
Abstract
Mesenchymal stem cells (MSCs) isolated from adipose tissue (adipose-derived stem cells [ADSCs]) are considered one of the most promising cell types for applications in regenerative medicine. However, the regenerative potency of ADSCs may vary because of heterogeneity. Long-term trypsin treatment (LTT) is known to significantly concentrate multilineage-differentiating stress-enduring (Muse) cells from human MSCs. In this study, we aimed to generate cells with high stem cell potency from canine ADSCs using LTT. After 16 h of treatment with trypsin, surviving ADSCs (LTT-tolerant cells) had significantly enhanced expression of stage-specific embryonic antigen (SSEA)-1, a mouse embryonic stem cell marker, and fucosyltransferase 9, one of several fucosyltransferases for SSEA-1 biosynthesis. However, LTT-tolerant cells did not enhance the expression of SSEA-3, a known human Muse cell marker. LTT-tolerant cells, however, showed significantly higher self-renewal capacity in the colony-forming unit fibroblast assay than ADSCs. In addition, the LTT-tolerant cells formed cell clusters similar to embryoid bodies and expressed undifferentiated markers. Moreover, these cells differentiated into cells of all three germ layers and showed significantly higher levels of α 2–6 sialic acid (Sia)-specific lectins, known as differentiation potential markers of human MSCs, than ADSCs. LTT-tolerant cells had a normal karyotype and had low telomerase activity, showing little carcinogenetic potency. LTT-tolerant cells also showed significantly increased activity of transmigration in the presence of chemoattractants and had increased expression of migration-related genes compared with ADSCs. In addition, LTT-tolerant cells had stronger suppressive activity against mitogen-stimulated lymphocyte proliferation than ADSCs. Overall, these results indicated that the LTT-tolerant cells in canine ADSCs have similar properties as human Muse cells (although one of the undifferentiated markers is different) and are expected to be a promising tool for regenerative therapy in dogs.
Introduction
The clinical application of stem cells in regenerative medicine is expected to result in tissue repair and functional recovery of lost tissues in many intractable diseases. Regenerative research using embryonic stem (ES) cells, induced pluripotent stem (iPS) cells, and mesenchymal stem cells (MSCs) have been actively conducted.
Although ES cells and iPS cells have the ability to differentiate into three germ layers, these cells have a tumorigenic risk in vivo [1 –3]. Hence, only differentiated cells must be directly transplanted to damaged tissues [4]. On the contrary, MSCs have some advantages, such as low risk of malignant transformation and homing ability to damaged tissues, despite their limited differentiation potential [5 –7]. In addition, MSCs have been reported to secrete a variety of growth factors and cytokines, inhibit T cell activation, and exhibit trophic effects of angiogenesis [6 –10]. Hence, MSCs are viable cells with high potential for clinical application.
MSCs are typically isolated as adherent cells from the mesenchymal tissues. Owing to the simple methods of isolation, MSCs are a heterogeneous population of cells containing cells other than stem cells such as endothelial cells, fibroblasts, and progenitor cells [4,5,11]. In addition, it has been indicated that there are some subpopulations of MSCs with a variety of stemness [12 –15]. Therefore, MSCs are expected to be purified to enhance the high therapeutic effects.
Multilineage-differentiating stress-enduring (Muse) cells that are double-positive for stage-specific embryonic antigen (SSEA)-3/CD105. They are rare pluripotent subpopulations within human MSCs and have the ability to differentiate into three germ layers [1]. Some of the advantages of Muse cells are that they have the high homing capacity to damaged tissues, can differentiate into specific or a variety of cells depending on the environment, and can contribute to tissue repair and functional recovery by intravenous (IV) administration [16 –18]. On the contrary, ES cells and iPS cells under undifferentiated state cannot be administered to patients. Therefore, Muse cells are the ideal cell type and superior to ES cells and iPS cells for clinical application [1,3].
A first step in the clinical application of Muse cells is to validate the scientific method and to collect the data using animal models. Dogs are unrivaled animal models for research of human diseases [19]. They share a common living environmental and lifestyle with humans. It has been reported that similar and spontaneous diseases such as intervertebral disc herniation, inflammatory bowel disease, Sjogren's disease, and osteoarthritis occur in dogs, unlike in laboratory rodents [20]. The results from this study and the development of Muse cell therapy in dogs might have the potential to serve as an animal model with future applications in human medicine.
Muse cells are collected by cell sorting from human MSCs [21] and by putting lipoaspirated human fat under severe cellular stress treatment [22]. However, these methods are not convenient and reasonable for clinical application because specific equipment and a large quantity of fat are necessary. On the contrary, long-term trypsin treatment (LTT) is an effective method of collecting Muse cells from human MSCs [1] and is easy to use. Adipose-derived stem cells (ADSCs) consist of a type of MSCs that are accessible and useful for clinical applications because fat tissues can be collected easily and compared with bone marrow-derived stem cells (BMSCs), ADSCs have high proliferative ability [23]. The aim of this study was to generate the cells with the high stem cell potency from canine ADSCs with LTT compared with ADSCs without LTT.
Materials and Methods
Ethics statement
The animal experiments were approved by the Institutional Animal Care and Use Committee of Osaka Prefecture University. The animals were neither operated on, nor sacrificed for this study. The uses of tissues were approved by the ethics committee of a local veterinary clinic. All dogs were privately owned and the owners provided consent before the samples were collected.
Isolation and culture of canine ADSCs and LTT-tolerant cells
Adipose tissues were obtained from 10 female healthy dogs between 10 and 18 months of age during surgery for spaying by clinical veterinarians. All collected samples were stored at 4°C and processed within 4 h of sampling. ADSCs were isolated from abdominal adipose tissues according to a previously published method [24] with modifications, using the Canine Adipose-Derived Stem Cell Culture Kit (J-ARM Co., Ltd, Osaka, Japan). In brief, fat tissues were incubated in collagenase (FUJIFILM, Tokyo, Japan) solution (2 mg/mL) for 1 h at 37°C, and then centrifuged at 340 g for 5 min. The cell pellet was resuspended in 5 mL Dulbecco's phosphate-buffered saline (DPBS; FUJIFILM) and centrifuged again under the same conditions. Finally, the pellet was resuspended in prewarmed Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum and 1% antibiotic and incubated at 37°C with 5% CO2 and 95% humidity for 48 h. Subsequently, the adherent cells were washed with DPBS and placed in fresh culture medium. Upon reaching 70% confluency, cells were trypsinized, and the cells were subcultured until passage 1. The cells were then cryopreserved, thawed, and cultured at the time of use for this experiment. Following previously described methods [1], expanded ADSCs were exposed under 0.05% trypsin solution for 16 h without CO2 exchange at 37°C. LTT-tolerant cells were collected by centrifugation at 536 g for 15 min, cultured in suspension in a 75T Flask for 5–7 days, and subcultured in adherent according to previously described methods [1].
Flow cytometry
Flow cytometric analysis was performed to evaluate the cell surface markers according to previously described methods [21,25]; 1 × 106 LTT-tolerant cells or ADSCs from four dogs were washed and resuspended in 100 μL FACS buffer and incubated with fluorescent isothiocyanate (FITC)-labeled antibody for CD14 and CD45 phycoerythrin-labeled antibody for CD44, nonlabeled antibody for CD90, SSEA-1, and SSEA-3. Isotype control was used in a negative control sample. Then, the cells were incubated with corresponding secondary antibodies. Data were measured using flow cytometry (10,000 events, Attune™ NxT; Thermo Fisher Scientific, Waltham, MA), repeated twice, and analyzed using the Attune NxT software (Thermo Fisher Scientific). The antibodies are described in Supplementary Table S1.
