
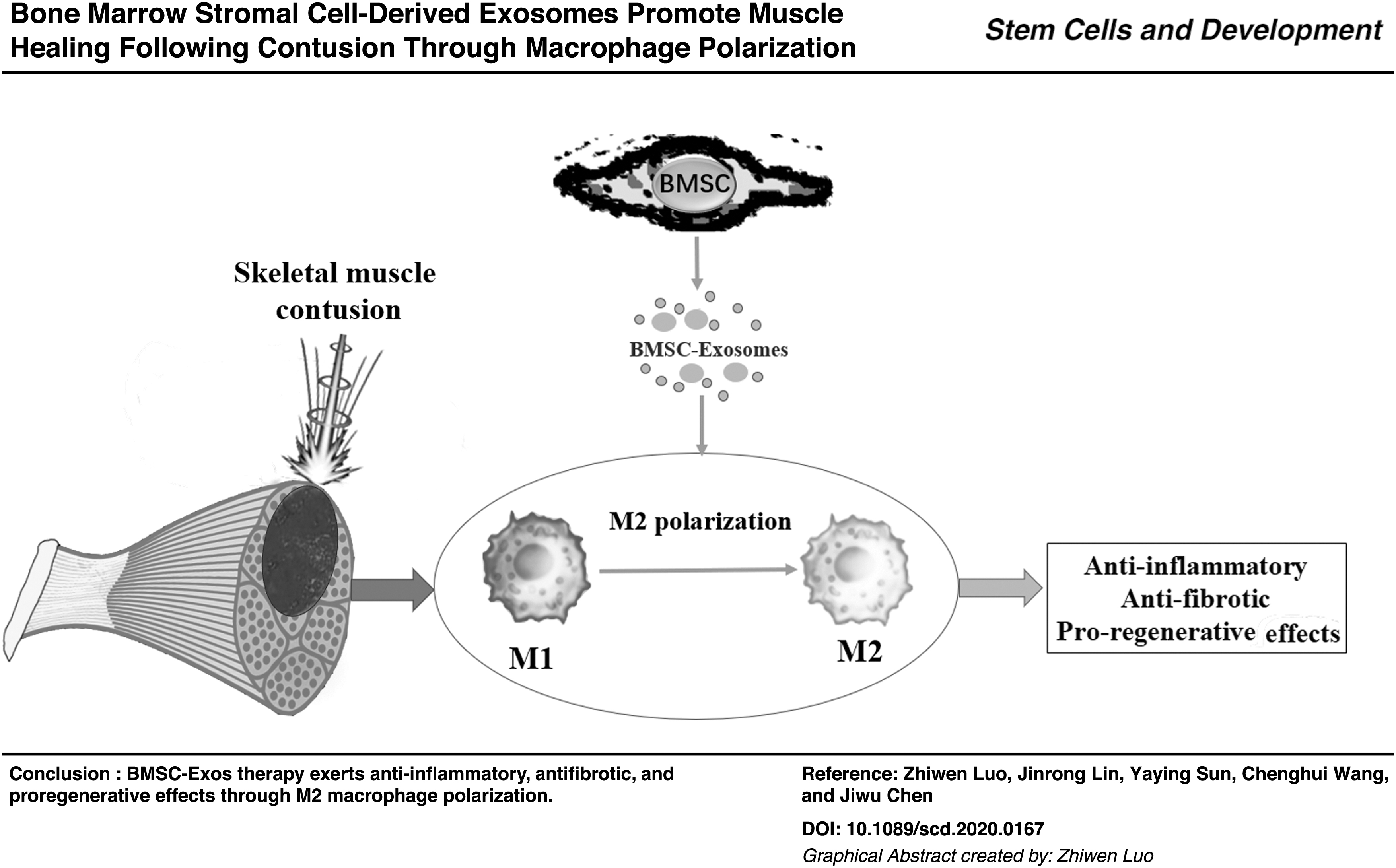
Abstract
Skeletal muscle contusion is among the most common injuries in traumatology and clinics of sports medicine. The injured muscle is vulnerable to re-injury owing to fibrosis formation. Given that the bone marrow stromal cell-derived exosomes (BMSC-Exos) displayed promising therapeutic effect for various tissues, we used BMSC-Exos to treat skeletal muscle contusion and investigated its effects on muscle healing. In this study, the in vivo model of skeletal muscle contusion was established by subjecting the tibialis anterior of young male mice to hit injury, and the in vitro inflammation model was established by lipopolysaccharide treatment on macrophages. Macrophage depletion model was built by intraperitoneal injection with clodronate-containing liposomes. Exosomes were isolated and purified from the supernatant of BMSCs using gradient centrifugation. Nanoparticle tracking analysis, transmission electron microscope, and western blot were used to identify the exosomes. HE stain, Masson stain, immunofluorescence, and biomechanical testing were carried out on the muscle tissue. In addition, enzyme-linked immunosorbent assay (ELISA) assays, real-time qPCR, flow cytometry, and PKH67 fluorescence trace were conducted in vitro. Intramuscular injection of BMSC-Exos to mice after muscle contusion alleviated inflammation level, reduced fibrosis size, promoted muscle regeneration, and improved biomechanical property. After macrophages depletion, the effects of BMSC-Exos were inhibited. In vitro, PKH-67 fluorescence was internalized into macrophages. BMSC-Exos promoted M2 macrophages polarization both in vivo and in vitro. At the same time, BMSC-Exos reduced the production of inflammatory cytokines under the inflammatory microenvironment and upregulated anti-inflammatory factors expression. In conclusion, BMSC-Exos attenuated muscle contusion injury and promoted muscle healing in mice by modifying the polarization status of macrophages and suppressing the inflammatory reaction.
Introduction
Skeletal muscle contusion is among the most common injuries in traumatology and clinics of sports medicine [1]. Because of the local fibrosis formation after contusion, the injured muscle is vulnerable to re-injury [2]. Skeletal muscle fibrosis was proved to be associated with excessive inflammatory reaction in the early stages of injury [3,4]. The macrophage, closely related to the inflammatory infiltration, was found to play a critical role during the skeletal muscle healing [1,5,6].
