
Abstract
Mesenchymal stem cells (MSCs) represent a population of adult stem cells that have potent immunoregulatory, anti-inflammatory, and antiapoptotic properties. In addition, they have ability to migrate to the site of inflammation or injury, where they contribute to the regeneration and healing process. For these properties, MSCs have been used as therapeutic cells in several models, including treatment of damages or disorders of the ocular surface. If the damage of the ocular surface is extensive and involves a limbal region where limbal stem cell reside, MSC therapy has been proved as the effective treatment approach. Although the anti-inflammatory properties of MSCs have been well characterized, mechanisms of antiapoptotic action of MSCs are not well recognized. Using a chemically damaged cornea in a mouse model, we showed that the injury decreases expression of the gene for antiapoptotic molecule Bcl-2 and increases the expression of proapoptotic genes Bax and p53. These changes were attenuated by local transplantation of MSCs after corneal damage. The antiapoptotic effect of MSCs was tested in an in vitro model of co-cultivation of corneal explants with MSCs. The apoptosis of corneal cells in the explants was induced by proinflammatory cytokines and was significantly inhibited in the presence of MSCs. The antiapoptotic effect of MSCs was mediated by paracrine action, as confirmed by separation of the explants in inserts or by supernatants from MSCs. In addition, MSCs decreased the expression of genes for the molecules associated with endoplasmic reticulum stress Atf4, Bip, and p21, which are associated with apoptosis. The results show that MSCs inhibit the expression of proapoptotic genes and decrease the number of apoptotic cells in the damaged corneas, and this action might be one of the mechanisms of the therapeutic action of MSCs.
Introduction
Injury of the cornea is associated with a degenerative process in the corneal epithelium and apoptosis of the corneal epithelial cells. Among the most important factors inducing the apoptosis of corneal cells are the activation of endoplasmic reticulum (ER) stress-induced molecules, a local inflammatory reaction, and the production of several proinflammatory cytokines by corneal and immune cells. It has been shown that cells of the damaged cornea produce proinflammatory molecules, such as interleukin-lα (IL-1α), IL-1β, and tumor necrosis factor-α (TNF-α) [1,2]. It has been demonstrated that increased levels of proinflammatory cytokines lead to apoptosis in in vivo and in vitro models [3,4].
Chronic inflammation of the ocular surface is associated with ER stress, which also promotes the induction of apoptosis [5]. Simultaneously, the site of injury is infiltrated with cells of the immune system, which produce various chemokines and cytokines that can potentiate the inflammatory and apoptotic reactions and attract other immune cells to the site of injury. Therefore, the inhibition of a local inflammatory reaction and alleviation of apoptosis might be a promising approach for the treatment of corneal damages and for the support of corneal epithelium regeneration. In this respect, stem cell-based therapy has proven to be a perspective treatment for severe ocular disorders, especially in cases where the limbal region is destroyed and the endogenous limbal stem cells, which are responsible for corneal regeneration, are missing or are nonfunctional.
Among the various stem cell types that have been tested so far, mesenchymal stem cells (MSCs) turned out to be a perspective candidate. These cells have potent immunoregulatory, secretory, differentiation, and antiapoptotic properties [6 –8]. Furthermore, MSCs can be obtained relatively easily from the patient, and thus can be used as own (autologous) cells. In addition, MSCs are the producers of numerous growth factors such as epidermal growth factor (EGF), fibroblast growth factor (FGF), insulin-like growth factor 1 (IGF-1), hepatocyte growth factor (HGF), or transforming growth factor-β (TGF-β), which contribute to corneal healing and regeneration [9 –11].
Therefore, due to their anti-inflammatory properties, secretion of growth factors, and antiapoptotic properties, MSCs are a perspective candidate for the stem cell-based therapy of corneal injuries or disorders. Although the MSC-based therapy for corneal injuries or diseases has not been approved for clinical use, the data from experimental models are very encouraging, and MSCs have been successfully used for the treatment of corneal injuries or limbal stem cell deficiencies in various models in mice, rats, and rabbits [12 –15].
While the anti-inflammatory properties of MSCs have been well described in various models [6,14], less information is available about the mechanisms of the antiapoptotic action of MSCs. Using an experimental model of a chemically injured cornea, we hereby demonstrated that corneal damage induces changes in the expression of genes associated with apoptosis and the transplantation of MSCs alleviated these changes. To analyze the mechanisms of the antiapoptotic effect of MSCs, we established an in vitro model of co-cultivation of corneal explants in the presence of MSCs. The apoptosis in the explants was induced by proinflammatory cytokines and was decreased in the presence of MSCs. Finally, we demonstrated a paracrine mechanism of the antiapoptotic action of MSCs.