Induced differentiation of ADSCs and LTT-tolerant cells in vitro
Cells from five dogs were evaluated for differentiation. For osteoblast and adipocyte differentiation, 5 × 104 LTT-tolerant cells (at passage 3) or ADSCs (at passage 2 or 3) were seeded onto each well of a six-well culture plate in canine induction medium for 14 days to osteoblasts and for 21 days to adipocytes. For chondrocyte differentiation, 2 × 105 ADSCs (at passage 2) were seeded into each a well of a 96V-well culture plate by induction medium for 21 days [21,26]. For hepatocyte differentiation, 2 × 104 LTT-tolerant cells (at passage 3) were seeded into each well of a collagen-coated 48-well cell culture plate by the induction medium for 14 days [21]. For neural cell differentiation, 1 × 105 LTT-tolerant cells (at passage 3) were seeded as nonadherent cells in polyHEMA-coated 25T flask in the presence of neural induction medium for 7 days. The cells were then transferred to recombinant laminin-coated PLL 48-well cell culture plate and cultured for another 10 days as adherent cells in the induction medium to differentiate into neurons [21]. Medium change was performed every other day. After induction, the cells were stained with von Kossa staining for osteoblasts, oil Red O staining for adipocytes, Alcian blue staining for chondrocytes, and immunocytochemistry for hepatocytes and neural cells, and repeated twice according to previously described methods [1,26].
Immunocytochemistry for pluripotent stem cell, hepatocytes, and neural cell markers
Cells from five dogs were evaluated for immunocytochemistry. Immunocytochemistry was performed to confirm their pluripotency in LTT-tolerant cells according to previously described methods [1]. LTT-tolerant cells placed in suspension culture at days 5–7 and induced for neural precursor cells were collected by centrifugation, fixed, embedded in OCT compound, and frozen at −80°C. Subsequently, the cryosections were cut to 5 μm thickness using a cryostat (HM525NX7; Thermo Fisher Scientific). LTT-tolerant cells induced for hepatocytes and neural cells in adherent culture were fixed. After washing, the cells were treated with 0.5% Triton™ X-100 and blocked with blocking solution. After another wash, the cells were incubated with primary antibodies overnight at 4°C. Anti-SSEA-1, SSEA-3, NANOG, OCT3/4, SOX2, and TRA-1 antibodies for pluripotency stem cell markers; anti-albumin and alpha 1 fetoprotein antibodies for endodermal lineage; and anti-NESTIN, Musashi-1, NeuroD 2, and MAP-2 antibodies for ectodermal lineage were used as the primary antibodies. The cells were washed again and incubated with secondary antibodies under protection from light at room temperature (R/T) for 2 h. Finally, cells were washed and treated with DAPI for nuclear staining (Sigma-Aldrich, St. Louis, MO). Cells were repeated two times for immunocytochemistry. The primary and secondary antibodies are described in Supplementary Table S1. Images were taken with a fluorescence microscope (Keyence, Osaka, Japan).
Colony-forming unit fibroblast assay
Cells from seven dogs were evaluated for colony-forming unit fibroblast (CFU-F) assay. A total of 500 cells were seeded onto each well of a six-well culture plate for 14 days in a culture medium. After 14 days, the adherent cells were washed with DPBS and fixed with 4% paraformaldehyde phosphate-buffered solution at R/T for 15 min. The cells were stained with 1 mL Giemsa stain solution at R/T for 30 min. The average number of cells in each colony in three wells was determined by counting, and colonies containing >50 cells were scored.
Alkaline phosphatase activity
Cells from five dogs were evaluated for alkaline phosphatase (ALP) assay; 1 × 105 LTT-tolerant cells (days 5–7 in suspension culture) were washed three times with a sufficient volume of saline. These cells in 1 mL of the buffer were stained using a leukocyte alkaline phosphatase kit (Sigma-Aldrich) according to the manufacturer's instructions.
Karyotype analysis
Cells from three dogs were evaluated for karyotype analysis. The karyotype of LTT-tolerant cells was analyzed to examine their influence on LTT according to previously described methods [27]. These cells were cultured in suspension until day 6 and subcultured in an adherent culture. Colcemid (0.1 mg/mL; Thermo Fisher Scientific) was added to the 75T flask for 5 h. The cells were treated with trypsin and collected by centrifugation, and 0.075 M potassium chloride was added in the pellets at 37°C for 15 min. Subsequently, Carnoy's fluid was added. The pellets were dropped to a glass slide, and the number of chromosomes was evaluated every two times and analyzed by Giemsa staining.
Lectins and antibodies
PSL1a lectin (FUJIFILM), which is known to be a marker of the differentiation potential of human MSCs, was labeled using fluorescein labeling kit-NH2 (Dojindo, Kumamoto, Japan) according to the manufacturer's protocol. ADSCs and LTT-tolerant cells in suspension from five dogs were incubated at 4°C for 30 min with FITC-labeled PSL1a lectin (6 μg/mL), according to previously described methods [28]. Cells were washed several times with FACS buffer and measured using flow cytometry, repeated twice, and analyzed using the Attune NxT software.
Telomerase activities
Telomerase reverse transcriptase (TERT), which is correlated with tumorigenic activities, of LTT-tolerant cells in suspension culture and ADSCs in adherent culture at day 7 from five dogs were examined by quantitative reverse transcription-PCR (RT-qPCR) analysis. Canine thyroid adenocarcinoma cells (CTACs) were used as a positive control.
Migration ability
Cells from six or seven dogs were evaluated for migration assay. Migration was assayed in 24-well cell culture companion plates (Corning, Glendale, AZ) with 8-μm pore inserts (Corning), according to previously described methods [29]. LTT-tolerant cells in suspension or ADSCs in adherence at 7 days were collected and washed with DPBS. After that, 1 × 104 LTT-tolerant cells or ADSCs in 300 μL of serum-free medium were seeded onto the apical surface of the insert. Chemoattractants were added to the basal chamber (500 μL of basal medium) as follows: recombinant feline/human stromal cell-derived factor 1β (SDF-1β; R&D Systems, Minneapolis, MN) and sphingosine 1-phosphate (S1P; Sigma-Aldrich). After overnight incubation at 37°C, the cells were washed and wiped out on the top surface of the insert membrane using a cotton swab. After staining with the Diff-Quik stain kit (Sysmex, Hyogo, Japan), the cells were air-dried. Cells outside the insert membrane were evaluated by counting the total number of observations in four different areas using a 10 × objective lens. In another experiment, we stimulated LTT-tolerant cells or ADSCs from eight dogs by tumor necrosis factor (TNF)-α, which is an inflammatory cytokine that stimulates SDF, to clarify the migration process. LTT-tolerant cells in suspension or ADSCs in adherence at 7 days were stimulated by a final concentration of 10 ng/mL TNF-α for 24 h. Subsequently, the stimulated cells were collected by centrifugation and total RNA was isolated.