In general, acute muscle damage initially recruits “classically activated” macrophages (M1) into the injury area, which promotes further muscle damage and satellite cell activation by generating nitric oxide (NO) and proinflammatory cytokines [7]. Next, “alternatively activated” M2 macrophages substitute the M1 phenotype to promote muscle regeneration and differentiation [8]. But in normal conditions, the M2 polarization is typically incomplete and suppressed after acute muscle contusion [9,10]. Consequently, how to regulate macrophage polarization remains an essential issue for muscle healing.
Exosomes, as an essential member of cellular vesicles (40–150 nm diameter, lipid bilayer membrane), are secreted by many kinds of living cells and can be transported all around the body through the blood [11]. Moreover, exosomes are wrapped with a variety of nucleic acids, proteins, and other bioactive substances from donor cells. When they contact with target cells, biological molecules carried by exosomes will be received to exert their functions through endocytosis [12]. Recent studies have shown that bone marrow stromal cell-derived exosomes (BMSC-Exos) can affect inflammation and tissue recovery [13 –15]. Therefore, we hypothesized that the BMSC-Exos could downregulate inflammation by influencing macrophage polarization and thus promote the healing of skeletal muscle after contusion.
In this study, we isolated BMSC-Exos from BMSCs, tested their functions both in vivo and in vitro, and investigated the potential underlying mechanisms of the effects of BMSC-Exos on immunomodulation of macrophages and muscle healing.
Materials and Methods
Cell culture
RAW 264.7 (mouse leukemia cells of monocyte macrophage) were purchased from the American Type Culture Collection. Cells were maintained in high glucose Dulbecco's modified Eagle's medium (DMEM) (HyClone) with 10% fetal bovine serum (FBS) and 0.5 mL of penicillin/streptomycin solution (#0503; ScienCell Research Laboratories) in a humidified incubator at 37°C and 5% CO2 atmosphere.
Human bone marrow stem cells (hBMSCs) were purchased from ScienCell Research Laboratories. Cells were maintained in complete mesenchymal stem cell medium (ScienCell Research Laboratories) in a humidified incubator at 37°C and 5% CO2 atmosphere.
Isolation and Identification of BMSC-Exos
BMSC-Exos extraction procedures were according to the previous method [16]. Fifty-milliliter conditioned culture medium containing 5 mL exosome-free FBS, 0.5 mL of mesenchymal stem cell growth supplement, and 0.5 mL of penicillin/streptomycin solution were used to culture BMSCs for 48 h. After the BMSCs grew over 90% confluence, cells were kept quiescent for 12 h, and then we collected all the supernatant. After this step, the media were subjected to sequential centrifugation (Optima XPN-100 ultracentrifuge; Beckman Coulter SW 41 Ti rotor) at 10,000 g for 35 min to remove the cell debris and then at 100,000 g for 70 min and washed in PBS twice under the condition of 100,000 g for 70 min. The BMSC-Exos were resuspended in PBS and stored at −80°C before use.
To learn the absolute size distribution of BMSC-Exos, exosomes were analyzed using a NanoSight NS300 (Malvern). With nanoparticle tracking analysis (NTA), the particles were automatically tracked and sized based on Brownian motion and the diffusion coefficient.
To examine the morphology of exosomes, the BMSC-Exos were observed directly under a transmission electron microscope (TEM, Tecnai G2 Spirit; Tecnai).
In addition, the BMSC-Exos were identified by western blotting with anti-CD63, anti-CD9, and anti-TSG101 antibodies (purchased from Abcam).
Animal model and surgical procedure
All animal experiments were conducted according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The experimental protocols were approved by the Animal Care Committee of Fudan University (approval no.20171248A703). All efforts were performed to minimize animal suffering.
A total of 60 male mice (10–11 weeks) used in this study were obtained from Shanghai Experimental Animal Center. Twelve of them were randomly selected into macrophages depletion group, six received BMSC-Exos treatment, whereas the others did not. All experiments were performed with acute muscle contusion on the right tibialis anterior (TA), as described in our previous study [2], and the left TA served as the control group. In brief, after anesthetization by intraperitoneal injection with 1% pentobarbital sodium (0.5 mL/100 g), the right hindlimb of the mouse was fixed on a plate with the exposed TA. A 15 g–2 cm stainless steel ball was released at the height of 1 m as an impactor on TA. Then the mice were randomly assigned into two groups: phosphate-buffered saline (PBS) injection (PBS group; n = 24) and BMSC exosomes injection (BMSC group; n = 24). In the BMSC group, 20 μL BMSC exosomes (equal to 1 × 1010 per mouse) were, respectively, injected into the proximal and distal end of the TA muscle on day 1, whereas the same volume of PBS was injected in the PBS group. At 3 and 7 days, six mice in each group were evaluated for macrophage polarization and muscle regeneration. Muscle functional recovery and fibrosis were analyzed at 14 and 28 days.
Macrophages depletion
We established the macrophages depletion model using a published method [1]. In general, 2 days before contusion, the mice were intraperitoneally injected with 2 mg clodronate-containing liposomes (purchased at Liposoma Research Liposomes). Then 0.5 mg of clodronate-containing liposomes were re-injected on days 0, 3, 6, 9, and 12 after muscle contusion to maintain macrophage at a low level. The spleen tissue of mice was used for flow cytometry to identify the ratio of lymphocytes (validated the macrophage depletion model). Muscle functional recovery and fibrosis were analyzed at 14 days for all the mice in macrophages depletion group.
Western blot
Protein was extracted and analyzed using an established method [17]. Anti-CD86, anti-CD206, anti-Arginase 1, anti-iNOS, anti-Myod1, anti-Myog, anti-beta-Tubulin, and anti-GAPDH antibodies were used as primary antibodies (Table 1). Each group contained 4 protein samples for calculation (n = 4/group).
