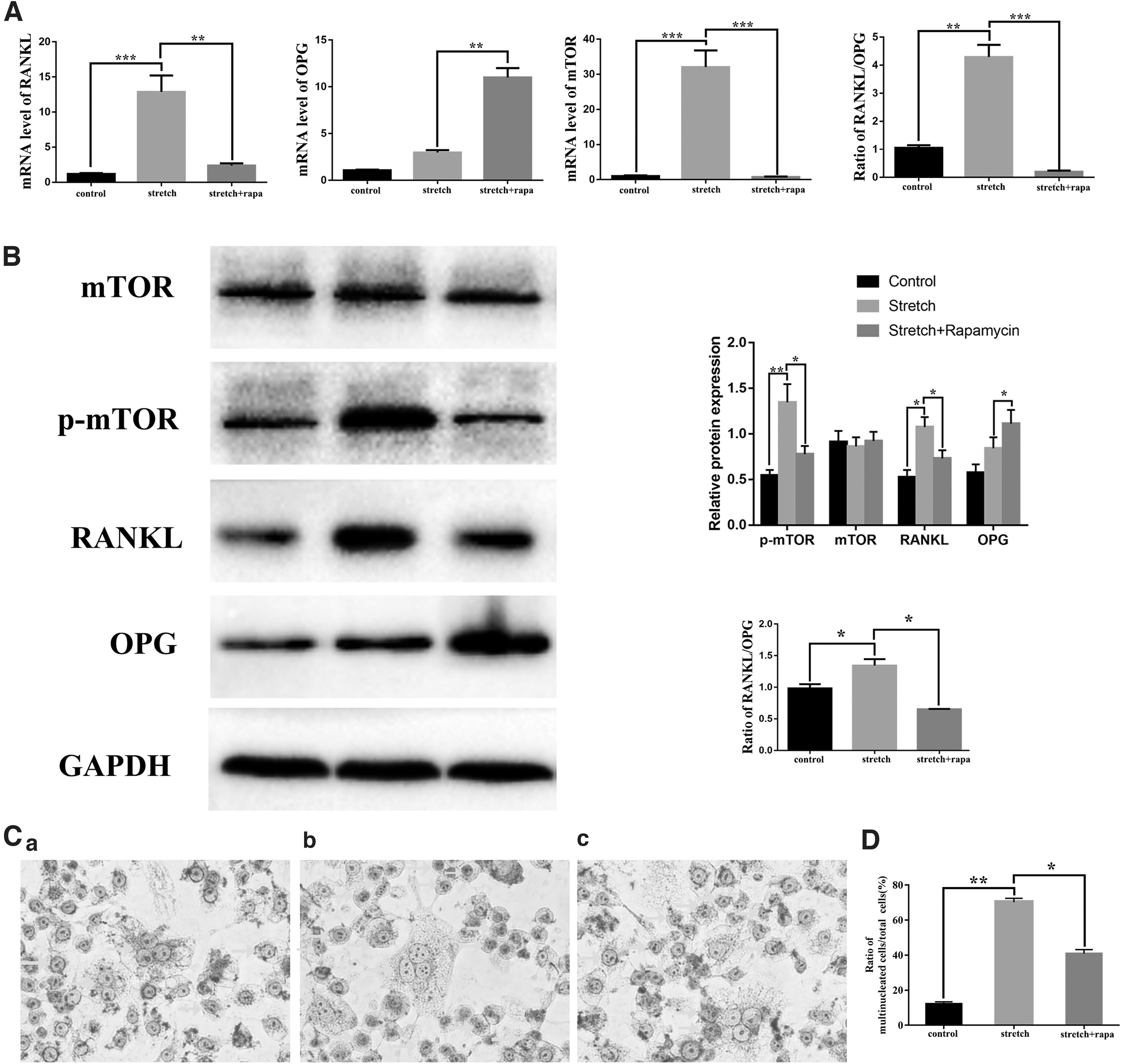
Overloaded mechanical stretch altered the expression of mTOR, RANKL/OPG ratio in osteoblasts, and the supernate promoted osteoclastic differentiation of MSCs.
To get more insights into the potential effects of mTOR of OBs on OC differentiation, we collected the supernate from different OBs with or without mechanical stretch and then added it to the culture medium of OCs differentiating in the late stage. The differentiation levels of OC were confirmed by TRAP staining. Typical mature OC appeared multinucleated, and its shape was irregular and spiny. As shown in Fig. 5C, the culture supernate of stretched OBs enhanced the OC differentiation of MSCs and increased the ratio of TRAP-positive mature OCs to total cells compared with the blank group (Fig. 5D). Moreover, the culture supernate from stretched OBs pretreated with rapamycin reversed the increased osteoclastogenesis.
Taken together, data from this part of our study clearly suggested that the activation of mTOR in OBs induced by overloaded orthopedic stretch increased the ratio of RANKL/OPG. The function of OB on osteoclastogenesis of MSCs was realized through mTOR of OBs by RANKL/OPG secretion under OOF.
Discussion
The mandibular condyle is mainly constitutive of overlying cartilage and the underlying subchondral bone. Previous researches about mandibular condyle were mainly focused on the condylar cartilage rather than subchondral bone [24,25]. Actually, in the state of loading, the mechanical force was transmitted into condyle cartilage and subchondral bone; ∼30% of it was effectively absorbed by subchondral bone, indicating the pivotal role of subchondral bone in maintaining bone homeostasis and alleviating bone destruction [26]. It's worth noting that, different from other arthrosis, mandibular condyle has a special multidirectional capacity of growth and remodeling and then adaptive to the mechanical force or positional changes, which is one of the bases of functional orthopedic treatment [5]. Excessive advancement of mandibular may lead to iatrogenic temporomandibular joint disease [6]. Actually, the effects of OOF exerted on subchondral bone of condyle and its molecular mechanism about osteogenesis and osteoclastogenesis have not been elucidated.