Inhibition of lymphocyte proliferation
LTT-tolerant cells or ADSCs from nine dogs were evaluated for flow cytometry analysis. Immune suppression assays were performed according to previously described methods [30]. Peripheral blood mononuclear cells (PBMCs) were isolated from five healthy donor dogs by density gradient centrifugation. For further experiments, a portion of the PBMCs were cryopreserved by CELLBANKER® 1 (TAKARA, Shiga, Japan). LTT-tolerant cells or ADSCs were seeded onto each well for 1 day before coculture with PBMCs. Subsequently, the culture media was exchanged, and PBMCs were cocultured for 3 days. Lymphocyte proliferation in a lymphocyte gate with forward-scatter and side-scatter was analyzed using flow cytometry.
Quantitative reverse transcription PCR
Total RNA was isolated using NucleoSpin® RNA Plus kit (TAKARA). cDNA was synthesized using a PrimeScript™ RT Master Mix (Perfect Real Time) (TAKARA). qRT-PCR was performed with the SYBR Green assay using TB Green® Premix Ex Taq™ II (Tli RNaseH Plus; TAKARA) analyzed with a real-time PCR detection system (Thermal Cycler Dice® Real Time System; TAKARA), with β-actin used for normalization. All primers used are listed in Supplementary Table S2.
Statistics
The data were tested for normality in this study. Experimental groups were compared using one-way analysis of variance followed by Fisher's protected–least significance–post hoc analysis [Microsoft® Excel® for Microsoft 365 MSO (16.0.13426.20270)] because that data did not have a normal distribution. Quantitative data are expressed as mean ± standard error. The significance levels were set at P < 0.01 and P < 0.05.
Results
Isolation and characterization of canine ADSCs and LTT-tolerant cells
We morphologically compared canine ADSCs and LTT-tolerant cells to examine their characteristics. ADSCs typically showed a spindle shape (Fig. 1A), had an osteogenic phenotype for 14 days, and adipogenic and chondrogenic phenotypes for 21 days after induction (Fig. 1B). We showed the result of cell survival rate after LTT. About 9.3% ± 0.5% cells in canine ADSCs survived. In this study, we defined the survived cells as LTT-tolerant cells. LTT-tolerant cells in suspension culture formed cell clusters (∼50 μm in diameter), similar to the embryoid body (Fig. 1C). After passaging to adherent culture, LTT-tolerant cells were adherent and exhibited a spindle shape without becoming flattened or large (Fig. 1C). There were no clear morphological differences between ADSCs and LTT-tolerant cells in adherent cultures. We evaluated the CD antigen surface using flow cytometry to examine the established MSC markers. ADSCs expressed for CD44 and CD90 were known as positive markers of human MSCs and those that lacked for CD14 and CD45 were known as negative markers of human MSCs (Fig. 2A). Similar results were obtained for LTT-tolerant cells for these markers (Fig. 2B). However, RT-PCR analysis revealed that just after LTT, the relative messenger RNA (mRNA) expression of FUT9 in LTT-tolerant cells was ∼74 times higher than that in ADSCs without LTT (Fig. 2C). The expression intensity of SSEA-1 was significantly enhanced by LTT (Fig. 2C). On the contrary, the expression intensity of SSEA-3, a known human Muse cell marker, did not change by LTT (Fig. 2C).
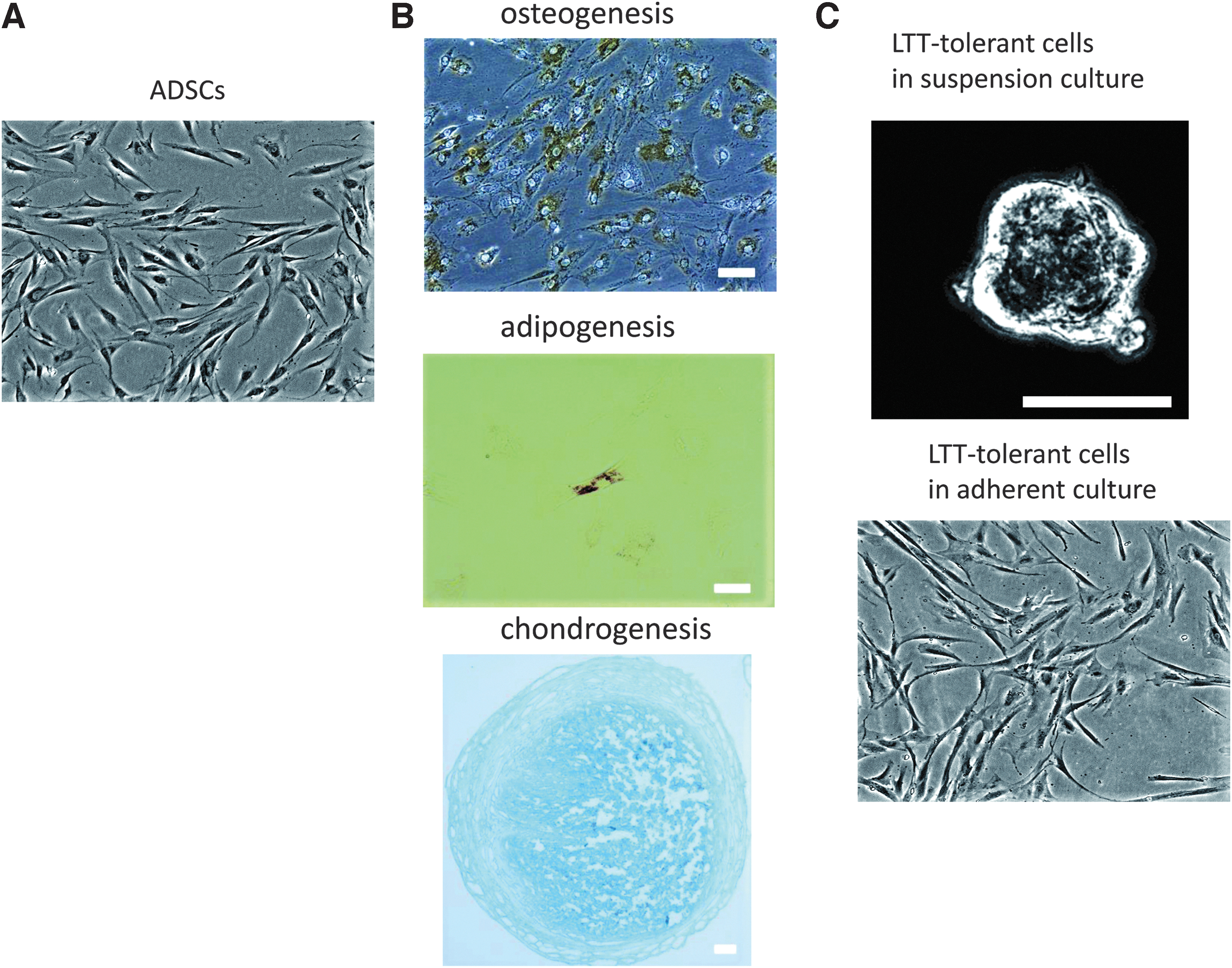
The morphologies of canine ADSCs and LTT-tolerant cells.
