
Abstract
A common observation in metabolic disorders and aging is the elevation of free fatty acids (FFAs), which can form ectopic fat deposition and result in lipotoxicity. Ectopic fat deposition of skeletal muscle has been recognized as an important component of aging, frailty, and sarcopenia. Previous studies have suggested that lipotoxicity caused by FFAs mainly stemmed from saturated fatty acids and decreased unsaturated/saturated fatty acid ratio in serum are also observed among metabolic disorder patients. However, the different effects of saturated fatty acids and unsaturated fatty acids on skeletal muscle are not fully elucidated. In this study, we verified that palmitate (PA), a saturated fatty acid, could lead to impaired differentiative capacity of C2C12 myoblasts by affecting Pax7, MyoD, and myogenin (MyoG), which are master regulators of lineage specification and the myogenic program. Then, oleate (OA), a monounsaturated fatty acid, were added to culture medium together with PA. Results showed that OA could ameliorate the impairment of differentiative capacity in C2C12 myoblast cells. In addition, we found PI3K/Akt signaling pathway played an important role during the process by RNA sequencing and bioinformatics analysis. The positive effect of OA on myoblast differentiative capacity disappeared if PI3K inhibitor LY294002 was added. In conclusion, our study showed that PA could destroy differentiative capacity of C2C12 myoblasts by affecting the expression of Pax7, MyoD, and MyoG, and OA could improve this impairment through PI3K/Akt signaling pathway.
Introduction
Skeletal muscle is one of the largest organs, accounting for about 40% of human body weight. It plays an important role, including sports, maintaining posture, endocrine and metabolism, and so on. Studies have shown that skeletal muscle decreases about 6%–7% per decade from middle-age period [1].
Sarcopenia is a kind of disease associated with increasing age, characterized by reduced skeletal muscle mass, along with the decline of muscle strength and/or physical function [2]. It can lead to weak activity ability, tumble, and increased mortality [3,4] and is jointly influenced by environmental factors and genetic factors, including hypokinesia, neuromuscular junction damage, the change of hormone related with age (such as insulin, estrogen and testosterone, growth hormone, glucocorticoid, etc.) and cytokines, inflammatory response, myocyte apoptosis, nutritional factors, and genetic changes, which can result in skeletal muscle mass, strength, and/or function decline by different mechanisms [5 –8].
At the cellular level, sarcopenia is displayed by the change of the proportion of muscle fiber type, muscle fiber atrophy, and lipid accumulation. In addition, increasing fibrous tissue and decreasing muscle satellite cells will occur at the later stage, which will lead to damaged skeletal muscle regeneration, repair, and remodeling [9,10]. Skeletal muscle regeneration is a complex process. Satellite cells remain quiescent normally. Once skeletal muscle is damaged, satellite cells are activated and begin to proliferate. Then they turn into myoblasts differentiating into mature muscle fibers, eventually merging with damaged muscle fibers or form new muscle fibers. The process requires a series of factors such as Myf5, Pax7, MyoD, and Myogenin (MyoG) [11].
Lipotoxicity, which can cause skeletal muscle damage and impaired regeneration [12], is an important risk factor for sarcopenia. A common observation in metabolic disorders and aging is the elevation of free fatty acids (FFAs), which can form ectopic fat deposition and result in lipotoxicity to target organs, such as the liver, skeletal muscle, and heart [13 –16]. Mitochondrial dysfunction, inflammation, oxidative stress, lipid accumulation, and insulin resistance are observed [17].
However, saturated fatty acids and unsaturated fatty acids make different contributions to lipotoxicity [16]. Previous studies in cardiac myocytes and hematopoietic precursor cell lines suggested that lipotoxicity caused by FFAs mainly stemmed from saturated fatty acids [18,19]. Reports have also shown that decreased unsaturated/saturated fatty acid ratio in serum are observed among metabolic disorder patients [20]. These results indicated that increased free saturated fatty acids or decreased unsaturated/saturated fatty acid ratio might promote nonadipose tissues with ectopic fat deposition damage, such as skeletal muscle.
However, the different effects of saturated fatty acids and unsaturated fatty acids on skeletal muscle are not fully elucidated. C2C12 myoblast model is always used to recapitulate some basic mechanisms of skeletal muscle regeneration. Therefore, this study was aimed to confirm the effect of palmitate (PA), a saturated fatty acid, and oleate (OA), a monounsaturated fatty acid, on differentiative capacity of C2C12 myoblasts and explore the mechanism to provide new ideas and methods for the prevention and treatment of sarcopenia.
Materials and Methods
Preparation of stock solutions of fatty acids
FFAs were conjugated with fatty acid-free bovine serum albumin (BSA, 36104ES60; Yeasen Biotech) using a modified method described by Cousin [21]. In brief, we prepared FFA stock solution first using PA (P0500; Merck) or OA (O1383; Merck). Around 100 mM PA or OA stock solution was prepared in 0.1 M NaOH by heating at 70°C in a shaking water bath. Then these solutions were mixed with 10% fatty acid-free BSA in deionized water at 55°C.
In our study, the concentration of FFA/BSA stock solutions was 5 mM. They were prepared by adding 50 μL 100 mM FFA stock solution to 950 μL 10% fatty acid-free BSA in deionized water, then vortex mixed for 10 s followed by a further 10-min incubation at 55°C. The FFA/BSA complexed solutions were cooled to room temperature and sterile filtered using 0.45-mm pore size membrane filters (SLHV033RB; Merck). The FFA/BSA stock solutions were stored at −20°C to prevent oxidation. Stock solutions were heated for 15 min at 55°C, then cooled to room temperature before using them.
Cell culture
Mouse C2C12 myoblasts (Fuheng Cell Center, Shanghai, China) were grown in the growth medium (GM) with Dulbecco's Modified Eagle Medium (DMEM) (Gibco, Waltham, MA), 10% fetal bovine serum (Gibco), and 1 U penicillin/streptomycin (15140122; Thermo Fisher Scientific, Waltham, MA) at 37°C in an atmosphere of 95% humidity and 5% CO2. When cells reached 90% confluence, the GM was changed to the differentiation medium (DM) with DMEM, 2% heat-inactivated horse serum (HS; Gibco), and 1 U penicillin/streptomycin. After DM was added, cells were incubated for 12, 24, 48, 72, and 96 h to analyze the dynamic change of myogenic factors between BSA control group and 100 μM PA group.