In this study, we established the model of mandibular advancement in vivo and in vitro to investigate the molecular mechanism of condyle subchondral bone using HE staining (HE staining), Micro-CT, IHC staining, ELISA, IF labeling, western blot, and PCR.
In vivo, we first established OOF model in rats through the ideal MAA at different time points. The anatomical changes of condyles on rats were detected by micro-CT. As the ultrastructural results were in accordance with expectations, OOF induced the condylar bone absorption of rats, representing the model's successful construction as well. There was no obvious difference that occurred after 1 week loading in both morphology and bone parameter analysis. From 2 to 8 weeks, the mandibular condylar exhibited the discontinuous cortical lines, unsmooth surface, and narrow and irregular shape. Notably, discontinuous cortical lines of mandibular condyle were first reported in animal experiments, which is the crucial diagnostic criteria of condylar absorption clinically. The rough appearance of condyles indeed increased the surface area. In fact, additional surface area would improve load transmission across the mandibular condyle, for bone is structurally prone to absorbing mechanical forces over a larger cross-sectional area than a small one [27]. Consequently, it could be speculated that under OOF, the weakened subchondral cortical plate and trabeculae led to deformation of the condyle surfaces, consequently resulting in the subchondral osteoporosis. In addition, Felson and coworkers discovered that compared with normal joint, the subchondral bone from bone absorption was less stiff and dense, with a greater porosity, reduced mineral content, and lower biomechanical property [28]. The similar changes in condyle in the early stages made it vulnerable to mechanical stimulation, accelerating bone resorption in advanced stages in turn. Moreover, with the onset and progression of condylar absorption, the condyle was characterized as the decreased BMD, BV/TV ratio, Tb.Th, and increased Tb.Sp, and the difference peaked at 8 weeks.
Bone is in dynamic balance of OCs and OBs throughout life. OB and OC are both the sensory cells of mechanical signals and the effector cells of stress stimulation, which play a central regulatory role in the process of bone metabolism [29]. In an activation bone remodeling sequence, each Basic Multicellular Unit (BMU) turns over a small packet of bone, where bone resorption of OCs and formation drifts of OBs could change bone surfaces and volume in tissue space to thicken, shape, and strengthen trabeculae [29]. When beyond the physiological range of force, bone formation of bone remodeling failed to compensate for bone loss, and bone adsorption occurred ultimately. In 8-week experiment group, both OBs and OCs were effectively activated, while OC differentiation was prominent, raising the OC/OB ratio. H.M. Frost stated that compared with those in subchondral cortex bone, BMUs existing near bone marrow make less changes of bone, and osseous may be a master risk factor of the bone marrow lesions in knee pain [3,28]. In our study, similar observations that the active fluorescence of OCs and OBs was mainly located in the deep regions under the subchondral cortical bone, next to bone marrow, were consistent with previous studies [4].
In addition, as shown in Fig. 3, the mTOR-positive areas and Osterix-positive areas overlapped, while part of mTOR-positive areas also occurred in Osterix-negative areas. Besides OBs, bone cells contain OCs, osteocytes, bone lining cell, and so on, which could not be labeled with Osterix. Actually, we could not rule out the other bone cells and defined that mTOR was selectively activated in OBs under OOF. The activation of mTOR may be involved in other bone cells, which need further investigation.
RANKL/OPG was recognized as the pivotal pathway implicated in the OC differentiation. In vivo, we observed the increased mTOR, RANKL/OPG ratio, and the enhanced osteoclastogenesis of subchondral bone under OOF. In vitro, mTOR in OBs was elevated significantly by overloaded mechanical stretch, as well as RANKL/OPG ratio. In vitro, under overloaded mechanical stretch, mTOR in OCs was activated and involved in promoting the OC differentiation through RANKL/OPG secretion.
Extracellular vesicles (EVs), including small exosomes and microvesicles, are able to shuttle biologically active molecules, like proteins, lipids, mRNAs, and miRNAs, by many mechanisms of cellular interaction, including juxtacrine signaling, ligand/receptor interaction, protein shedding, and cargo internalization [30 –32]. EVs can shuttle both ligands and receptors, thus activating new signaling pathways between cell communication. Alfredo and his colleagues discovered that the EVs originating from OBs were shuttled to transport RANKL and modulate osteoclastogenesis. In vitro, the supernatant of stretched OBs significantly promoted OC differentiation of MSCs. Actually, the constituents of supernatant were extraordinarily complex. The unknown pathways may be involved in the mTOR and RANKL/OPG of OBs in upstream or downstream in regulating osteoclastogenesis of MSCs under OOF. More actions are needed for the detailed mechanism research like gene chip technology.
In summary, our study proposed that OOF induced condylar bone absorption of rats, in which osteoclastogenesis was prominent than osteogenesis. The mTOR derived from OBs under OOF could increase OC differentiation through RANKL/OPG.
Footnotes
Acknowledgment
The authors wish to thank the reviewers for their valuable comments and suggested improvement to the article.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This study was supported by the National Natural Science Foundation of China (31870929) and Natural Science Foundation of Shandong (ZR2019MH007).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.