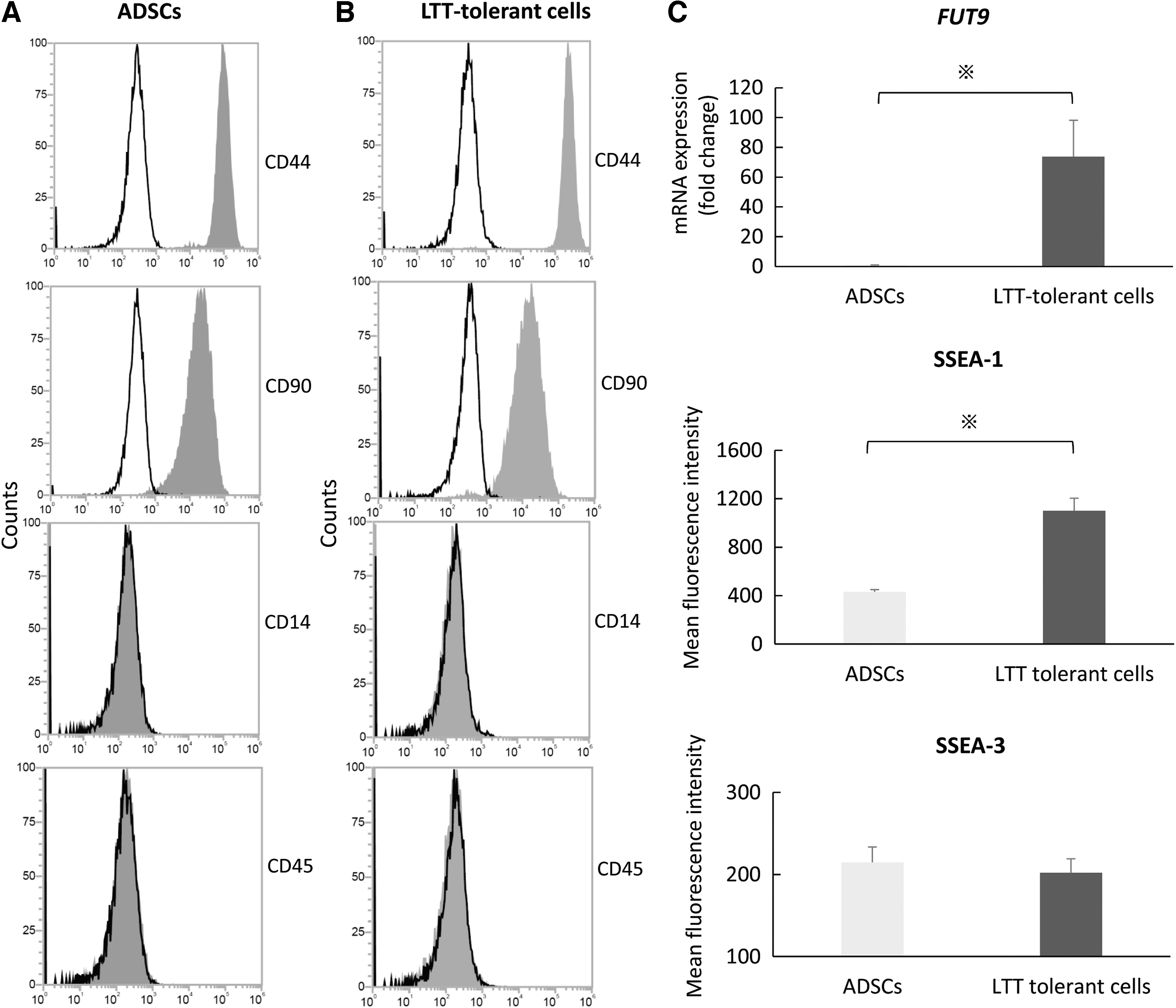
The characteristics of ADSCs and LTT-tolerant cells.
Self-renewal capacity and pluripotency in LTT-tolerant cells
We conducted CFU-F assays in vitro to compare the self-renewal capacity of ADSCs and LTT-tolerant cells. LTT-tolerant cells in suspension generated a significantly (P < 0.01) higher number of CFU-F than ADSCs when assayed 14 days later (Fig. 3A, B).

Self-renewal capacity and pluripotency of LTT-tolerant cells in suspension culture.
We evaluated the cell clusters in LTT-tolerant cells in suspension culture to examine their pluripotency. LTT-tolerant cells were positive for ALP activity (Fig. 3C) and expressed pluripotent stem cell markers such as SSEA-1, SSEA-3, NANOG, SOX2 and OCT3/4, and TRA-1-60 (Fig. 3D).
We analyzed the karyotype of LTT-tolerant cells to examine their influence on LTT. LTT-tolerant cells had a normal 78, XX karyotype, with 38 matched pairs of autosomes at passage 4, with no identifiable gross structural rearrangements (Fig. 4).
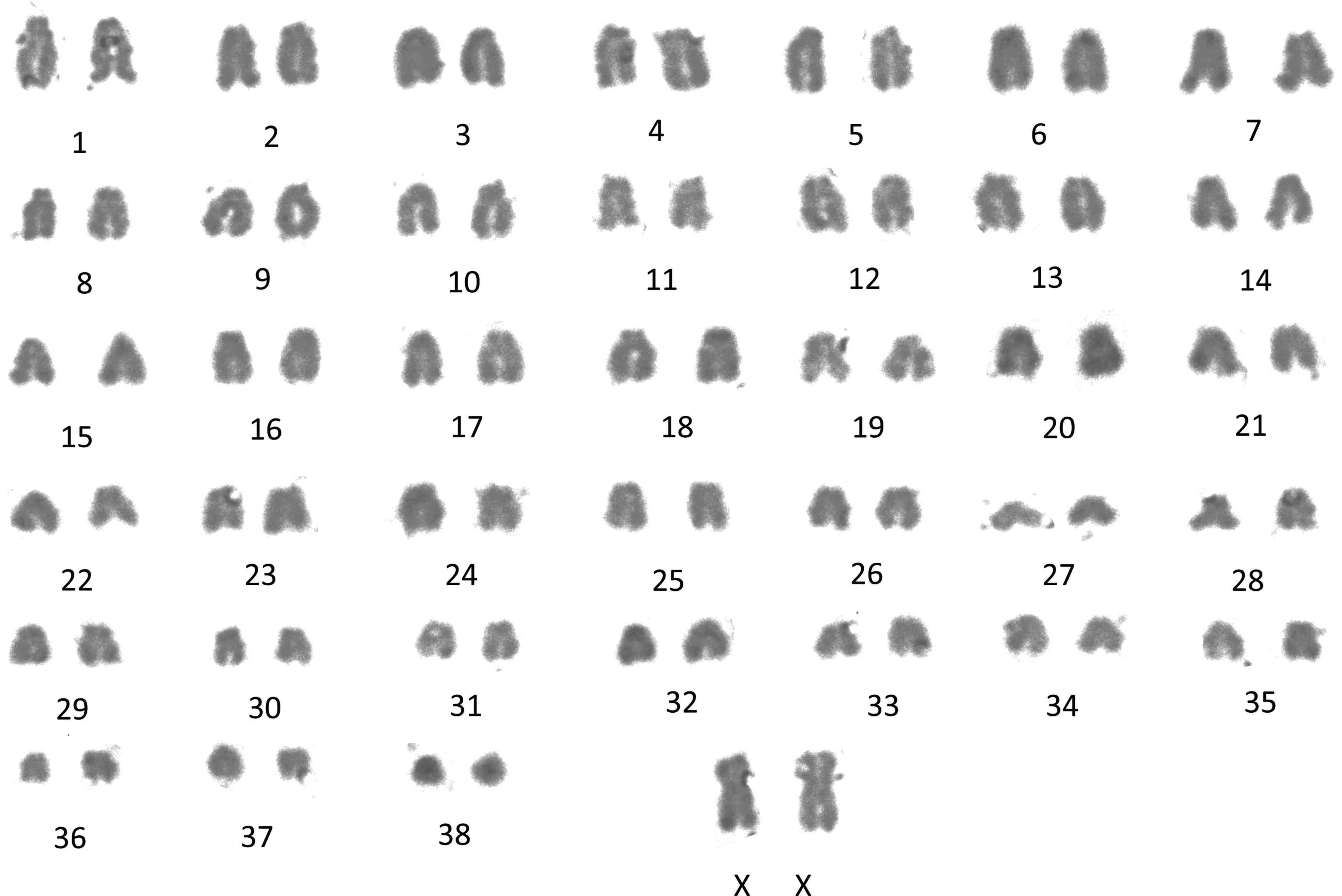
Karyotype analysis of LTT-tolerant cells. Normal: 78, XX with 38 matched pairs of autosomes. Magnification, 1,000 × .