In addition, cells were harvested at 48 or 96 h after differentiation to observe the change of early or late myogenic factors in BSA control or 100 μM PA or 100 μM OA or equimolar (100 μM) PA and OA (PO) groups. The concentration of BSA was equal in every group. When needed, myoblasts were exposed to PO in combination with PI3K inhibitor LY294002.
RNA isolation and real-time quantitative PCR analysis
Total RNA from C2C12 myoblasts were isolated using the TRIzol regent (Invitrogen, Waltham, MA). One thousand nanograms of total RNA was reverse transcribed into cDNA with the Prime Script RT Reagent Kit (TaKaRa) after RNA concentration was measured. cDNA was mixed with the SYBR Green PCR Master Mix Reagent Kit (TaKaRa) before mRNA was measured using quantitative real-time polymerase chain reaction (PCR) with the StepOnePlus Real-time PCR Instrument (Applied Biosystems, Waltham, MA). All samples were prepared in technical duplicates. The data were analyzed for fold change using 2−ΔΔCt. The 18s ribosomal RNA (18s) served as an internal control. Primer sequences were listed in Supplementary Table S1.
Western blotting
Protein extracts were obtained after cultured C2C12 myoblasts were collected using radioimmunoprecipitation assay (RIPA) lysis buffer (P0013B; Beyotime) supplemented with 1 mM phenylmethanesulfonyl fluoride (PMSF, ST506; Beyotime), and 1 mM phosphatase inhibitors (ab201112; Abcam).
Then, proteins were denatured at 99°C for 10 min with 5 × sodium dodecyl sulfate (SDS) loading buffer (LT101S, Epizyme). SDS-polyacrylamide gel electrophoresis was performed using an 8% or 10% gel to separate protein. Thereafter, proteins in the gels were transferred onto polyvinylidene fluoride (PVDF, IPVH00010; Merck) membranes, and the membranes were blocked with 5% BSA (A500023-0100, Sangon Biotech) in 1 × TBST with tris-buffered saline (TBS; B548105-0500, Sangon Biotech) and tween (ST825; Beyotime) for 2 h at room temperature. The membranes were then incubated overnight at 4°C with the primary antibodies against β-tubulin (abs830032, 1:1,000; Absin), MyoD (sc-377460, 1:100; Santa Cruz Biotechnology), MyoG (ab1835, 1:250; Abcam), p-AKT (4060, CST, 1:1,000), t-Akt (4691, CST, 1:1,000), and MyHC (MF20, 1:1,000; Developmental Studies Hybridoma Bank). The PVDF membranes were washed in TBST and incubated with anti-mouse IgG, HRP-linked secondary antibody (7076s, CST, 1:2,000) or anti-rabbit IgG, HRP-linked secondary antibody (7074P2, CST, 1:2,000) for 1.5 h at room temperature.
Reacting bands were detected with electrochemiluminescence detection reagents (34076; Thermo Fisher Scientific) after they were washed in TBST. β-Tubulin served as an internal control.
Immunofluorescence analysis
Cells were fixed in 4% paraformaldehyde (PFA, P0099; Beyotime) on slides for 30 min. Slides were washed in phosphate-buffered saline (PBS) (ST476; Beyotime) and were blocked with 5% goat serum and 0.3% Triton (ST795; Beyotime) in PBS for 2 h at room temperature. The cells were then incubated overnight at 4°C with the primary antibodies against MyoD (sc-377460, 1:50; Santa Cruz Biotechnology) and MyHC (MF20, 1:100; Developmental Studies Hybridoma Bank). Slides were washed in PBS again and incubated with the secondary antibodies against mouse IgG (H+L), F(ab’)2 Fragment (Alexa Fluor® 488 Conjugate) (4408S, CST, 1:500) for 1.5 h at room temperature. Then 4′,6-diamidino-2-phenylindole (DAPI) was used to visualize nuclei after slides were washed in PBS. In the end, Fluorescence Mounting Medium (S3023; Dako) was used to prevent fluorescence from disappearing.
Stained cells were viewed under a fluorescence microscope (Leica, Germany) and analyzed by ImageJ. The differentiation index was calculated as the percentage of nuclei in MyHC-positive cells. The fusion index was calculated as the percentage of MyHC-positive myotubes that contained at least two nuclei.
Oil Red O staining
Cells were fixed in 4% PFA for 30 min after being washed with PBS. Then cells were washed with PBS three times and stained with Oil Red O solution for 2–3 h at room temperature. Lipid accumulation was observed with a light microscope and photographed after residual Oil Red O was eluted with 60% isopropanol.
RNA sequencing
The integrity and quantitation of RNA were measured using the RNA Nano 6000 Assay Kit of the Bioanalyzer2100 system (Agilent Technologies, Santa Clara, CA) and the Nano Photometer® spectrophotometer (IMPLEN, Westlake Village, CA), respectively. Sequencing libraries were generated using the NEBNext® Ultra TMRNA Library Prep Kit for Illumina® (NEB, Ipswich, MA) following the manufacturer's recommendations and index codes were added to attribute sequences to each sample. Library quality was assessed on the Agilent Bioanalyzer2100 system. The clustering of the index-coded samples was performed on a cBot Cluster Generation System using the TruSeq PE Cluster Kit v3-cBot-HS (Illumina) according to the manufacturer's instructions. After cluster generation, the library preparations were sequenced on an Illumina Hiseq platform and 125 bp/150 bp paired-end reads were generated. Three repeats from each group were performed.
Identification of differentially expressed genes
Data were analyzed after experiencing RNA quantification and qualification, library preparation, clustering, and sequencing. Differentially expressed gene (DEG) analyses between two groups were performed using the DESeq2 R package (1.16.1). Genes with P < 0.05 and |logFC| > 1 found by DESeq2 were assigned as differentially expressed. Volcano plots and Heatmap were visualized through Pheatmap R package.
Enrichment analyses of gene function and pathways
Gene ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway analysis of DEGs were implemented by online website DAVID (
Statistical analyses
In this study, each experiment was performed five times. For single comparison between two groups, Student's t-test was used. The quantitative data were expressed as the mean ± standard deviation. All statistical analyses were performed using the GraphPad Prism 6.0. Threshold of significance was defined as P < 0.05.