Primary Antibodies Used in the Experiment
IF, immunofluorescence; W.B., western blot.
Histological analysis
After the mice were killed, the bilateral tibialis anterior were dissected and frozen in optimal cutting temperature with liquid nitrogen precooled and stored at −80°C until section. Then muscles were serially cryosectioned into 8 μm thickness for routine HE, Masson stain (for fibrosis analysis), and immunofluorescence stain for the interest proteins.
To examine the expression and location of target proteins, immunofluorescence staining on TA slices was conducted as previously described [18]. Primary antibodies used were anti-CD206, anti-α-SMA, anti-MyoD, and anti-Pax7. DAPI was used to locate nuclei. Images were observed by a fluorescence microscope (ECHO Revolve).
Biomechanical testing
The testing procedure for fast-twitch and tetanus strength was according to the established method [19,20]. In brief, TA muscles were harvested and kept in PBS until testing. The fast-twitch and tetanus strengths of 6 mice from each group (Contusion and BMSC-Exos groups) were examined 14 and 28 days after the injury to determine functional recovery using the strain gauge transducers coupled with a TBM4 strain gauge amplifier (World Precision Instruments, Inc., Sarasota, FL), which is recorded on a computer using a data acquisition program (Windaq; DATAQ Instruments, Inc., Akron, OH).
Exosome labeling with PKH67
Isolated BMSC-Exos were labeled with PKH67 [a Green Fluorescent Labeling Kit (Sigma, Aldrich, St. Louis, MO)], and the procedures were according to the company's protocol [21]. The BMSC-Exos or PBS was stained with PKH67 dye in 250 μL of Diluent C fluid for 5 min at room temperature. After that, 600 μL 1% FBS was used to stop the labeling process. They were then re-purified through ultracentrifugation with PBS rinse for 60 min (Fig. 1). The labeled PBS or BMSC-Exos were co-incubated with macrophages (M0) for 16 h in a 37°C, 5% CO2 cell incubator. After incubation, the culture medium was abandoned and washed three times with PBS. Cells were fixed with 4% paraformaldehyde and counterstained nuclei with DAPI for 3 min. The uptake of the labeled exosomes by macrophages was evaluated using the confocal microscope (SP8; Leica, Germany).
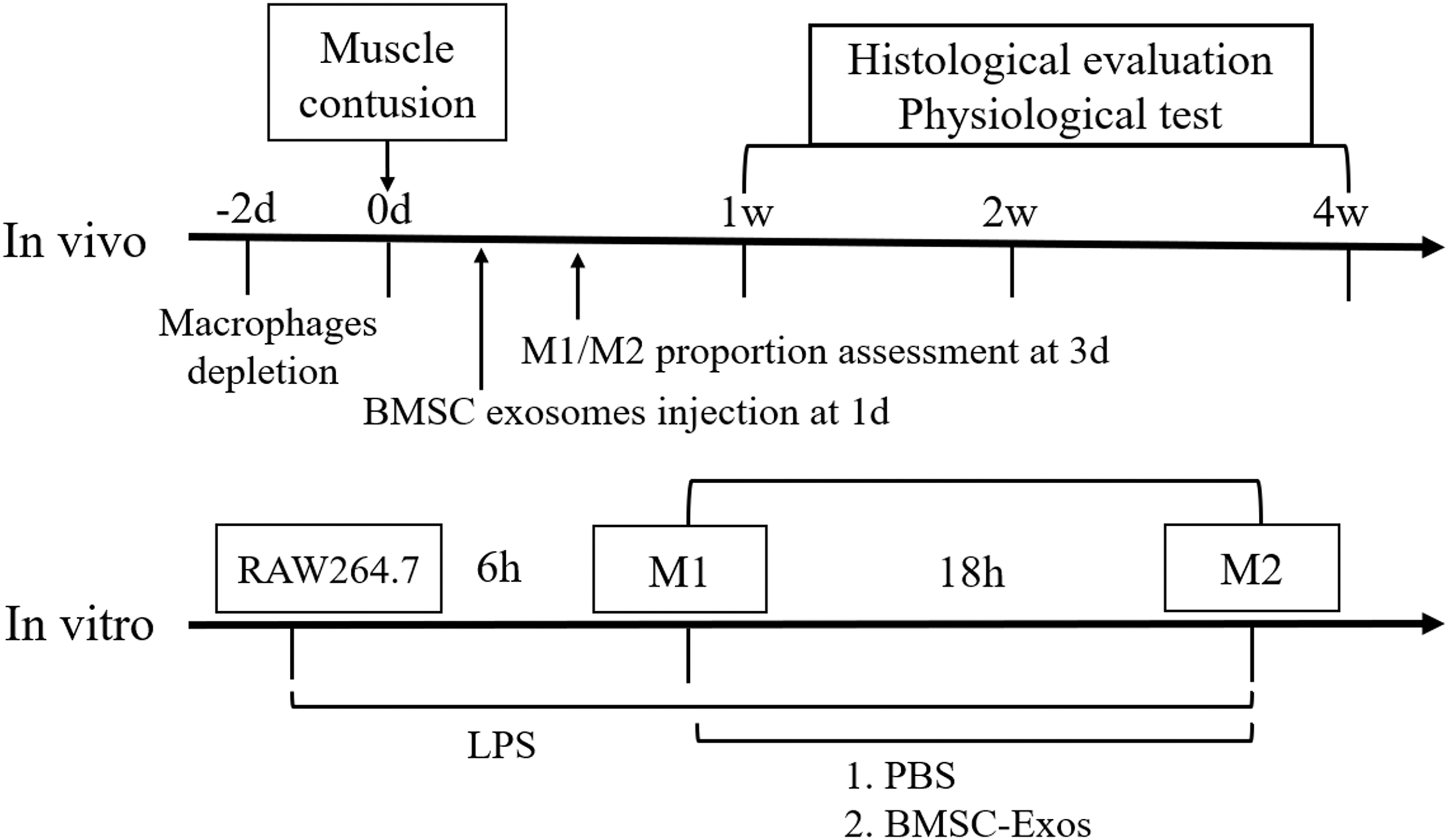
Experimental design for this study both in vitro and in vivo. BMSC-Exos, bone marrow stromal cell-derived exosomes; PBS, phosphate-buffered saline; M1/2, macrophage phenotype 1/2; LPS, lipopolysaccharide.