In vitro differentiation of LTT-tolerant cells into the three germ layers
The in vitro differentiation capacity of LTT-tolerant cells was evaluated by von Kossa, Oil Red O, and immunocytochemical staining to examine the three germ layers. Seven days after adipocyte and osteoblast induction, LTT-tolerant cells had osteogenic and adipogenic phenotypes (endodermal), faster than that observed in ADSCs (Fig. 5A, B). Fourteen days after hepatocyte induction, LTT-tolerant cells were positive for albumin and α-fetoprotein (endodermal) (Fig. 5C). Seven days after neural induction, LTT-tolerant cells formed neurospheres and were positive for NESTIN, Musashi-1, and NeuroD 2 (Fig. 5D). Ten days after additional induction, LTT-tolerant cells were positive for MAP-2 (ectodermal) (Fig. 5E).
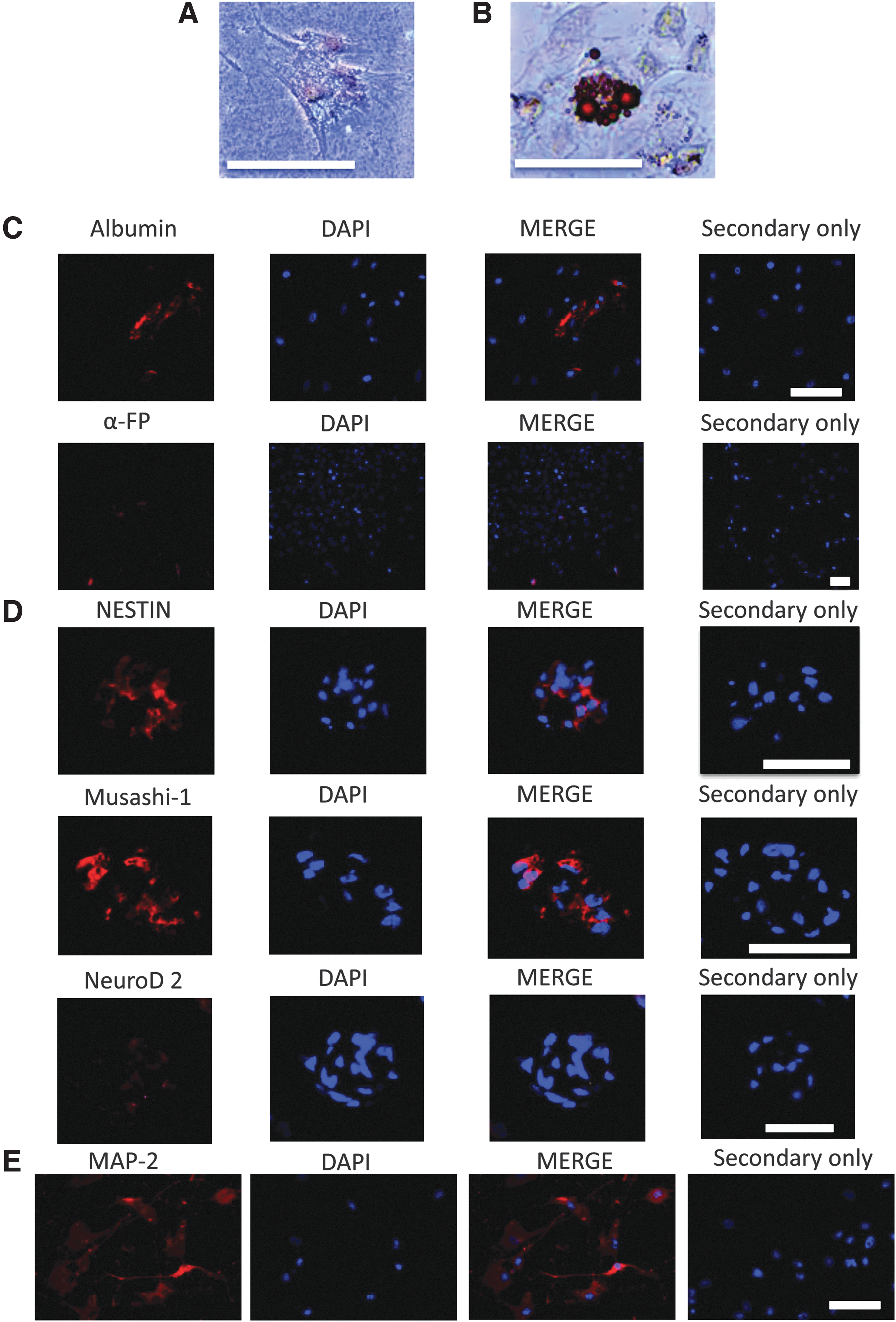
Differentiation of LTT-tolerant cells into the three germ layers in vitro.
Distinct differentiation ability of LTT-tolerant cells
To compare the differentiation potential of ADSCs and LTT-tolerant cells, we conducted an evaluation with PSL1a lectin using flow cytometry. PSL1a lectin produced stronger signals to LTT-tolerant cells than to ADSCs. The expression intensity of FITC-labeled PSL1a lectin was higher than that of ADSCs (Fig. 6A, B). To confirm this result, we morphologically analyzed the differentiation ability of ADSCs and LTT-tolerant cells into osteoblasts and adipocytes. A minority of ADSCs in representative areas differentiated into osteoblasts and adipocytes after induction. On the contrary, the majority of LTT-tolerant cells in representative areas differentiated into osteoblasts and adipocytes under the same differentiation conditions (Fig. 6C).