Results
PA impairs differentiative capacity of C2C12 myoblasts
As shown in Fig. 1, C2C12 myoblasts expressed different myogenic regulatory factors (MRFs) in different periods of myoblast differentiation (Fig. 1A). To explore the role of PA in differentiative capacity of C2C12 myoblasts, we treated myoblasts with 100 μM PA and tested the levels of MyoD and MyoG, which were important MRFs during myoblast differentiation, after differentiation for 12, 24, 48, 72, and 96 h. Reverse transcription-polymerase chain reaction (RT-PCR) showed the levels of MyoD, which is one of the earliest myogenic markers, were increased gradually within 48 h after differentiation, and decreased after 48 h in BSA group. The levels of MyoD were increased continually within 96 h after differentiation in PA group. However, the degrees of elevation were lower compared with BSA group. The differences were significant at 12, 24, and 48 h after differentiation, whereas the levels of MyoD were similar at 72 and 96 h after differentiation (Fig. 1B).
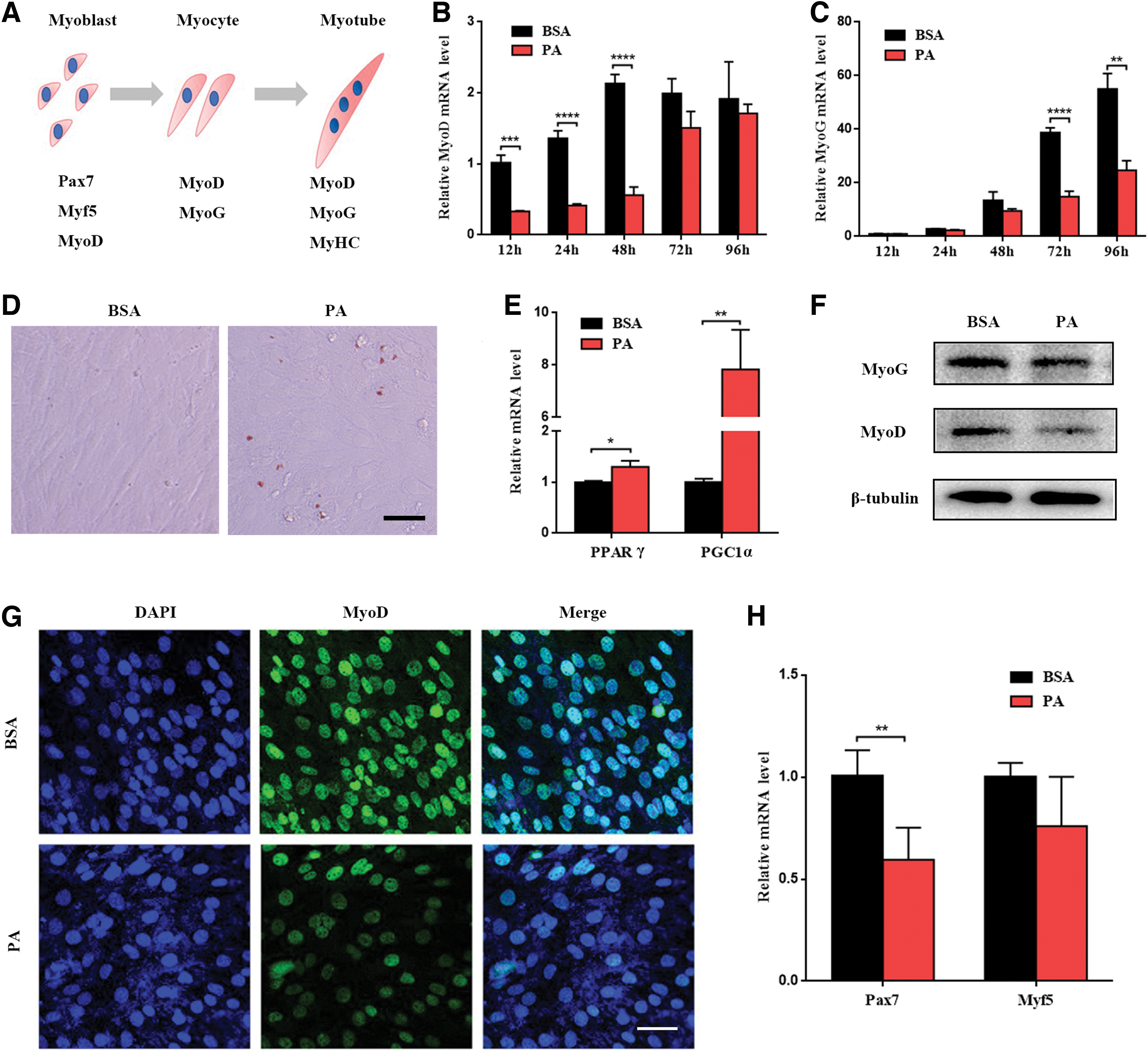
PA impaired differentiative capacity of C2C12 myoblasts.
The levels of MyoG, which is a crucial gene that regulates the secondary muscle fibers' maturation and occur after MyoD, were all increased gradually within 96 h after differentiation in two groups. However, the degrees of elevation in PA group were lower at all time points than BSA group. The differences were obvious at 72 and 96 h after differentiation and were not significant at 12, 24, and 48 h (Fig. 1C). The results might be attributed to the slow elevation of MyoG at the first 48 h and rapid elevation of MyoG after differentiation from 48 h. Given that the levels of MyoD were highest at 48 h after differentiation and the levels of MyoG were increased obviously at 72 and 96 h, we chose 48 h as the time point to explore the influence of PA on the differentiative capacity of C2C12 myoblasts in the early stage and 96 h after differentiation to analyze the influence on late differentiation.
First, we examined the lipid accumulation with Oil Red O staining at 48 h after differentiation. The results showed lipid accumulation occurred in PA group (Fig. 1D). Peroxisome proliferator-activated receptor gamma (PPARγ) and PPARγ coactivator-1α (PGC-1α) are also higher in PA group (Fig. 1E). Western blots showed that the levels of MyoD were reduced in PA group and the levels of MyoG were similar between two groups (Fig. 1F). The results were in accord with RT-PCR. Immunofluorescence staining indicated the same result (Fig. 1G).