Materials and Methods
Animals
Female BALB/c mice 10–16 weeks of age were used in the experiments. The animals were obtained from the Institute of Molecular Genetics of the Czech Academy of Sciences, Prague. The use of animals was approved by the Local Ethical Committee of the Institute of Experimental Medicine of the Czech Academy of Sciences, Prague.
Isolation and cultivation of MSCs
MSCs were isolated from the femurs and tibias of the mice. The bone marrow was flushed out and a single-cell suspension was prepared by homogenization. The cells were seeded in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO) containing 10% fetal calf serum (Gibco BRL, Grand Island, NY), antibiotics (100 U/mL penicillin and 100 mg/mL streptomycin; Sigma-Aldrich), and 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid buffer (Sigma-Aldrich) in 75 cm2 tissue culture flasks (techno plastic products; Trasadingen, Switzerland). After 48 h of incubation, the nonadherent cells were washed out and the adherent cells were cultured at 37°C in an atmosphere of 5% CO2 with a regular exchange of the medium and passaging of the cells to maintain their optimal concentration. The cells were harvested after the third passage approximately after 2 weeks of cultivation with 1 mL of 0.5% trypsin (Sigma-Aldrich) for 5 min and by gently scraping.
Purification and characterization of MSCs
The suspension of MSCs was incubated for 15 min with CD11b and CD45 Microbeads (Miltenyi Biotec, Belgisch Gladbach, Germany) according to the manufacturer's instructions. CD11b− and CD45− cells were isolated by a magnetic activated cell sorter (AutoMACS; Miltenyi Biotec). Purified cells were washed in phosphate-buffered saline (PBS) containing 0.5% of bovine serum albumin and were incubated for 30 min with anti-mouse monoclonal antibodies: allophycocyanin (APC)-labeled anti-CD44 (clone IM7; BD PharMingen, San Jose, CA), phycoerythrin (PE)-labeled anti-CD73 (cloneTY/11.8; eBioscience, San Diego, CA), PE-labeled anti-CD105 (clone TY/11.8; eBioscience), APC-labeled anti-CD11b (clone M1/70; BioLegend, San Diego, CA), and fluorescein isothiocyanate-labeled anti-CD45 (clone 30-F11; BioLegend). Cells stained with PE-labeled rat IgG2a (clone RTK2758; BioLegend), APC-labeled rat IgG2b (clone RTK4530; BioLegend), or fluorescein isothiocyanate-labeled rat IgG2b (clone RTK4530; BioLegend) were used as negative controls. Dead cells were stained using Hoechst 33258 fluorescent dye (Invitrogen, Carlsbad, CA) added to the samples 10 min before the flow cytometry analysis. Data were collected using an LSRII cytometer (BD Biosciences, Franklin Lakes, NJ) and analyzed using FlowJo software (Tree Star, Ashland, OR). The ability of MSCs to differentiate to adipocytes and osteoblasts was tested as we described [16].
Preparation of supernatants from stimulated MSCs
Purified MSCs were cultured in a concentration of 2 × 105 cells per well in 24-well tissue culture plates (TPP) in 1 mL of complete DMEM. Cells were stimulated by a mixture of proinflammatory cytokines IL-1β, interferon-γ (IFN-γ), and TNF-α (purchased from Peprotech, Rocky Hill, NJ), each cytokine in a final concentration 10 ng/mL. After a 48-h incubation, MSCs were washed several times with an excess of medium to remove added cytokines and cultured for an additional 48 h in a fresh culture medium without cytokine stimulation. Supernatants were harvested and the cell debris was removed by centrifugation (8 min, 2000 g) and stored at −80°C.
Cultivation of MSCs with cytokines in vitro
Purified MSCs were cultured in 24-well TPP with 1 mL of complete DMEM in a concentration 1 × 105 cells per well. Cells were stimulated by proinflammatory cytokines IL-1β, IFN-γ, and TNF-α (a cocktail of cytokines, each in a final concentration of 10 ng/mL). After a 48-h cultivation, the cells were harvested and tested for the expression of genes for immunoregulatory molecules and growth factors.
In vivo model of chemically induced corneal injury
A model of a chemically impaired ocular surface established in our laboratory [15,17] was used to study the expression of genes associated with apoptosis. In brief, the mice were anesthetized by an intramuscular injection of 1:1 mixture of xylazinum hydrochloridum 2% (0.175 mL; Rometar; Spofa, Prague, Czech Republic) and ketaminum hydrochloridum 5% (0.175 mL; Narkamon; Spofa). The right corneas of the anesthetized mice were treated with 0.25 N sodium hydroxide (NaOH) using 8 μL of NaOH on the cornea-size filter paper, attached for 20 s on the cornea. The eye was rinsed with an excess of PBS. To test the effect of MSCs on gene expression in the damaged cornea, MSCs were transferred onto the damaged eye surface using a nanofiber scaffold, as we have described in detail elsewhere [15,17]. The corneas were excised 7 days after corneal injury and immediately transferred into 500 μL of TRI Reagent® [Molecular Research Centre (MRC), Cincinnati, OH] and stored at −80°C.