Cell intervention
RAW264.7 cells were incubated at 1 × 106 cells per well in a six-well plate and divided into two groups: LPS+PBS group and LPS+BMSC-Exo group. The cells of all three groups were polarized 500 ng/mL lipopolysaccharide (LPS) (M1 polarization). Six hours later, LPS was discarded and washed with PBS. The final concentration of BMSC-Exos in the culture medium reached 40 ng/mL in the LPS+BMSC-Exo group, whereas in the LPS+PBS group, an equal volume of PBS was added. After 18 h, all the cells were collected for further analysis.
Enzyme-linked immunosorbent assay
ELISA kits, including interleukin (IL)-6, IL-10, transforming growth factor β (TGF-β), and tumor necrosis factor α (TNF-α), were purchased from Laizee (LEM060-2, LEM100-2, LEM822-2, and LEM810-2). Cell supernatant of each group was collected, and then those kits were used according to the manufacturer's instructions.
Real-time qPCR
RNA was extracted and analyzed using the previous method [22]. Total RNA was obtained by the Trizol reagent (Invitrogen, Carlsbad, CA) and quantified by Nanodrop. RNA was then reversely transcribed by the PrimeScript RT reagent kit (Takara Bio). Specific primers used in the experiment are from PrimerBank (Table 2). The operation was performed on the ABI7900 Real-Time PCR System (Applied Biosystems). The expression of mRNAs relative to the expression of GAPDH was calculated and normalized to the control group.
Primers Used in the Experiment
Genes primers (5′-3′).
IL, Interleukin; TNF-α, tumor necrosis factor α; TGF-β, transforming growth factor β.
Flow cytometry for M1/M2
Flow cytometry was carried out using anti-CD86-PE, anti-CD206-APC, anti-CD11b-APC, anti-F4/80-FITC (Thermo/eBio). The percent of CD86+/CD206+/CD11b+ F4/80+ cell population (macrophage) was calculated using Cytomics™ FC 500 (Beckman Coulter).
Statistical analysis
All experiments were performed at least three times. Data were analyzed with GraphPad Prism 7.0 (GraphPad Software, La Jolla, CA) and were presented as mean ± SEM. Significance was typically analyzed by Student's t-test, one-way analysis of variance (ANOVA) followed by post hoc LSD test, and two-way ANOVA followed by multiple t-tests. P < 0.05 was regarded as significant.
Results
Identification and Characterization of BMSC-Exos
The NTA experiment measured the size of the BMSC-Exos. The diameters ranged from 40 to 150 nm, which were consistent with data from previous studies (Fig. 2B, Supplementary Fig. S1). Western blot results showed that exosomal marker proteins (CD9, CD63, and TSG101) were highly expressed in exosomes, whereas these proteins were expressed obviously in BMSC (Fig. 2C). Under transmission electron microscopy, BMSC-Exos presented a goblet or spherical shape (Fig. 2D).