Besides, we further investigated the change of some markers, which could be expressed in the early phase of differentiation and regulate myoblast differentiation, including Myf5 and Pax7. RT-PCR indicated that Myf5 did not show any obvious change between the two groups. However, Pax7 reduced in PA group compared with BSA group (Fig. 1H). The results demonstrated that PA potently impaired differentiative capacity of C2C12 myoblasts in the early stage of differentiation.
OA ameliorates PA-induced impairment of differentiative capacity in C2C12 myoblasts
DM was supplemented with BSA control, 100 μM PA, 100 μM OA, or equimolar (100 μM) PA and OA (PO) for 48 h. Compared with BSA control group, the mRNA levels of MyoD increased in OA group (Fig. 2A), but the protein levels of MyoD did not change significantly (Fig. 2B, C). Immunofluorescence staining showed the same result (Fig. 2D). Whereas, the mRNA and protein levels of MyoD decreased markedly in PA group compared with BSA group (Fig. 2A–C). Immunofluorescence staining intensity also weakened (Fig. 2D). Then, we found that the mRNA and protein levels of MyoD significantly increased when OA was added to PA group (Fig. 2A–C). In addition, immunofluorescence assays showed increasing MyoD expression in PO group compared with PA group (Fig. 2D). Although the mRNA levels of Pax7 did not significantly change between BSA group and OA group, variations were the same as MyoD between other groups (Fig. 2A). The results indicated that OA ameliorated PA-induced impairment of differentiative capacity in C2C12 myoblasts.
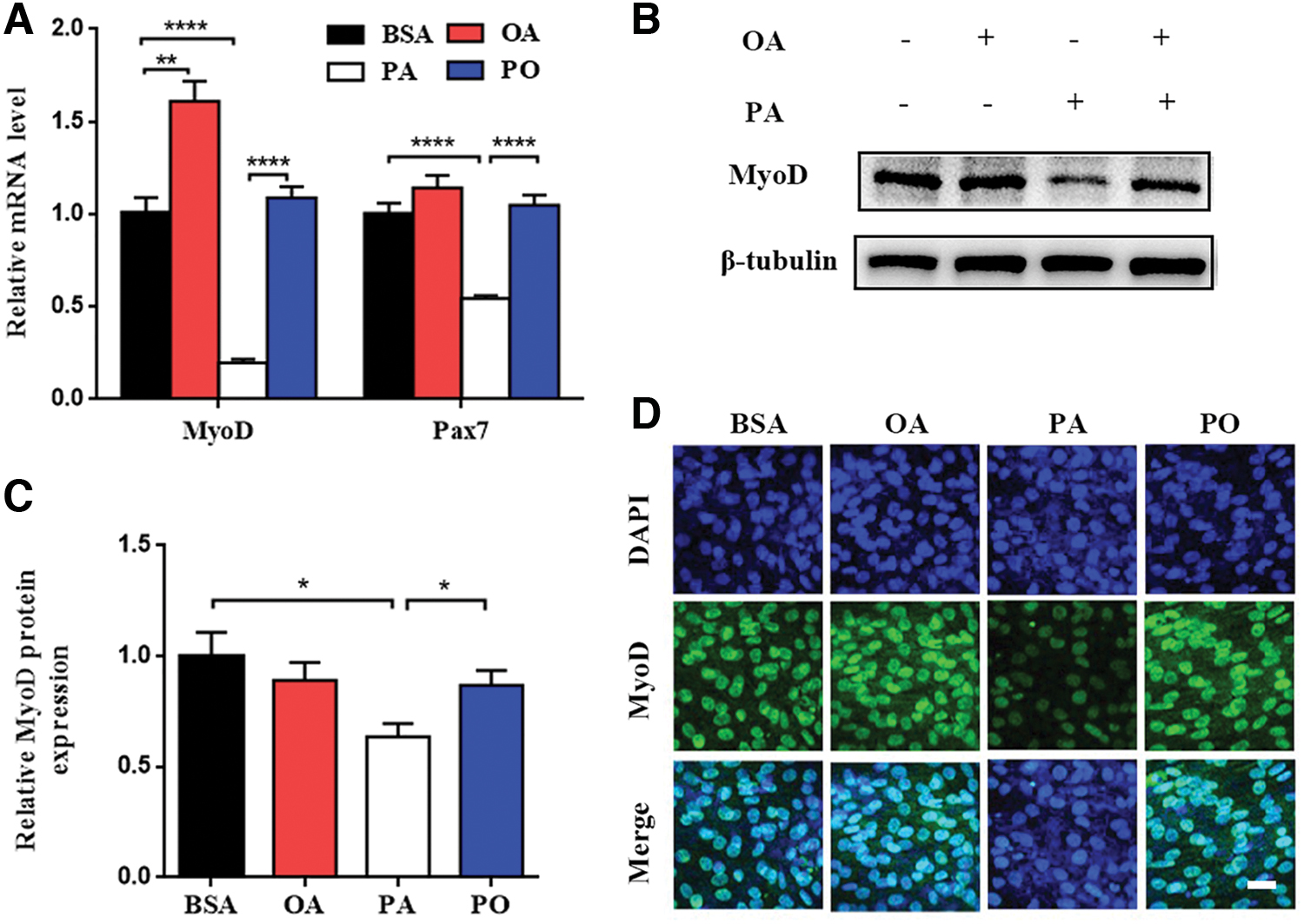
OA ameliorated PA-induced impairment of differentiative capacity in C2C12 myoblasts. C2C12 myoblasts were treated with BSA control, OA (100 μM), PA (100 μM), and equimolar (100 μM) PA and OA (PO) after differentiation for 48 h.
Microarray analysis finds the DEGs and results of GO and KEGG enrichment analyses between PA and PO groups
To identify key genes and functions during the progression of myoblast differentiation between PA and PO groups, we analyzed the DEGs between two groups. There were 2947 DEGs (1196 downregulated and 1751 upregulated genes), which were screened (P < 0.05 and |logFC| > 1) from all DEGs. Heatmap and volcano plots were visualized to show the correlation between DEGs (Fig. 3A, B).