Induction of apoptosis in corneal explants
The mice were sacrificed, and the central cornea (without limbus) was excised from the enucleated eye. Corneal explants were cultured in 24-well TPP in 1 mL of complete DMEM for 48 h unstimulated or stimulated with IL-1β, IFN-γ, and TNF-α in a concentration 10 ng/mL or with H2O2 in a final concentration of 50 μM (multiple concentrations were tested to establish an apoptosis model in stimulated corneas). H2O2 represents an apoptosis inducer [18] and was used as a positive standard during establishment of the model.
Cultivation of corneal explants with MSCs or MSC supernatant
MSCs (1 × 105 cells/well) were seeded in 24-well TPP and cultured in 500 μL of complete DMEM to adhere. After a 24-h incubation, the cells were washed with an excess of medium and 1 mL of fresh complete DMEM was added. The corneal explants were cultured unstimulated or stimulated, with proinflammatory cytokines (IL-1β, IFN-γ, and TNF-α) or with previously seeded MSCs directly, in insert (NUNC CC Insert, 0.4 μm PC; Thermo Fisher Scientific Nunc A/S, Roskilde, Denmark) or with MSC supernatant (500 μL of previously prepared supernatant and 500 μL of fresh complete DMEM).
Production of IL-6, IGF-1, and HGF by cytokine-stimulated or cytokine-pretreated MSCs
MSCs were stimulated with IL-1β, IFN-γ, and TNF-α and supernatants were harvested after a 48-h incubation. The cells were washed several times with the excess of medium and the cells were incubated for another 48 h in a fresh medium without cytokines. Supernatants obtained after the first stimulation, and after prolonged cultivation, were tested for the presence of IL-6, HGF, and IGF-1 using enzyme-linked immunosorbent assay kits purchased from R & D Systems (Minneapolis, MN).
Detection of apoptotic corneal cells by flow cytometry
The excised cornea was cultured for 48 h untreated or with proinflammatory cytokines, cut into small pieces, and digested with collagenase II (Sigma-Aldrich) in a concentration of 1 mg/mL in Hank's balanced salt solution for 50 min at 37°C. The digestion was stopped by the addition of an excess of complete DMEM. The cell suspension was centrifuged (8 min, 250 g) and single cells were washed in PBS. The cells were stained for Annexin V using an Annexin V apoptosis detection kit (Apronex, Jesenice, Czech Republic) according to the manufacturer's protocol. Dead cells were discriminated using Hoechst 33258 fluorescent dye (Sigma-Aldrich), added to the samples 15 min before the flow cytometry analysis. Data were collected using an LSRII cytometer (BD) and analyzed using FlowJo software (LLC, Ashland, OR).
Detection of gene expression by real-time polymerase chain reaction
The expression of genes in the cornea and MSCs was detected using real-time polymerase chain reaction (RT-PCR). Total RNA was extracted by TRI reagent (MRC) according to the manufacturer's instructions. The total RNA (1 μg) was treated by deoxyribonuclease I (DNase I; Promega, Madison, WI) in a DNase I buffer (Promega) and used for reverse transcription. The first cDNA strand was synthesized by random primers (Promega) using M-MLV reverse transcriptase (Promega) in a total reaction volume of 25 μL. Quantitative RT-PCR was performed by StepOnePlus Real-Time PCR System (Applied Biosystems, Foster City, CA) using SYBR green (Applied Biosystems). The primers used for amplification are shown in Table 1. The quantitative PCR parameters included denaturation at 95°C for 3 min, followed by 40 cycles at 95°C for 20 s, annealing at 60°C for 30 s, and elongation at 72°C for 30 s. Fluorescence data were collected at each cycle after the elongation step at 80°C for 5 s. The relative gene expression was analyzed using StepOne Software 2.3 (Applied Biosystems). A relative quantification model was applied to calculate the expression of the target gene in comparison to glyceraldehyde 3-phosphate dehydrogenase gene (GAPDH).
Sequences of Oligonucleotides Used in Real-Time Polymerase Chain Reaction
Statistical analysis
The results are expressed as the mean ± standard deviation. Comparisons between the two groups were analyzed by Student's t-test or in the case of multiple comparisons by analysis of variance (ANOVA). A value of P < 0.05 was considered statistically significant.