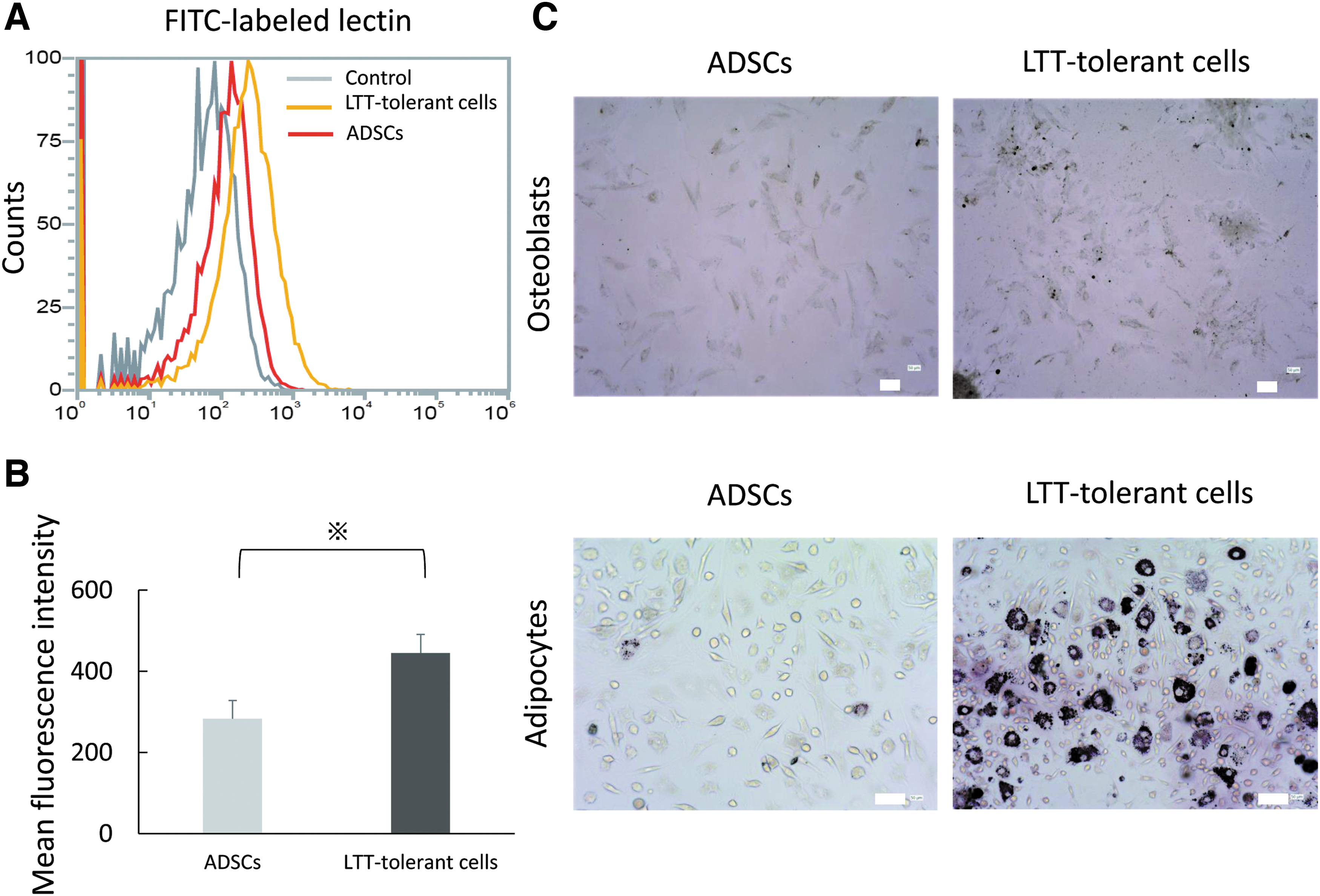
Distinct differentiation ability of LTT-tolerant cells and ADSCs.
In vitro nontumorigenic activities
To confirm tumorigenic activities in LTT-tolerant cells, we examined telomerase activity using RT-PCR analysis. The mRNA expression of TERT in LTT-tolerant cells and ADSCs was significantly lower than in CTAC, the positive control (Fig. 7).

Nontumorigenic characteristics of LTT-tolerant cells. Relative mRNA expression of TERT in LTT-tolerant cells and ADSCs. CTACs were used as the positive control. n = 5. *P < 0.05. CTACs, canine thyroid adenocarcinoma cells; TERT, telomerase reverse transcriptase.
In vitro migration ability
To examine the migration ability of LTT-tolerant cells and ADSCs, we conducted a migration assay and investigated the mRNA expression of the migrated receptors by RT-PCR analysis. Compared with ADSCs, LTT-tolerant cells had migrated significantly to S1P and SDF-1β and were upregulated in a concentration-dependent manner (Fig. 8A). The mRNA expressions of sphingosine-1-phosphate receptor 2 (S1PR2) and C-X-C chemokine receptor type 4 (CXCR-4) were higher in LTT-tolerant cells than in ADSCs. Moreover, the mRNA expression of CXCR-4 was upregulated by TNF-α in both LTT-tolerant cells and ADSCs (Fig. 8B).
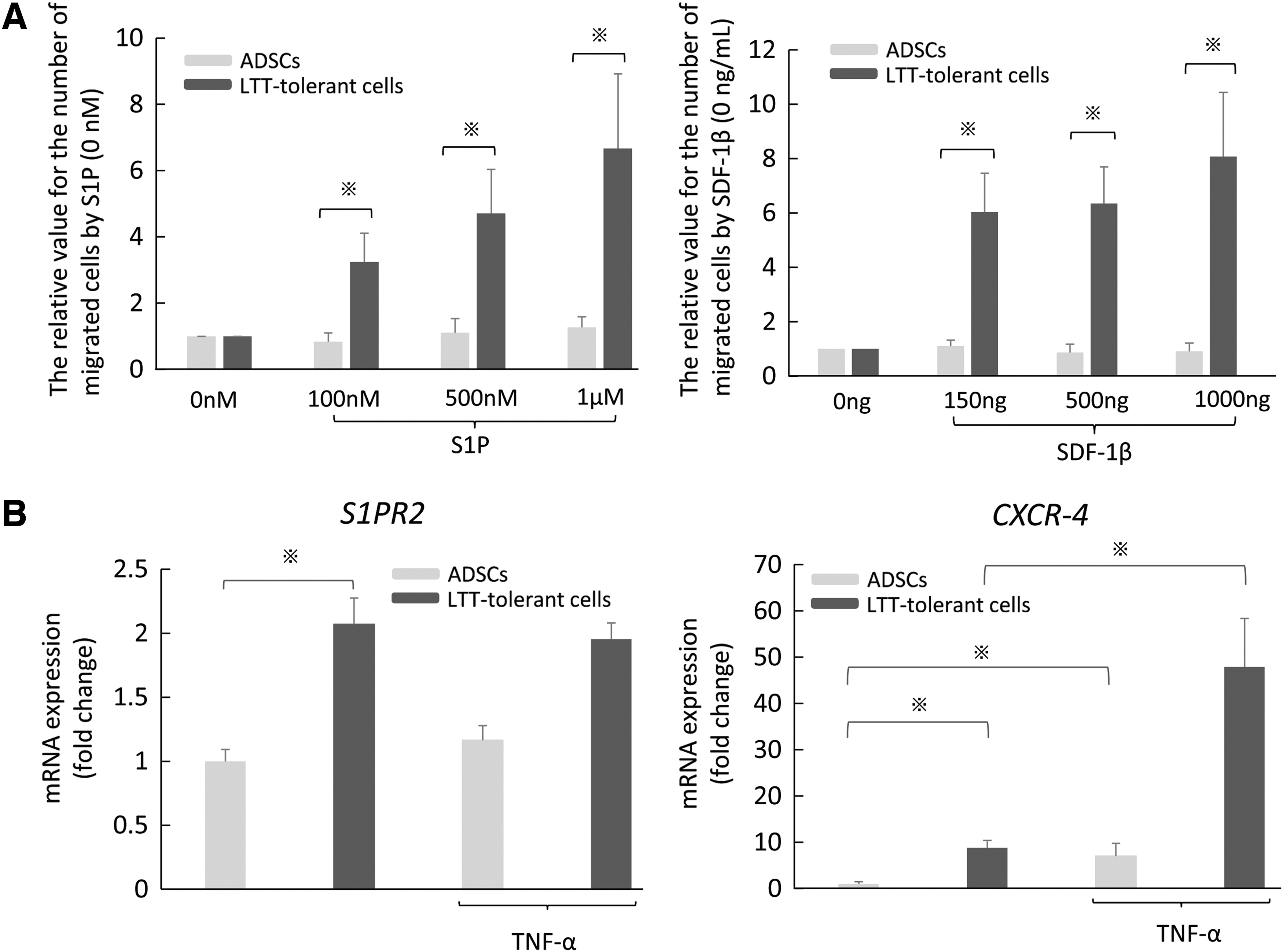
Migration ability of LTT-tolerant cells and ADSCs in vitro.