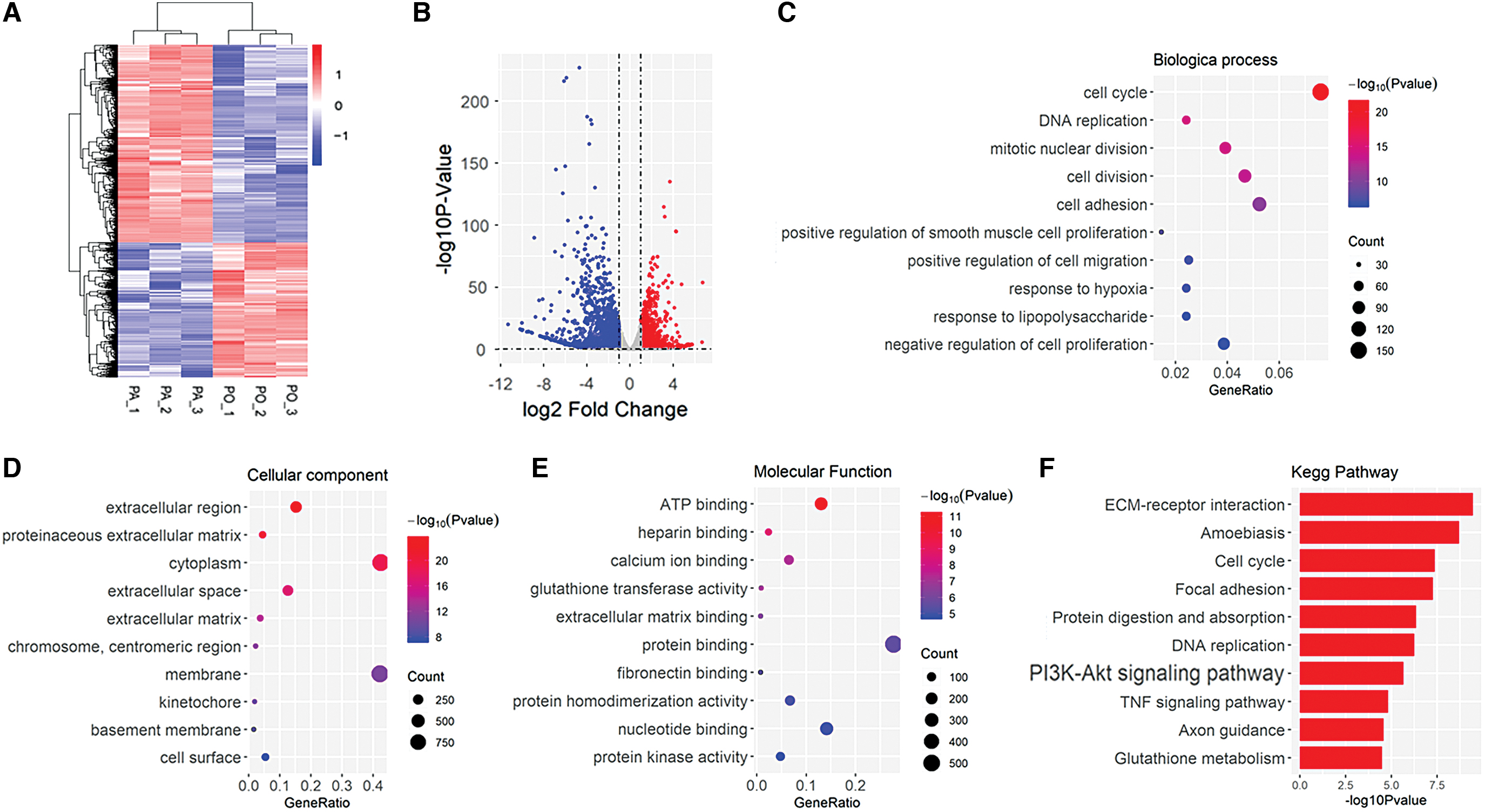
Identification of DEGs and function enrichment between PA and PO groups.
Furthermore, we performed GO enrichment analysis to explore potential roles of DEGs between PA and PO groups. All screened DEGs were uploaded to the online website DAVID to discern GO classification. The results of GO enrichment analysis were in Fig. 3C–E. The DEGs were mainly enriched in cell cycle, DNA replication, mitotic nuclear division, cell division, cell adhesion, positive regulation of smooth muscle cell proliferation, positive regulation of cell migration, the response to hypoxia, response to lipopolysaccharide, and negative regulation of cell proliferation in biological process (BP) (Fig. 3C), extracellular region, proteinaceous extracellular matrix (ECM), cytoplasm, extracellular space, ECM, chromosome, membrane, kinetochore, basement membrane, and cell surface in cellular component (CC) (Fig. 3D), and ATP binding, heparin binding, calcium ion binding, glutathione transferase activity, ECM binding, protein binding, fibronectin binding, protein homodimerization activity, nucleotide binding, and protein kinase activity in molecular function (MF) (Fig. 3E).
KEGG pathway analysis showed that DEGs were mainly enriched in ECM/receptor interaction, amoebiasis, cell cycle, focal adhesion, protein digestion and absorption, DNA replication, PI3K/Akt signaling pathway, TNF signaling pathway, axon guidance, and glutathione metabolism (Fig. 3F). The most number of genes were enriched in PI3K/Akt signaling pathway in the top 10 KEGG pathways.
The PI3K/Akt signaling pathway is known to positively regulate myogenic differentiation. We also found the PI3K/Akt signaling pathway was significantly different between the two groups. In addition, many studies have demonstrated that OA could activate the PI3K/Akt pathway to regulate peripheral metabolism. To test this hypothesis, the change of PI3K/Akt pathway was examined. We compared the levels of total Akt (t-Akt) and phosphorylated Akt (p-Akt) between BSA, OA, PA, and PO groups. The results indicated the p-Akt/t-Akt level significantly elevated in OA group and reduced in PA group compared with BSA group. However, the p-Akt/t-Akt level was improved in PO group compared with PA group (Fig. 4A, B).