Results
Phenotypic characterization of MSCs
The phenotype of MACS-separated MSCs was characterized by flow cytometry. The cells were positive for CD44, CD73, and CD105 and negative for hematopoietic markers CD11b and CD45 (Fig. 1). The cells had typical fibrocyte-like morphology, were adherent to a plastic surface, and were able to differentiate to adipocytes and osteoblasts, as we have described elsewhere [19].
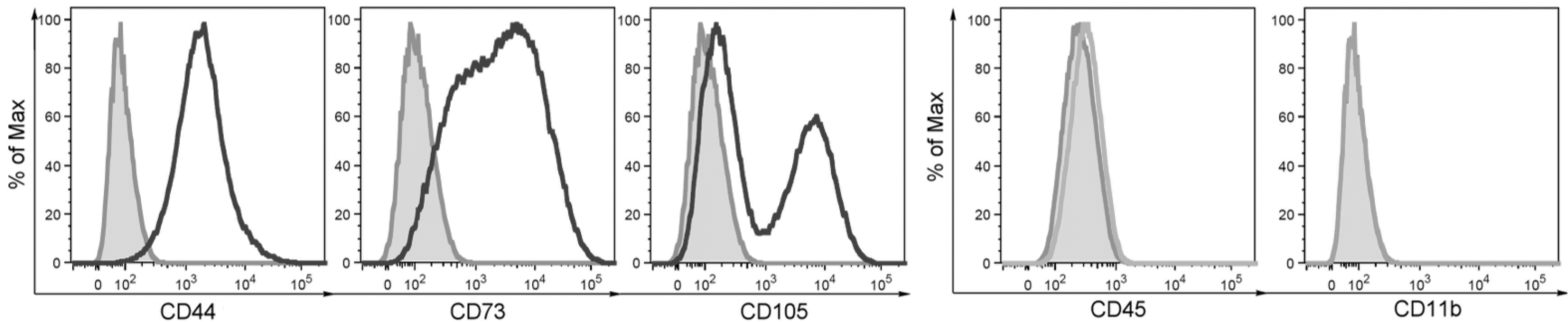
Phenotypic characterization of purified MSCs. The cells were separated by MACS and the expression of CD44, CD73, CD105, CD11b, and CD45 markers was assessed by flow cytometry. One of three similar experiments is shown. MACS, magnetic activated cell sorter; MSC, mesenchymal stem cell.
The production of immunomodulatory molecules and growth factors by MSCs
To characterize the antiapoptotic potential of MSCs, the cells were cultured for 48 h unstimulated, or in the presence of a mixture of proinflammatory cytokines IL-1β, IFN-γ, and TNF-α, which can be detected in corneal injury. The expression of genes for immunomodulatory molecules cyclooxygenase-2 (COX-2), indolamine 2,3-deoxygenase (IDO), IL-6, and programmed death-ligand 1 (PD-L1), and IGF-1 was significantly increased in the presence of proinflammatory cytokines. On the contrary, the expression of genes for FGF, HGF, and TGF-β was decreased after stimulation with cytokines (Fig. 2).
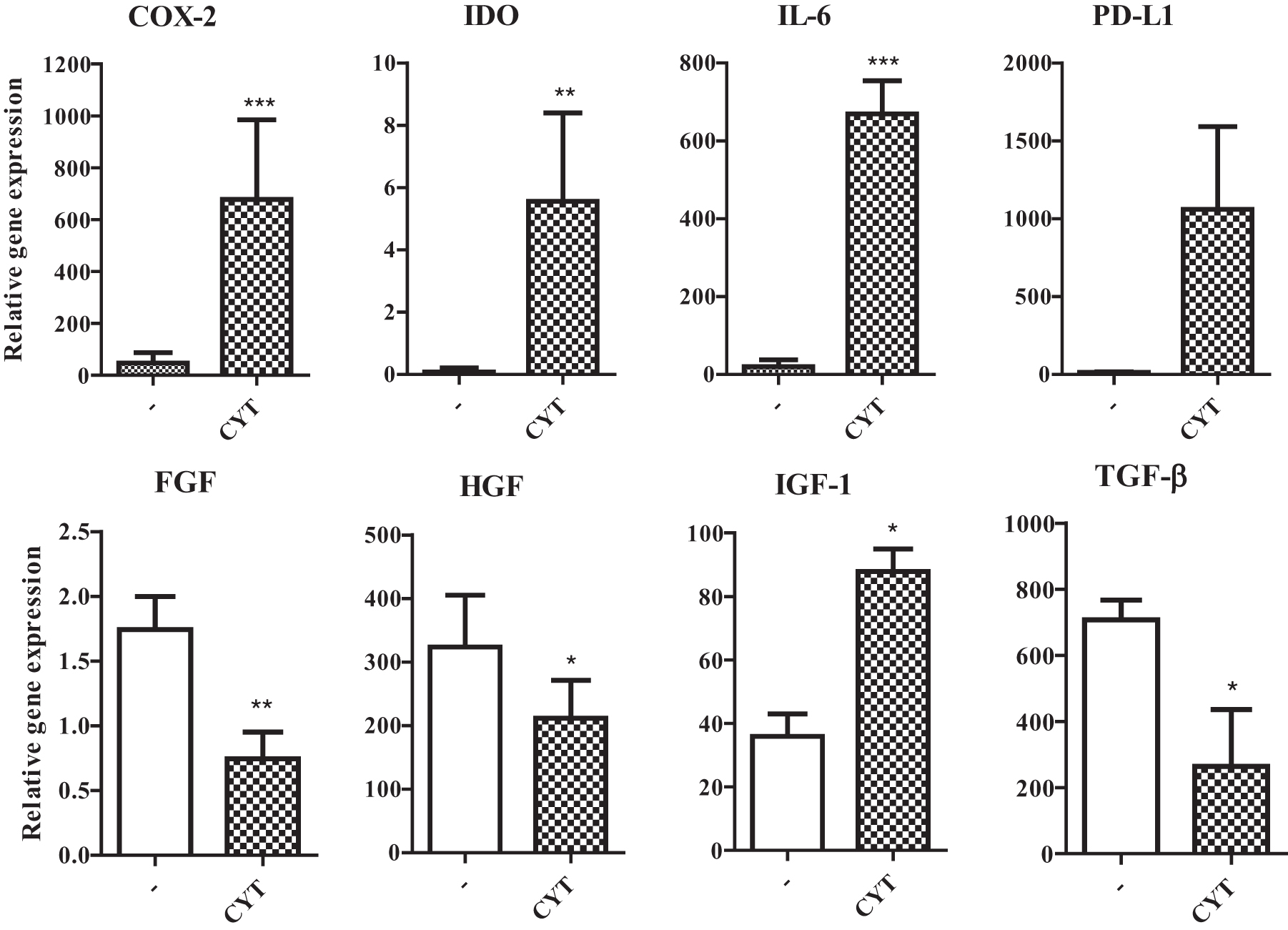