Inhibition of phytohemagglutinin-stimulated PBMC proliferation in LTT-tolerant cells
To determine the immunomodulatory properties of LTT-tolerant cells, we examined the effects of LTT-tolerant cells and ADSCs on PBMC proliferation by flow cytometry. We found that, compared with ADSCs, LTT-tolerant cells significantly inhibited phytohemagglutinin (PHA)-stimulated PBMC proliferation (Fig. 9).
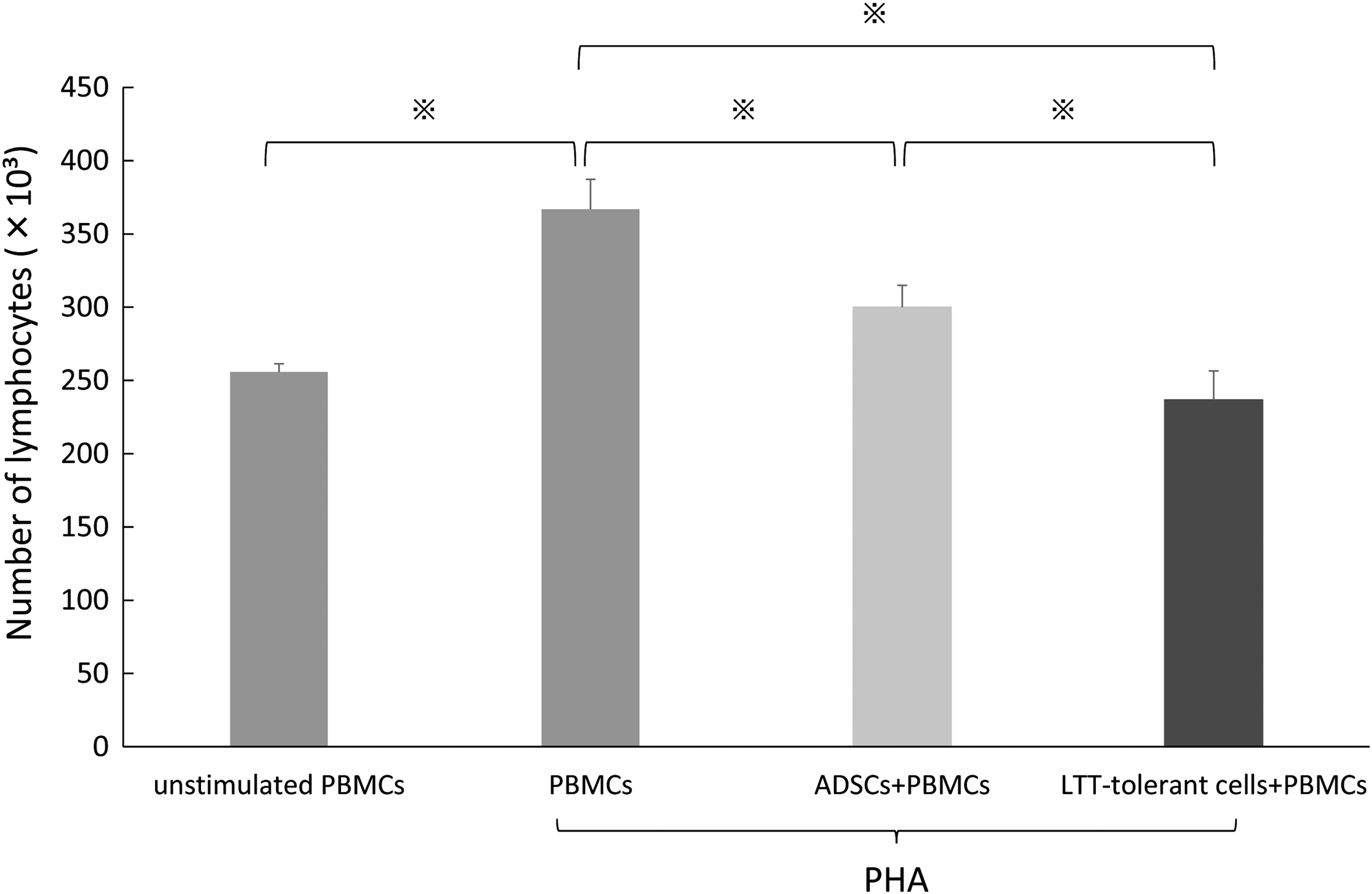
Suppression of PHA-stimulated PBMC proliferation in LTT-tolerant cells and ADSCs. PBMCs were stimulated with PHA and then incubated with LTT-tolerant cells and ADSCs. LTT-tolerant cells had stronger suppressive activity against the proliferation of mitogen-stimulated lymphocytes than that of ADSCs. n = 9. *P < 0.05. PBMCs, peripheral blood mononuclear cells; PHA, phytohemagglutinin.
Discussion
We showed here that LTT-tolerant cells from canine ADSCs had high stem cell potency. This is the first report on the generation of pluripotent stem cells from canine ADSCs. To date, few studies have reported a subpopulation of canine ADSCs by cell sorting. In a previous study, the aldehyde dehydrogenase high-activity (ALDHHi) subpopulation was shown to have a high capacity for adipogenic and osteogenic differentiation [12]. The subpopulation with cystine transporter expression (xCTHi)/CD44+ is reported to have higher proliferation capacity and show higher gene expression of stem cell markers than the xCTLo/CD44+ subpopulation [13]. However, their pluripotency has not been investigated in detail [12,13]. The LTT-tolerant cells we isolated had similar properties to human Muse cells, such as high self-renewal capacity, formation of cell clusters in suspension culture, expression of pluripotent stem cell markers, and differentiation into three germ layers in vitro. However, cells from different species often have unique characteristics that are absent in cells from other species. Further studies are needed to examine whether LTT-tolerant cells from canine ADSCs have more unique characteristics.