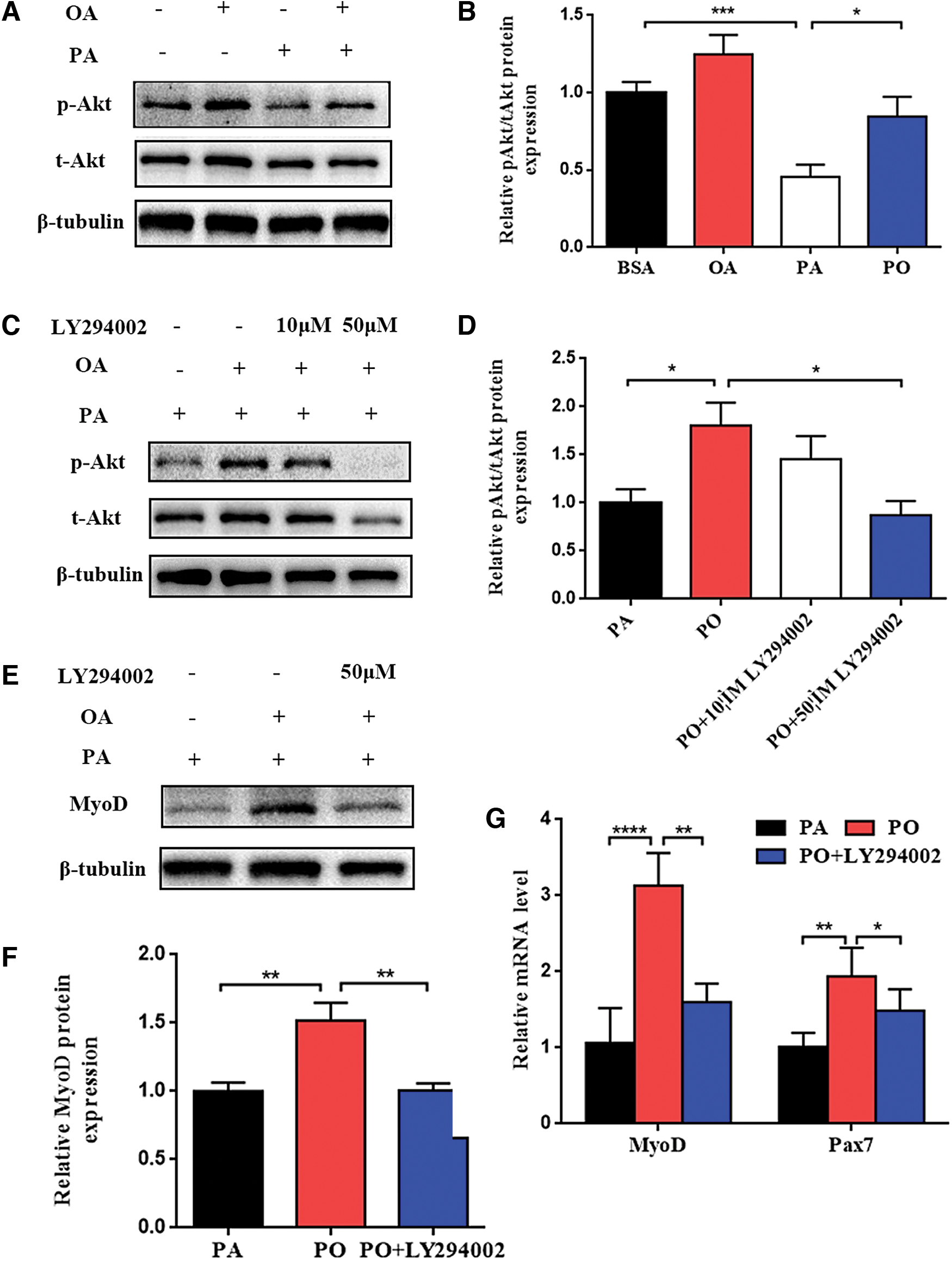
OA could rescue PA-induced impairment of differentiative capacity in C2C12 myoblasts through PI3K/Akt signaling pathway.
OA can rescue PA-induced impairment of differentiative capacity in C2C12 myoblasts through PI3K/Akt signaling pathway
To further determine whether OA rescues PA-induced impairment of myoblast differentiative capacity through PI3K/Akt pathway in vitro, C2C12 myoblasts were treated with PO in the presence or absence of the PI3K inhibitor LY294002. Different concentrations (10, 50 μM) of LY294002 were added to DM with PO at the beginning of differentiation for 48 h. The p-Akt/t-Akt levels were similar in the presence or absence of 10 μM LY294002, and markedly reduced in the presence of 50 μM LY294002 (Fig. 4C, D). Therefore, we chose the concentration of 50 μM LY294002 in further experiments.
The levels of MyoD and pax7 were examined between PA and PO with the presence or absence of 50 μM LY294002. The results showed that PO could increase the protein and mRNA levels of MyoD, which were consistent with our previous results. However, the improvement was reduced if the PI3K inhibitor LY294002 was added to PO group (Fig. 4E–G). The change of the mRNA levels of Pax7 were the same as MyoD (Fig. 4G). Taken together, these data demonstrated that OA could rescue PA-induced impairment of differentiative capacity through PI3K/Akt signaling pathway in C2C12 myoblasts.
OA can improve the inhibition of PA on the formation of myotubes
Previous results showed that PA impaired early differentiative capacity of C2C12 myoblasts. To further explore the effect of PA on the late differentiation and formation of myotubes and whether OA can improve the impairment, the myoblast differentiation time was extended to 96 h to induce the formation of myotubes. The results were compared between BSA and PA and PO with or without 50 μM LY294002 groups. Immunofluorescence staining showed that PA inhibited MyHC expression and the formation of myotubes (Fig. 5A). MyHC expression and the formation of myotubes increased after OA was added to culture medium together with PA (Fig. 5A). However, LY294002 impeded the recovery of MyHC expression and the formation of myotubes even though OA was added to PA group. The differentiation index and fusion index were also employed to evaluate roles of PA, OA, and LY294002 (Fig. 5B, C).