The expression of genes for immunomodulatory molecules and growth factors by MSCs. MSCs were cultured unstimulated (−) or stimulated with a mixture of proinflammatory CYT IL-1β, TNF-α, and IFN-γ. The expression of genes for COX-2, IDO, IL-6, PD-L1, bFGF, HGF, IGF-1, and TGF-β was determined by RT-PCR. Each bar represents the mean + SD from three independent determinations. Values with asterisks are statistically different (*P < 0.05, **P < 0.01, ***P < 0.001) from unstimulated MSCs (−). bFGF, basic fibroblast growth factor; COX-2, cyclooxygenase-2; CYT, cytokines; HGF, hepatocyte growth factor; IDO, indolamine 2,3-deoxygenase; IFN-γ, interferon-γ; IGF-1, insulin-like growth factor 1; IL, interleukin; PD-L1, programmed death-ligand 1; RT-PCR, real-time polymerase chain reaction; SD, standard deviation; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-α.
The concentrations of IL-6, HGF, and IGF-1 proteins were measured in supernatants after a 48-h cultivation of MSCs with proinflammatory cytokines, and after a 48-h consequent cultivation of MSCs in fresh medium without cytokines. After primary stimulation, the concentrations of IL-6 and IGF-1 were significantly increased, while the concentration of HGF was decreased. This pattern of cytokine production was preserved, if the cell cultures were washed and the cells were cultured for another 48 h without cytokines (Fig. 3) for the preparation of supernatants to test the possible paracrine effects of MSCs on the expression of genes associated with apoptosis.