Our ADSCs were spindle-shaped, showed adherence to plastic, displayed self-renewal, expressed CD44 and CD90 surface markers, lacked expression of CD14 and CD45, and exhibited adipogenic, osteogenic, and chondrogenic differentiation, similar to canine MSCs [31 –33]. We conducted severe stress stimulation, that is, LTT, to generate cells with high stem cell potency from canine ADSCs, similar to the collection of human Muse cells [1]. The presence of ∼1% Muse cells in human BMSCs using flow cytometry analysis [1,21], and 3% survival of human BMSCs after LTT [1] has been reported. These results suggest that LTT-tolerant cells from human BMSCs are enriched in Muse cells. In addition, Muse cells from human ADSCs were found to be present at 8.8 ± 1.3% in flow cytometry analysis [34]. In this study, LTT-tolerant cells were isolated at 9.3% ± 0.5%. These results indicate that the ratio of LTT-tolerant cells from canine ADSCs might be similar to that of human Muse cells. Further studies should be performed to determine the presence of pluripotent stem cells in canine ADSCs without LTT.
This study revealed that LTT-tolerant cells from canine ADSCs were not distinguishable from ADSCs without LTT in adherent culture, retained the typical surface markers of ADSCs and differentiated into osteocytes and adipocytes. Furthermore, our LTT-tolerant cells had similar properties to human Muse cells, such as being LTT tolerant, possessing self-renewal abilities, being able to form cell clusters in suspension culture, being positive for ALP activity and pluripotency stem cell markers, being able to differentiate into three germ layers, possessing a normal karyotype, and having low telomerase activity in vitro. SSEA-1 and SSEA-3 are well-known as a cell marker of pluripotent stem cell. These markers are effectively used to distinguish pluripotent stem cells from other cells and to track the differentiation process in stem cells and generally disappear once the pluripotent stem cells differentiate. Therefore, they are useful to check whether stem cells maintain the stemness. Of interest, our study also demonstrated that the expression of FUT9 mRNA and the expression intensity of SSEA-1 in LTT-tolerant cells were significantly increased, and that the expression intensity of SSEA-3 in LTT-tolerant cells did not change immediately after LTT. In pigs, it has been reported that SSEA-1 expression enhances reprogramming of cells, which are negative for SSEA-3, in embryonic fibroblasts [35]. In rats, it has also been reported that SSEA-1- and SSEA-3-positive cells are mixed in LTT-tolerant cells from rat BMSCs, and ∼58% of LTT-tolerant cells are double positive for SSEA-1 and SSEA-3 [36]. Our results indicated that SSEA-1- and SSEA-3-positive cells might be mixed in LTT-tolerant cells from canine ADSCs because the expression of SSEA-1 was increased and that of SSEA-3 was slightly decreased. Moreover, LTT-tolerant cells in dogs might be characterized by FUT9 mRNA and SSEA-1, unlike human Muse cell, although the details of the differences in the two pluripotent stem cell markers have not been clarified. Further studies are needed to analyze the ratio of expression of both SSEA-1 and SSEA-3 using a cell sorting. SSEA-3 is a glycolipid, whereas SSEA-1 is a glycoprotein. Therefore, the gene analysis and the investigation in a cell origin using SSEA-1 is possible in LTT-tolerant cells from canine ADSCs. Further studies are necessary using progressive methods such as the analysis of single cell levels in cell sorting by SSEA-1, the formation of cell clusters, and analysis using other FACS antibodies to confirm specific cross-reactivity.
Human Muse cells have been reported to have superior differentiation ability to that of MSCs [22]. Human Muse cells with BODIPY-C16 (which is a lipid marker) were present at 80% ± 4% for 3 days after induction, although ADSCs did not yet possess a signal for the marker, according to a previous report [22]. In this study, we evaluated the differentiation ability of LTT-tolerant cells and ADSCs on PSL1a lectin. The morphology of osteogenic and adipogenic differentiation after induction correlated well with that of PSL1a lectin. These results indicated that, compared with ADSCs like human Muse cells, LTT-tolerant cells had a superior ability to differentiate into mesodermal lineage cells. In addition, human Muse cells have been reported to have spontaneous differentiation ability into the three germ layers in vitro [1,22]. More studies on canine LTT-tolerant cells are necessary to investigate the spontaneous in vitro differentiation ability without induction into three germ layers and into a variety of damaged cells in vivo using animal models of disease.
In the migration of MSCs, the interaction between SDF-1β, which is a small chemotactic cytokine that is often induced by proinflammatory stimulation, such as TNF-α and interleukin-1-produced inflammatory cells and damage tissues, and CXCR-4, which is the receptor of this chemokine in MSCs, has been reported to play an important role [6,37]. Human Muse cells have been reported to have a high migration ability to damaged tissues through S1P-S1PR2 interaction [18]. In our study, LTT-tolerant cells exhibited migration ability to S1P and high expression of S1PR2 mRNA like human Muse cells in vitro. This result indicated that LTT-tolerant cells might have a similar migration mechanism as that of human Muse cells. In addition, LTT-tolerant cells showed migration ability to SDF-1β and high expression of CXCR-4 mRNA, similar to human MSCs in vitro. Overall, these results suggest that the LTT-tolerant cells may have two processes for migration. This ability in LTT-tolerant cells can be advantageous in case of IV administration for clinical applications similar to human Muse cells. Further studies are necessary to investigate migration abilities to damaged tissues in vivo after IV administration of LTT-tolerant cells using animal models of disease.
The immunosuppressive ability of Muse cells has not yet been reported. In one previous report, human Muse cells were described as possessing immunosuppressive capacity, such as the inhibition of lymphocyte and macrophage proliferation by producing transforming growth factor-β1 [38]. The immunosuppressive function of Muse cells is expected to play a useful role in MSCs. In this study, LTT-tolerant cells were superior to ADSCs in the immunosuppression ability of PHA-stimulated PBMC proliferation. We believe that LTT-tolerant cells will be useful for the treatment of immune-mediated diseases. MSCs have been reported to have immunosuppressive ability to immune cells. In T cells, MSCs suppress the proliferation of CD4+ and CD8+ T cells and promote the proliferation of Treg. In B cells, MSCs suppress the production of antibodies and their proliferation. In NK cells, MSCs suppress the proliferation and production of cytokine. In macrophages and DCs, MSCs suppress maturation [10]. Further research in LTT-tolerant cells is necessary to investigate their interaction with immune cells.
In conclusion, we successfully generated LTT-tolerant cells with high stem cell potency from ADSCs of adult dogs. These cells had similar properties as human Muse cells, that is, they were LTT tolerant, nontumorigenic, expressed pluripotent stem cell markers, differentiated into the three germ layers, and had superior migration and immunosuppressive capacities compared with ADSCs. We believe that these cells are not only useful cell sources for clinical application in veterinary regenerative medicine but also meaningful for human medicine.
Footnotes
Acknowledgments
The authors thank Nippon Lighthouse Guide Dog Training Center (Osaka, Japan) for providing blood samples and fat tissues. The authors also thank Editage for English language editing.
Author Disclosure Statement
Y.T. is a Chief Executive Officer and a stock-holder of J-ARM Co., Ltd., the company that markets the cell culture kit that have been used in the study reported herein. K.M. and Y.I. are employees of J-ARM Co., Ltd. The other authors declare no conflicts of interest.
Funding Information
This study was supported by the Japan Society for the Promotion of Science (Grant No.: 18K19274).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.