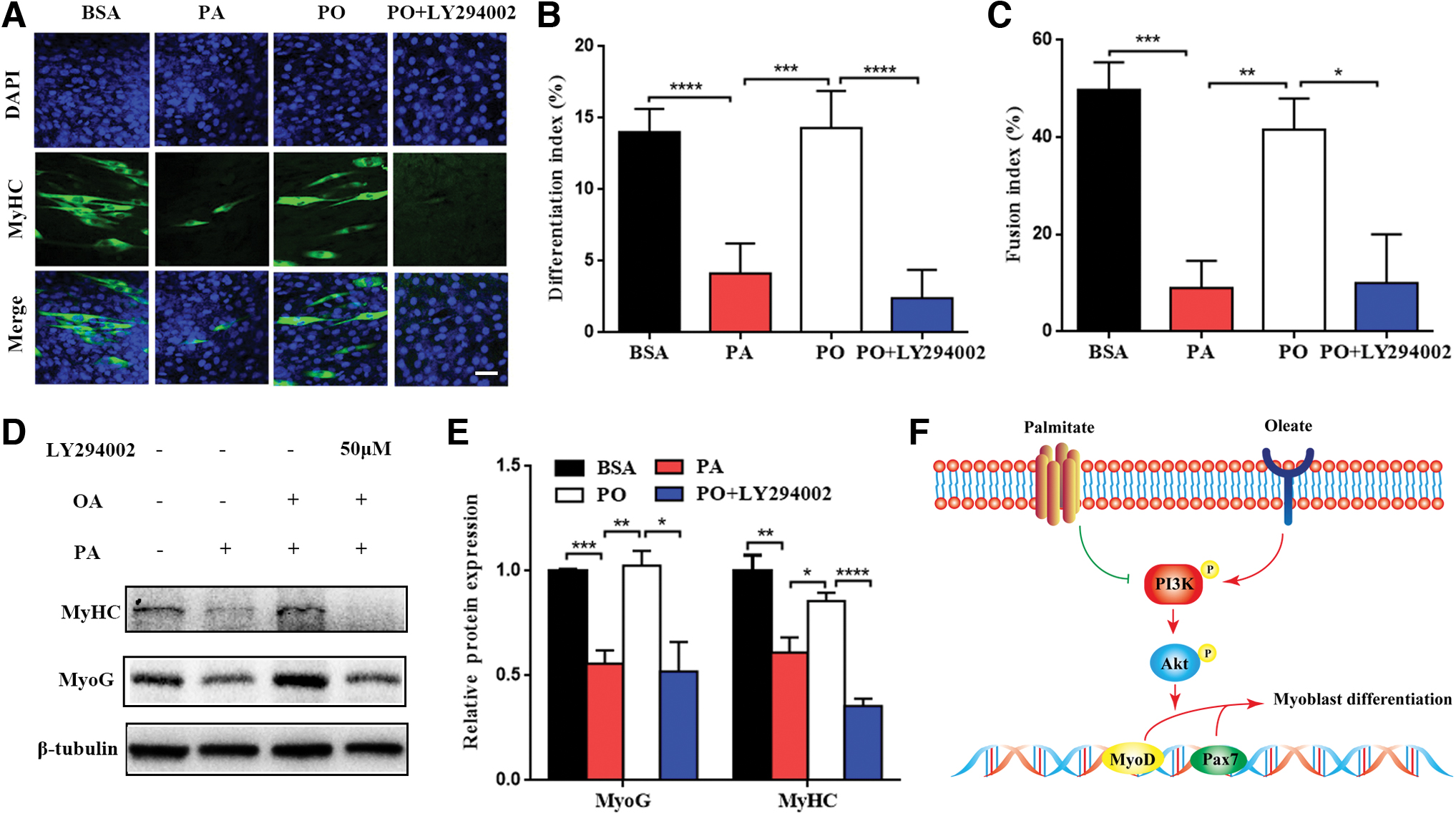
OA could improve the inhibition of PA on the formation of myotubes. C2C12 myoblasts were treated with BSA control, PA (100 μM), equimolar (100 μM) PA and OA (PO) with or without PI3K inhibitor LY294002 after differentiation for 96 h.
In addition, we examined the protein levels of MyoG and MyHC. The results were the same (Fig. 5D, E). In summary, PA-induced impairment of differentiative capacity and myotube formation can be rescued by OA. Once the PI3K/Akt signaling pathway was inhibited by LY294002, the positive influence weakened. That is to say, OA can ameliorate PA-induced impairment of differentiative capacity and myotube formation through the activation of PI3K/Akt signaling pathway in C2C12 myoblasts (Fig. 5F).
Discussion
Our study verified that PA could induce impairment of differentiative capacity in C2C12 myoblasts. Meanwhile, we found that OA could improve the impaired differentiative capacity and clarified the probable mechanism by RNA sequencing and bioinformatics analysis for the first time. Myoblast differentiation is a complex process which is regulated by a series of genes, including all kinds of transcription factors. The researches about MRFs (MyoG, MyoD, Myf5, Mrf4) have been widely developed and become relatively mature. Myf5 and MyoD are the early markers of myoblast differentiation. MyoG and Mrf4 regulate secondary muscle fibers to mature [22]. MyoD, which acts as a myogenic determination gene [23], and MyoG, which is essential for the terminal differentiation of myoblasts [24], have been studied most widely. They can regulate the transcription of most of the muscle-specific genes. These factors and other transcription factors such as pax7 and SIX commonly adjust myoblast differentiation [25 –29].
Previous studies have shown that saturated fatty acid can induce reactive oxygen species (ROS), inflammation, insulin resistance (IR), and mitochondrial dysfunction [30,31]. These conditions may lead to cell function change and even death. PA is one of the most representative saturated fatty acids. There has been evidence that PA can inhibit the myoblast differentiation. Our research reached the same conclusion. MyoD, the early marker of myoblast differentiation, reduced in PA group. Lots of studies have suggested that MyoD is the key to myoblast differentiation.
Except muscle cells, myoblasts can also differentiate into other cells, including adipocytes and osteoblasts. PPARγ and PGC1α play important roles in the fate of satellite cell differentiation [32,33]. PPARγ is a nuclear transcription factor that regulates adipocyte differentiation and lipogenic genes during adipogenesis. PGC1α is related to mitochondrial biogenesis and thermogenesis [34]. The elevation of PPARγ and lipid accumulation indicated that the process of myoblast differentiation into myotubes was hindered. The disappearance of the MyoD can lead to the failure of myoblast differentiation into muscle cells [35].
Pax7 is an important gene expressed in myoblasts and has a great influence on myoblast differentiation. On the one hand, Pax7 is a master regulator of early lineage specification to maintain the potency of myoblast differentiation into muscle cells [25]. Studies have shown that the lack of Pax7 can lead to the failure of muscle satellite cells differentiating into muscle cells [29,36,37]. On the other hand, Pax7 is highly expressed in proliferating myoblasts. Once myoblasts begin differentiation, the expression of Pax7 is downregulated gradually [38]. Our study shows that the expression of Pax7 was reduced in PA group, along with reduced MyoD. Therefore, we speculated that the results might be associated with the impaired potency of myoblast differentiation.