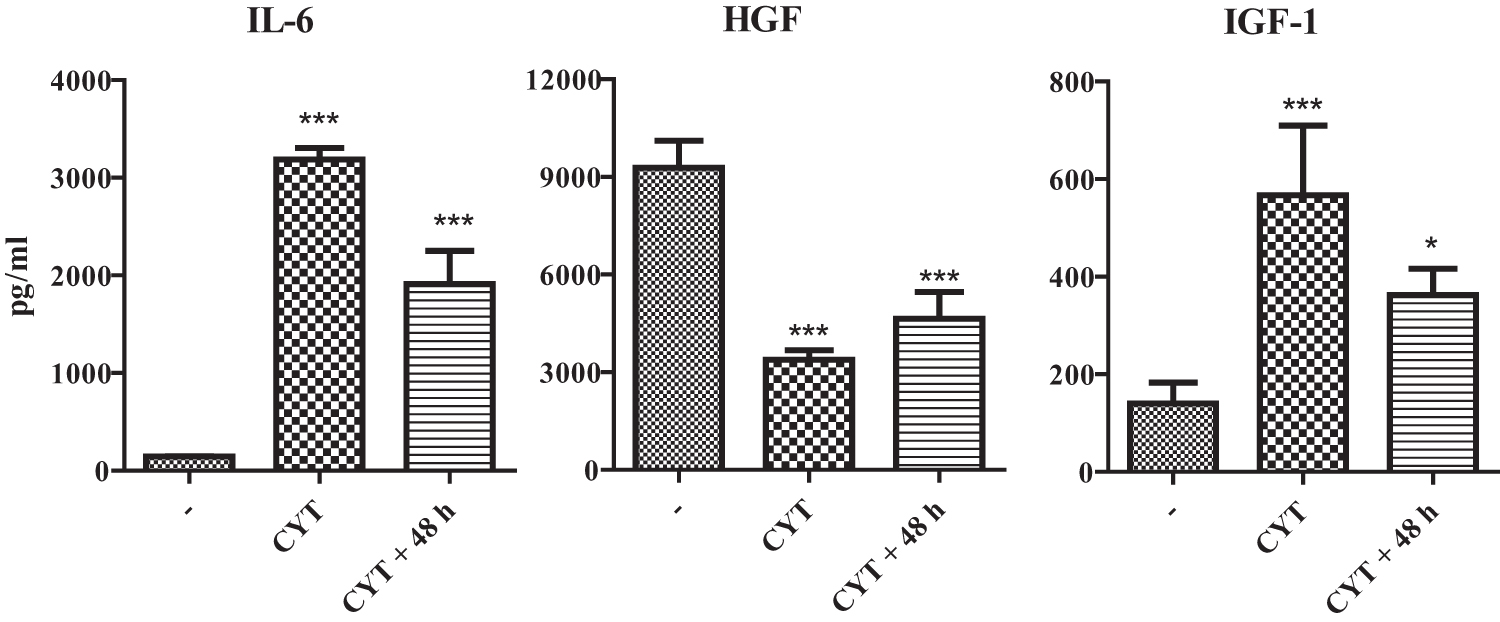
Production of IL-6, HGF, and IGF-1 by MSCs. MSCs were cultured unstimulated (−), stimulated for 48 h with a mixture of proinflammatory CYT or stimulated with CYT, and then washed and cultured for another 48 h without CYT (CYT +48). The production of IL-6, HGF, and IGF-1 was measured by ELISA. Each bar represents the mean + SD from five independent determinations. Values with asterisks are statistically different (*P < 0.05, ***P < 0.001) from unstimulated MSCs (−). ELISA, enzyme-linked immunosorbent assay.
Expression of genes associated with apoptosis in injured corneas and in corneas treated with MSCs
The ocular surface was chemically burned and treated with MSCs on a nanofiber scaffold. The scaffold was removed 3 days after the injury and the corneas were excised on day 7 and analyzed by RT-PCR. As demonstrated in Fig. 4, the expression of gene for Bcl-2 was significantly decreased in injured corneas and this decrease was significantly inhibited after treatment with MSCs. On the contrary, the expression of genes for Bax and p53 was increased in injured corneas and this increase was diminished in corneas treated with MSCs (Fig. 4).

Antiapoptotic effect of MSCs on chemically burned cornea. Control eyes (C) were left untreated, chemically burned corneas were treated with NaOH for 20 s without additional treatment (NaOH) or were treated by nanofiber scaffold without MSCs (NANO) or with nanofiber scaffold with MSCs (MSC). The expression of genes for Bcl-2, Bax, and p53 was determined by RT-PCR. Each bar represents the mean + SD from six mice. Values with asterisks are statistically different (*P < 0.05, **P < 0.01) from control untreated corneas.
Expression of apoptotic genes in corneal explants cultured in the presence of proinflammatory cytokines
To confirm the published observations that proinflammatory cytokines induce apoptosis in corneal cells [3,4], the corneal explants were cultured with IL-1β, IFN-γ, and TNF-α, or with H2O2 as an inductor of apoptosis [18]. After a 48-h cultivation, the expression of genes for antiapoptotic molecule Bcl-1 and for proapoptotic molecules Bax and p53 was determined by RT-PCR. As demonstrated in Fig. 5, the expression of Bcl-2 gene was significantly inhibited in the presence of proinflammatory cytokines or H2O2, while the expression of Bax and p53 genes was increased in the presence of both proinflammatory cytokines and H2O2 (Fig. 5).