Purification, isolation, and characterization of BMSC-Exos.
Macrophage depletion
Sixteen days after first clodronate-containing liposomes injection, the percent of CD11b+ F4/80+ cell population (macrophages) in mice spleen was calculated using flow cytometry. The data showed that macrophages were significantly reduced by 82.63% in the clodronate-containing liposomes treatment mice as compared with the control mice (Supplementary Fig. S5A, B).
Histological analysis
General appearance, relative weight, morphological characteristics
Three days after contusion injury, the TA muscle in the contusion group presented larger muscle hematoma than that in the BMSC-Exos group (Fig. 3A). The relative muscle weight of TA muscle was also significantly different on day 3 between the two groups, with the TA muscle in the contusion group being heavier (Fig. 3B).
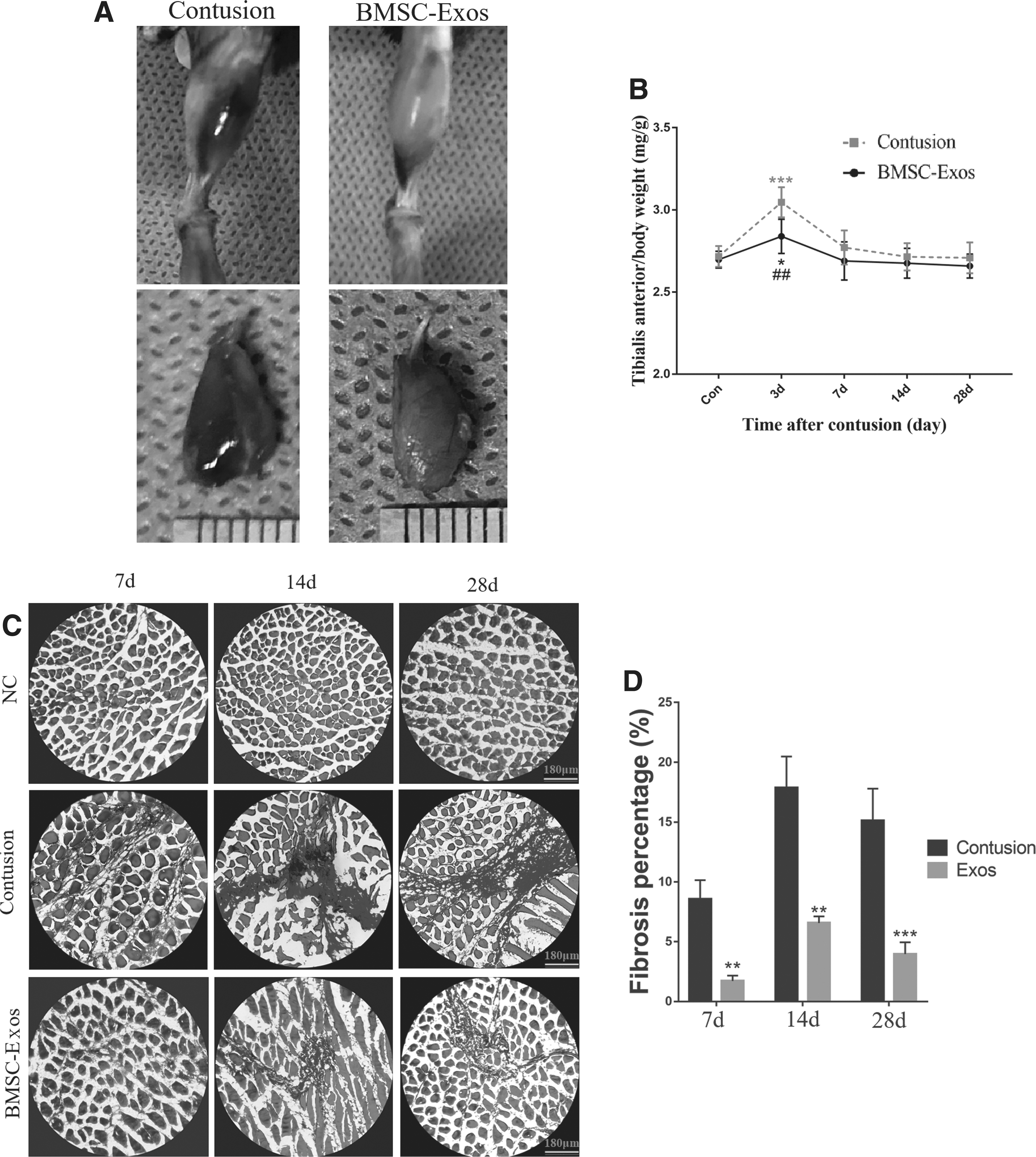
The effects of BMSC-Exos on general appearance, relative weight, and fibrosis of TA muscle.
As for HE stain, in the negative control group, the muscle fibers were arranged regularly and densely. However, in the contusion group, the muscle fibers became more and more scattered over time, and many proliferating collagen fibers occupied the spaces of muscle fibers. These phenomena were improved in the BMSC-Exos group, although there was still a small amount of fibrous tissue occupying the muscle space (Supplementary Fig. S4).
Fibrosis
To determine the fibrosis situations of different treatment groups at different time points, Masson staining was conducted. A large number of collagen fibers were observed in the contusion groups, whereas the BMSC-Exos therapy significantly reduced the fibrosis area at all time points (Fig. 3C, D). Moreover, to further examine the fibrosis condition, we performed the immunofluorescence stain for α smooth muscle actin (α-SMA). The morphological results showed that the BMSC-Exos therapy prominently reduced the high protein expression and wide distribution of α-SMA caused by contusion injury at different time points (Fig. 4). After macrophage depletion, the anti-fibrotic effects of BMSC-Exos inhibited large fibrosis area and wide distribution of α-SMA protein were observed in both contusion and BMSC-Exos treatment group at 14 days (Supplementary Fig. S5D).
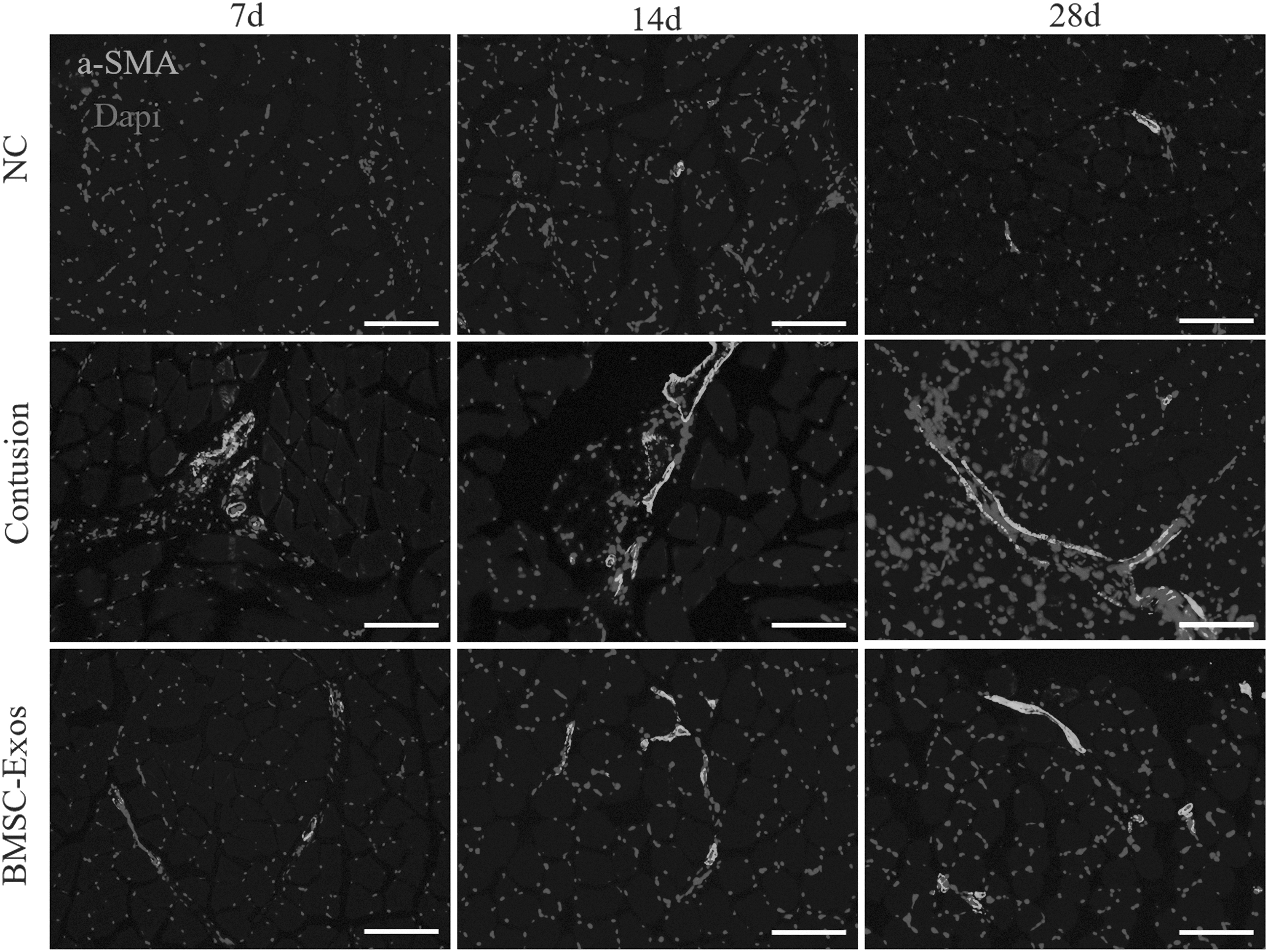
BMSC-Exos therapy reduced the expression of α-SMA. Immunofluorescence was used to detect relative expression and distribution of α-SMA among the three groups at 7, 14, and 28 days, including the standard control group, the contusion group, and the BMSC-Exos group (contusion + BMSC-Exos injection). 100 × magnification. α-SMA, α smooth muscle actin.