MyoD functions downstream from Pax7, but Myf5 can act in parallel with the Pax transcription factors [28,39,40]. This may explain why Myf5 has no obvious change, although MyoD reduced in PA group. Besides, MyoG also did not change between the two groups, which might be associated with the length of differentiation time.
Differing from saturated fatty acids, monounsaturated fatty acid is beneficial to human physiology. It can regulate blood lipid and blood sugar to reduce the risk of cardiovascular events. High monounsaturated fat diet has caught more and more attention recently. Several RCT meta analyses showed that high monounsaturated fat diet can improve some indicators of the metabolism such as blood sugar, blood fat, and blood pressure compared with high-carbohydrate diets [41 –44]. Besides, monounsaturated fatty acids can reverse the lipotoxicity of saturated fatty acid, such as oxidative stress, inflammation, and insulin resistance [31,45]. Taking these effects into consideration, we wanted to observe whether the monounsaturated fatty acid can improve the influence of the saturated fatty acid on myoblast differentiation.
OA is the most representative monounsaturated fatty acid. Our results showed that the protein levels of MyoD were not significantly different and the mRNA levels were higher when only OA was added to culture medium compared with BSA group. However, the distinction in mRNA levels was not apparent. The mRNA levels of Pax7 were also similar between two groups. Nevertheless, the levels of MyoD and Pax7 increased obviously when OA was added to PA group. The results implied that the inhibitory effect of PA on myoblast differentiation can be improved by OA.
Microarray analysis showed that the most significant BP enriched were cell cycle, DNA replication, mitotic nuclear division, cell division, cell adhesion, positive regulation of smooth muscle cell proliferation, positive regulation of cell migration, the response to hypoxia, response to lipopolysaccharide, and negative regulation of cell proliferation. Cell cycle withdrawal plays an important role in myoblast differentiation and muscle regeneration. First of all, satellite cells will be activated largely. Then, myoblasts exit to cell cycle and the expressions of some MRFs, such as MyoD and MyoG increase. These changes promote myoblast differentiation and the formation of myotubes [25 –29,46]. In the processes, a lot of genes related with cell cycle, cell division, and cell proliferation change.
Cell components showed that the most significant CC were enriched in extracellular region, proteinaceous ECM, cytoplasm, extracellular space, ECM, chromosome, membrane, kinetochore, basement membrane, and cell surface. The results display that changes were mainly in extracellular region and the cell surface. Indeed, we know that myoblast differentiation is regulated by various factors inside and outside cells. Many extracellular factors play important roles during differentiation, such as growth factors and cytokines. Several growth factors and cytokines, leukemia inhibitory factor (LIF), transforming growth factor (TGF-β), and hepatocyte growth factor (HGF), have had been implicated as key factors in the processes [47 –52]. These secreted factors work through combining with cell surface receptors to deliver signals into the inside of cells. Our results are consistent with the theory.
MF showed that ATP binding, heparin binding, calcium ion binding, glutathione transferase activity, ECM binding, protein binding, fibronectin binding, protein homodimerization activity, nucleotide binding, and protein kinase activity are most important. The primary MF is the ATP binding. As is known to all, mitochondria are the site of ATP production. Lots of studies have shown that mitochondrial function is closely related to myoblast differentiation [34,53]. Mitochondrial dysfunction may lead to the damage of ATP generation and binding. Calcium ion balance is also critical for myoblast differentiation [54]. Keeping with the CC, ECM also has changed obviously.
In addition to GO enrichment analysis, we also analyzed the KEGG pathway. Results showed the most important pathways were ECM/receptor interaction, amoebiasis, cell cycle, focal adhesion, protein digestion and absorption, DNA replication, PI3K/Akt signaling pathway, TNF signaling pathway, axon guidance, and glutathione metabolism. The majority of pathways agreed with GO enrichment analysis, including ECM receptor interactions and cell cycle. Beyond that, most number of genes were enriched in PI3K/Akt signaling pathway.
Previous reports have shown that the activation of PI3K/Akt pathway can promote myoblast differentiation [55]. Akt activation can enhance the activity of transcription factors MyoD to induce myoblast differentiation and fusion into mature muscle fibers [56 –58]. PI3K/Akt pathway is a connection from the cell surface to the intracellular signal transmission of transcription factors, which affect muscle regeneration and myotube fusion. Adding exogenous molecules involved PI3K/Akt signaling pathway, such as apelin-13 peptide, Sphingosine 1-phosphate lipid (S1P) 16 or 17, or growth factors, such as IGF or vascular endothelial growth factor can improve muscle regeneration repair after injury [59 –62].
Some reports indicated that OA could improve insulin resistance by PI3K/Akt pathway and rescue the toxicity of PA [45,63,64]. However, the effect was not validated during the myoblast differentiation. Our results show that the level of p-Akt/t-Akt was reduced in PA group comparing with BSA group. The level of p-Akt/t-Akt recovered after OA was added. The positive effect disappeared after adding PI3K inhibitors. The change was consistent with the change of MyoD, Pax7, and MyoG levels. The results showed that PA could impair differentiative capacity and OA could rescue the impairment through stimulating PI3K/Akt pathway in C2C12 myoblasts. Nevertheless, PI3K/Akt pathway is affected by PA through which specific pathways or molecules remain to be further studied, which provides a new direction for future researches.
Conclusions
In conclusion, our study showed that the PA could damage myoblast differentiative capacity by affecting Pax7, MyoD, and MyoG, and OA could improve this impairment through the activation of PI3K/Akt signaling pathway. It provides a new diet intervention and treatment advice for the elderly or obese patients with high FFA to maintain skeletal muscle mass and delay the development of sarcopenia.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The research was supported by grants from the National Key Research and Development Program of China (No: 2018YFC2002000), the National Natural Science Foundation of China (No: 81901408 and 82071581), Shanghai Sailing program (No: 19YF1414500), Shanghai Medical Leadership Training Program (2019LJ09), and Shanghai Science and Technology Committee Project (17411950702).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.