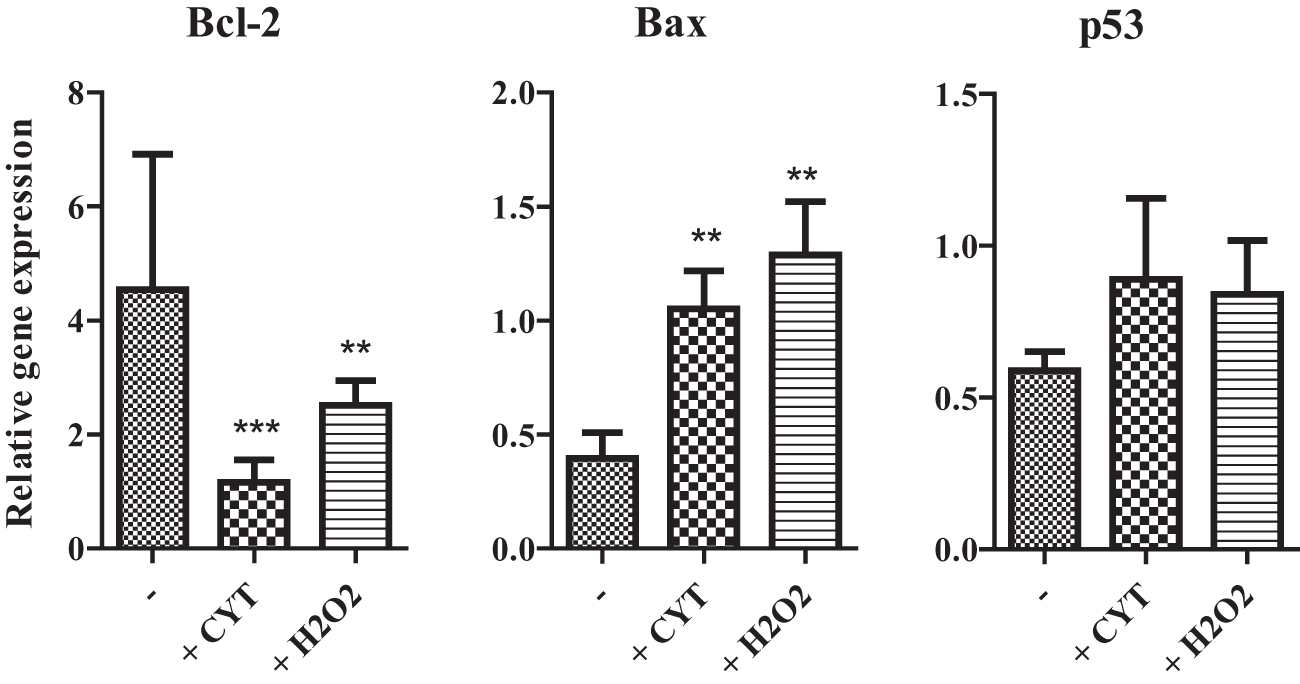
Expression of genes for proapoptotic and antiapoptotic molecules in the corneas cultured with CYT or H2O2. Corneal explants were cultured for 48 h without (−) or with proinflammatory CYT IL-1β, TNF- and IFN-γ (+CYT), or were treated with H2O2 (+H2O2). The expression of genes for Bcl-2, Bax, and p53 was determined by RT-PCR. Each bar represents the mean + SD from five independent determinations. Values with asterisks are statistically different (**P < 0.01, ***P < 0.005) from unstimulated corneas.
The effect of MSCs on the expression of genes for molecules associated with apoptosis in the corneal explants
The corneal explants were cultured untreated, with proinflammatory cytokines, with cytokines and MSCs, either directly or in inserts, and with the supernatant from MSCs to test the antiapoptotic properties of MSCs. After a 48-h co-cultivation, the corneal explants were transferred to a TRI reagent, and the expression of genes for molecules associated with apoptosis or with ER stress was determined by RT-PCR. The expression of gene for antiapoptotic molecule Bcl-2 was decreased after the stimulation by proinflammatory cytokines in comparison with the untreated explants. This decrease was inhibited, if the explants were stimulated in the presence of MSCs, either directly or in insert, or even in the presence of supernatant from MSCs (Fig. 6A). The expression of proapoptotic Bax and p53 genes was increased in the presence of proinflammatory cytokines (in comparison with the untreated explants) and this enhanced expression was diminished in the presence of MSCs or supernatants from MSCs (Fig. 6B). Furthermore, the expression of genes for molecules associated with ER stress (Atf4, Bip, and, P21) was enhanced after cultivation of the corneal explants with proinflammatory cytokines, and this increase was significantly inhibited by MSCs (Fig. 6C).