BMSC-Exos promoted muscle regeneration at an early stage
To identify muscle regeneration of different treatment groups, myogenic marker proteins (Myod and Myog) were measured. The western blot results showed that BMSC-Exos therapy significantly upregulated both regenerative proteins at day 3, compared with the contusion group. In contrast, at day 7, there was no significant increase between the two groups (Fig. 5A–C, Supplementary Fig. S2).

BMSC-Exos therapy increased the expression of muscle regeneration maker.
To determine the different myogenic status of satellite cells, the colocalization of Pax7 and MyoD was performed. Morphologically, total Pax7+/MyoD+ satellite cells prominently increased from 3 days after injury. Obviously, the quantity of the BMSC-Exos treatment group was more than that of the contusion group and negative control group. The relative Myod protein expression in the three different groups also presented a mimic tendency (Fig. 5D). To sum up, the BMSC-Exos injection promoted muscle regeneration at an early stage.
Biomechanical analysis
Fast-twitch and tetanus strength were significantly different between the contusion group and the BMSC-Exos groups at 14 and 28 days. Fast-twitch and tetanic strength in the negative control group and the groups treated with the BMSC-Exos presented a considerable difference on day 14, which disappeared at day 28 (Fig. 6A, B). But after macrophages depletion, both contusion and BMSC-Exos groups presented a significant decrease in tetanus strength compared with negative control group, whereas no difference existed between contusion group and BMSC-Exos group (Supplementary Fig. S5C). Overall, the BMSC-Exos injection improved muscle strength after contusion injury.
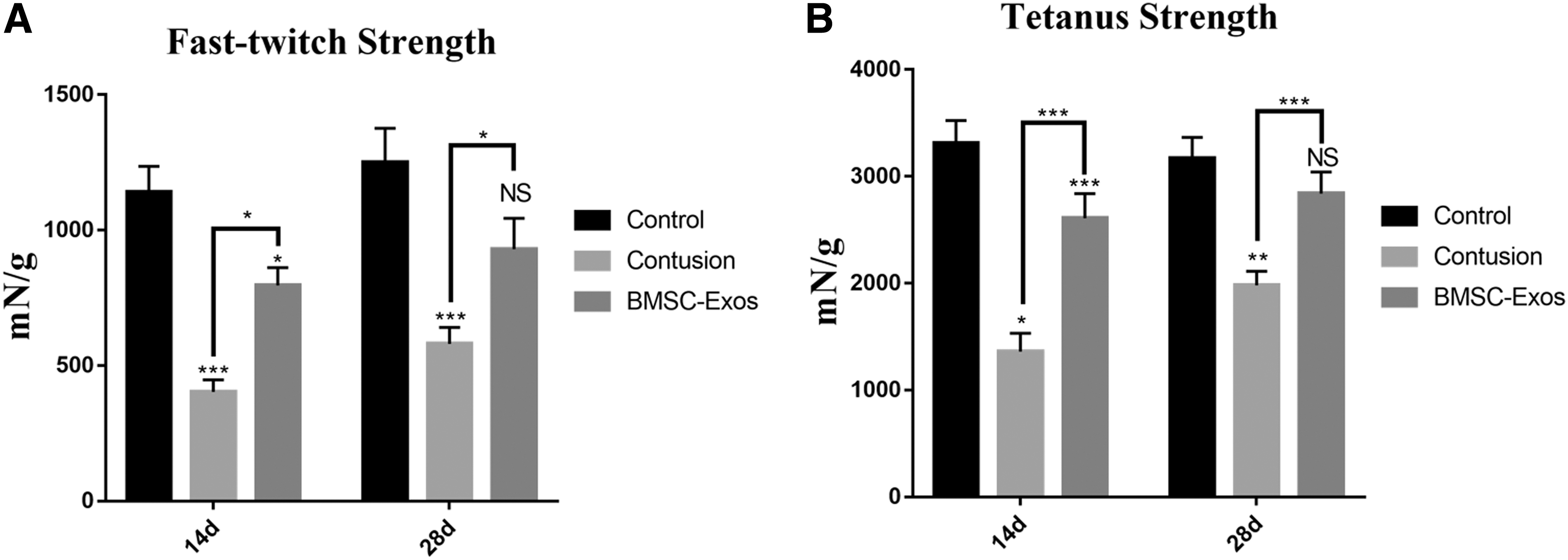
Physiologic analysis of the effect of BMSC-Exos therapy on muscle strength.
BMSC-Exos induced M2 macrophage polarization in vivo
To further examine the effects of BMSC-Exos on macrophages, BMSC-Exos were injected into the TA muscles of mouse 1 day after contusion. The similar results of BMSC-Exos were shown in vivo that BMSC-Exos significantly reduced the M1 marker protein expression (iNOS and CD86) and upregulated that of the M2 marker (CD206 and Arg1) (Fig. 7A–F, Supplementary Fig. S3). The immunofluorescence pictures have also presented that CD206+ cells localized relatively more extensive in the BMSC-Exos group than that in the contusion group on day 3 (Fig. 7G). Taken together, these data revealed that BMSC-Exos promoted the polarization of macrophages to M2 rather than M1 in a contusion mouse model.
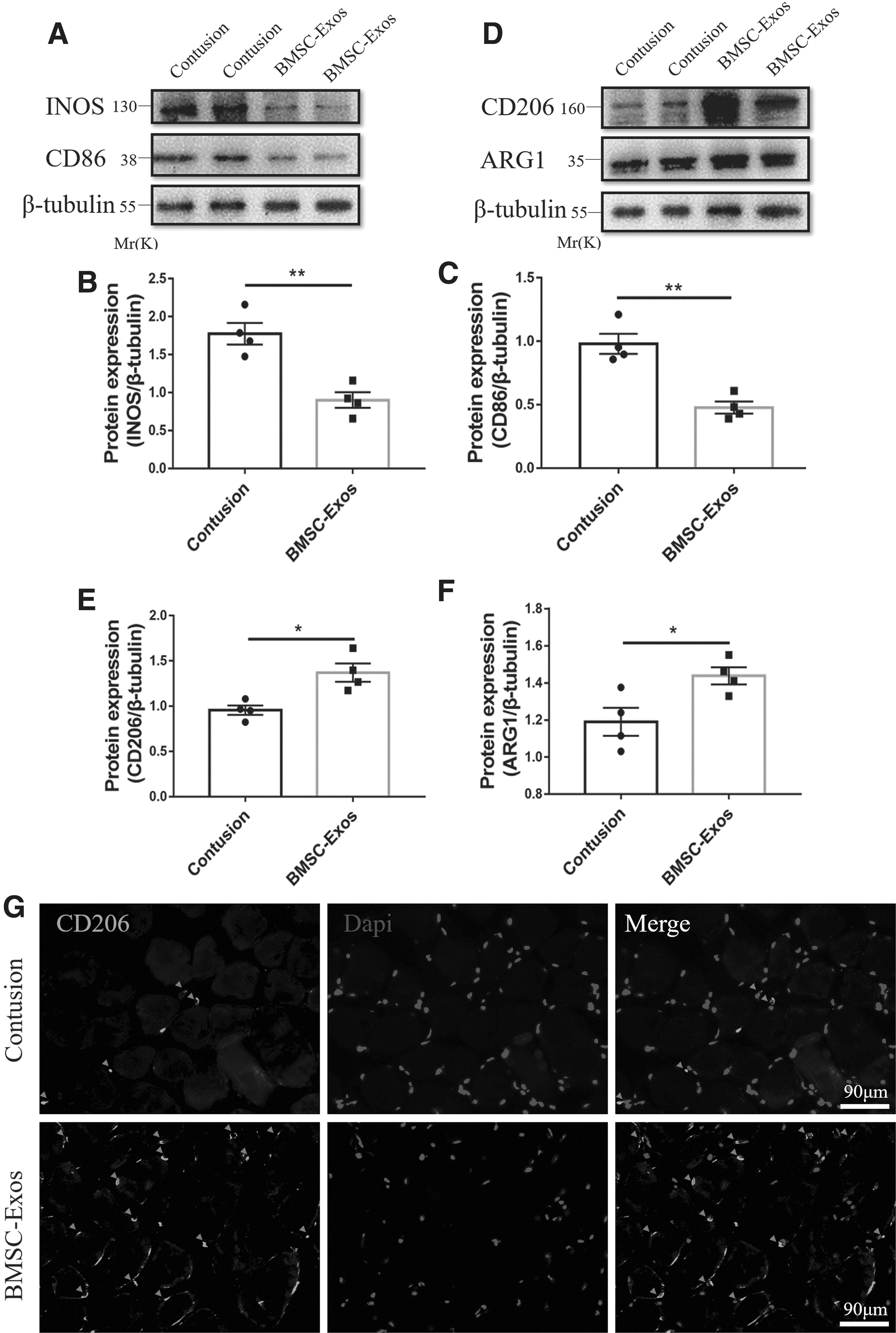
BMSC-Exos induced M2 macrophage polarization in vivo.
BMSC-Exos induced M2 macrophage polarization in vitro
To investigate how BMSC-Exos interact with macrophages, BMSC-Exos were applied to the cultured RAW264.7 cells. The PKH-67 (green)-labeled BMSC-Exos were internalized into macrophages and presented in the cytoplasm after 16 h of coculture (Fig. 8A). To further examine the effects of BMSC-Exos on macrophages, LPS was administrated to the culture system to induce an inflammatory microenvironment before the BMSC-Exos treatment. Six hours later, either PBS or BMSC-Exos was added to the stimulated-macrophages culture medium. Eighteen hours later, we measured the concentrations of cytokine IL-6, TGF-β, TNF-α, and IL-10 in the culture supernatants. We found that the BMSC-Exo effectively inhibited the LPS-induced IL-6 and TNF-α production and upregulated IL-10 and TGF-β (Fig. 8B). Moreover, flow cytometry results showed that the ratio of M1 macrophages significantly decreased, whereas that of M2 macrophages markedly increased in the BMSC-Exos group (Fig. 8C, D). Furthermore, the qRT-PCR analysis also demonstrated that after BMSC-Exos treatment, macrophages dramatically expressed more M2 markers (TGF-β, IL-10, Arg1, and CD206) and less M1 markers, including IL-6, TNF-α, IL-1β, and iNOS (Fig. 8E, F). Above all, these data demonstrated that BMSC-Exos induced the polarization of macrophages to M2 under the inflammatory environment in vitro.

BMSC-Exo induced M2 macrophage polarization under the inflammatory environment in vivo.
Discussion
Through BMSC-Exos treatment at the cellular level and in the contusion mice model, the M1 macrophages were prominently polarized to M2 phenotypes, which relived the inflammation, reduced the fibrosis scar, enhanced the muscle regeneration, and improved biomechanical properties after acute muscle damage. To the best of our knowledge, this is the first study demonstrating that BMSC-Exos attenuated muscle injury and promoted muscle healing through macrophage polarization.
Skeletal muscle injury is prevalent in clinical practice [1]. There is an inflammatory phase in the repair process after muscle injury [23]. Some studies have proved that inflammation may lead to injured skeletal muscle fibrosis [3,4]. Fibrosis tissue weakens the muscle biomechanical properties, which leads to second muscle injury [24,25]. Therefore, how to promote skeletal muscle regeneration and inhibit fibrosis is the research difficulty of promoting skeletal muscle healing. The application of stem cells in tissue injury has been widely studied [20,26 –28]. However, potential tumorigenicity and ethical problems hindered its clinical application [29 –31]. In recent years, researches have shown that the paracrine effects of stem cells play an essential role in the tissue regeneration process [32]. For example, several studies have found that the conditioned medium of stem cells could accelerate graft-bone incorporation [18,33]. Besides, many studies found that exosomes from many kinds of stem cells show similar effects on tissue regeneration, especially MSC-Exos [14,34 –37].
In this study, we found that BMSC-Exos could promote muscle healing and suppress fibrosis. Our findings showed that BMSC-Exos could promote regeneration markers (PAX7, MyoD, and MyoG) expression at an early stage (3 days). But the protein levels of MyoD and MyoG on day 7 did not change obviously. Iyer et al. found similar results with ours that BMSC-Exos therapy did not increase the mRNA level of MyoG and MyoD 10 days after muscle injury [38]. We suspected that the BMSC-Exos treatment indeed promoted the muscle regeneration process at 3 days, while at day 7, regenerative markers also achieved a peak in the contusion-only group. At 14 and 28 days, the fibrosis percentages in the BMSC-Exos group were significantly lower than those in the PBS group. In addition, the α-SMA relative protein expression showed the same downregulated trend after BMSC-Exos treatment. α-SMA is commonly regarded as a biomarker for a subset of activated fibrogenic cells, and its expression level can reflect the process of fibrosis [39]. This finding agrees with a recent study that reported exosomes containing miR-29 can reduce α-SMA and fibrosis levels [40]. Taken together, our results demonstrated that BMSC-Exos therapy potentially reduced the fibrosis level through downregulating α-SMA protein expression. Furthermore, we have also analyzed the biomechanical properties of the injured tibialis anterior. BMSC-Exos administration significantly improved the relative biomechanical indexes (Fast-twitch and tetanus strength) compared with the contusion group. This result demonstrated that the damaged muscle regained its biomechanical properties after BMSC-Exos treatment, rather than repair only at the cellular/histological level.
Our study found that BMSC-Exos could suppress the inflammatory reaction regulation of macrophage polarization. Specifically, BMSC-Exos significantly reduced the expression of proinflammatory factors (iNOS, TNF-α, IL-1β, and IL-6) and increased the expression of anti-inflammatory factors (TGF-β and IL-10) in the cell model. In addition, in the contusion model, proinflammatory factors (iNOS) were also downregulated by BMSC-Exos treatment. At 3 days, the relative muscle weight of TA muscle in BMSC-Exos group was significantly lighter than that in contusion group. It was reported that contusion injury induced a large number of neutrophils that infiltrated to the injured areas, which caused local inflammatory hematoma and wet weight increase in the early stage [41,42]. BMSC-Exos limited neutrophil infiltration and might facilitate earlier macrophage infiltration into the injured area. Studies have proved that BMSC-Exos also exerted anti-inflammatory effects in other tissues [43 –45].
Some studies have demonstrated that macrophage polarization plays an important role in inflammation regulation [46,47]. M1 macrophages infiltrated into the injured area and secreted many proinflammatory factors, including TNF-α, IL-1β, and IL-6, which resulted in enhancing local inflammatory reactions and cell proliferation [48]. However, if M1 macrophages are not converted into the M2 phenotype (anti-inflammatory class) in time, overexpression of proinflammatory factors would promote the production of excessive extracellular matrix by stimulating fibroblasts, which would ultimately result in scar tissue formation [49,50]. In this study, BMSC-Exos displayed the ability to induce the M2 polarization both in vitro and in vivo. In addition, we found that the effects of BMSC-Exo on muscle fibrosis and functional recovery were inhibited after macrophages depletion, which also indicated that the effects of BMSC-Exos in muscle healing were achieved by regulating macrophages.
Conclusion
Overall, this study demonstrated that the local administration of BMSC-Exos reduced fibrosis tissue, enhanced muscle regeneration, and improved biomechanical properties of the skeletal muscles after contusion. BMSC-Exos inhibited the inflammatory microenvironment in the injured muscle because of its promotion on M2 macrophages polarization. Our research provides strong support that BMSC-Exos can be useful for muscle healing in clinical practice.
Footnotes
Author Disclosure Statement
The authors declare that they have no competing interests, and all authors should confirm its accuracy.
Acknowledgments
The authors thank all of the members of our laboratory for their encouragement and assistance with this study. The authors also thank Qingyan Chen for her professional language editing.
Availability of Data and Materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Funding Information
This study was supported by the project of the National Natural Science Foundation of China (grant no. 81772419; 81972062).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.