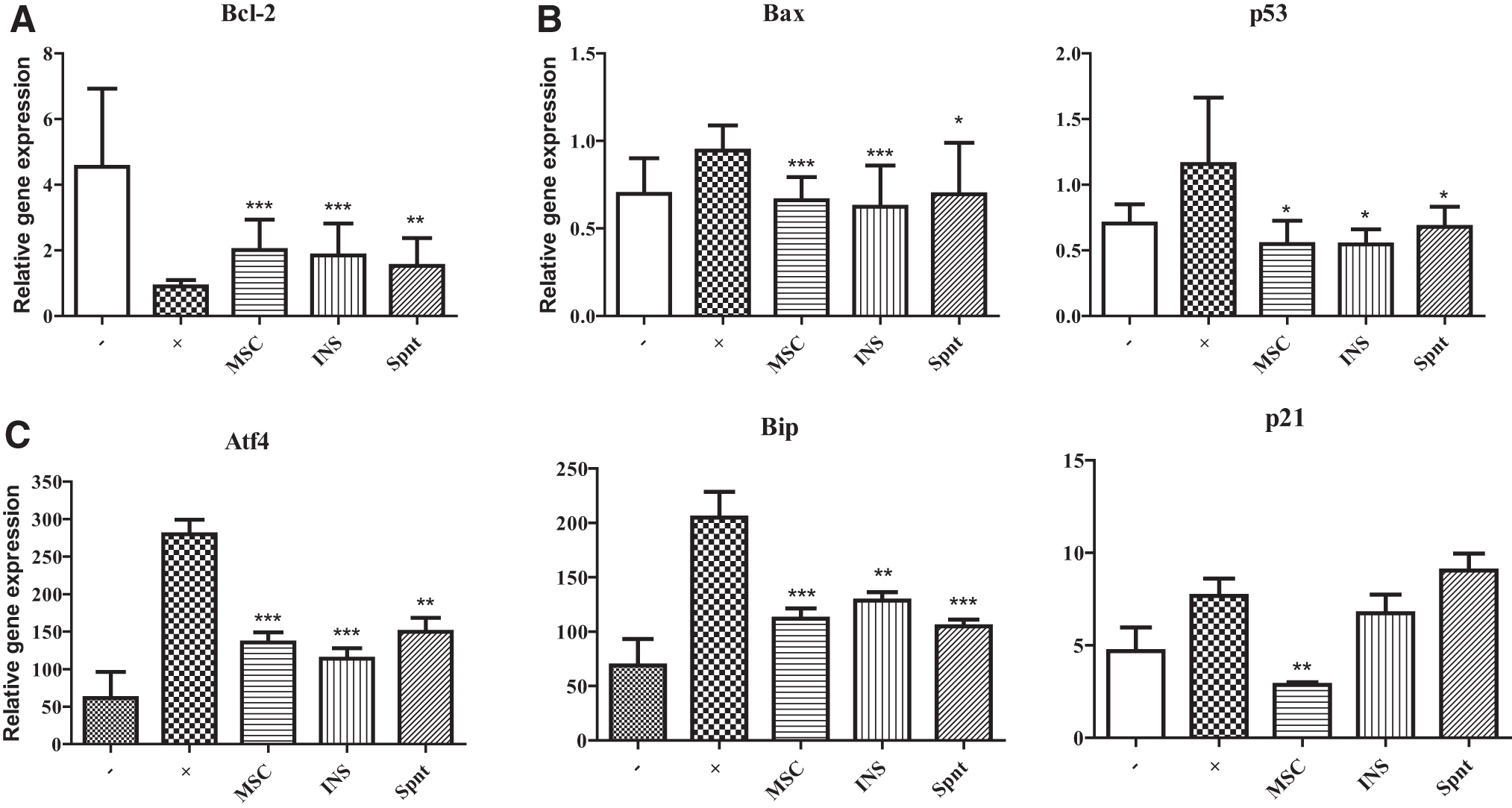
Antiapoptotic effect of MSCs on stimulated corneal explants in vitro. Corneal explants were cultured for 48 h unstimulated (−), stimulated by IL-1β, TNF-α, and IFN-γ (+) in the presence of MSCs directly (MSC) or in insert (INS), or were stimulated in the presence of supernatant from cultures of MSCs (Spnt).
The inhibition of apoptosis of corneal cells by MSCs
The corneal explants were cultured for 48 h in the presence of proinflammatory cytokines with or without MSCs (or with supernatants from MSCs) and then homogenized and digested by collagenase II to obtain single-cell suspensions. A flow cytometry analysis was used to detect phosphatidylserine on the surface of cells by annexin V, and to determine the percentage of apoptotic cells. As demonstrated in Fig. 7, the percentage of apoptotic cells was increased after cultivation of the explants with proinflammatory cytokines. The percentage of apoptotic cells was significantly decreased in the presence of MSCs. This effect of MSCs was observed, even when the MSCs were separated from the explant in inserts or if the explants were stimulated with cytokines in the presence of supernatants from MSCs (Fig. 7).
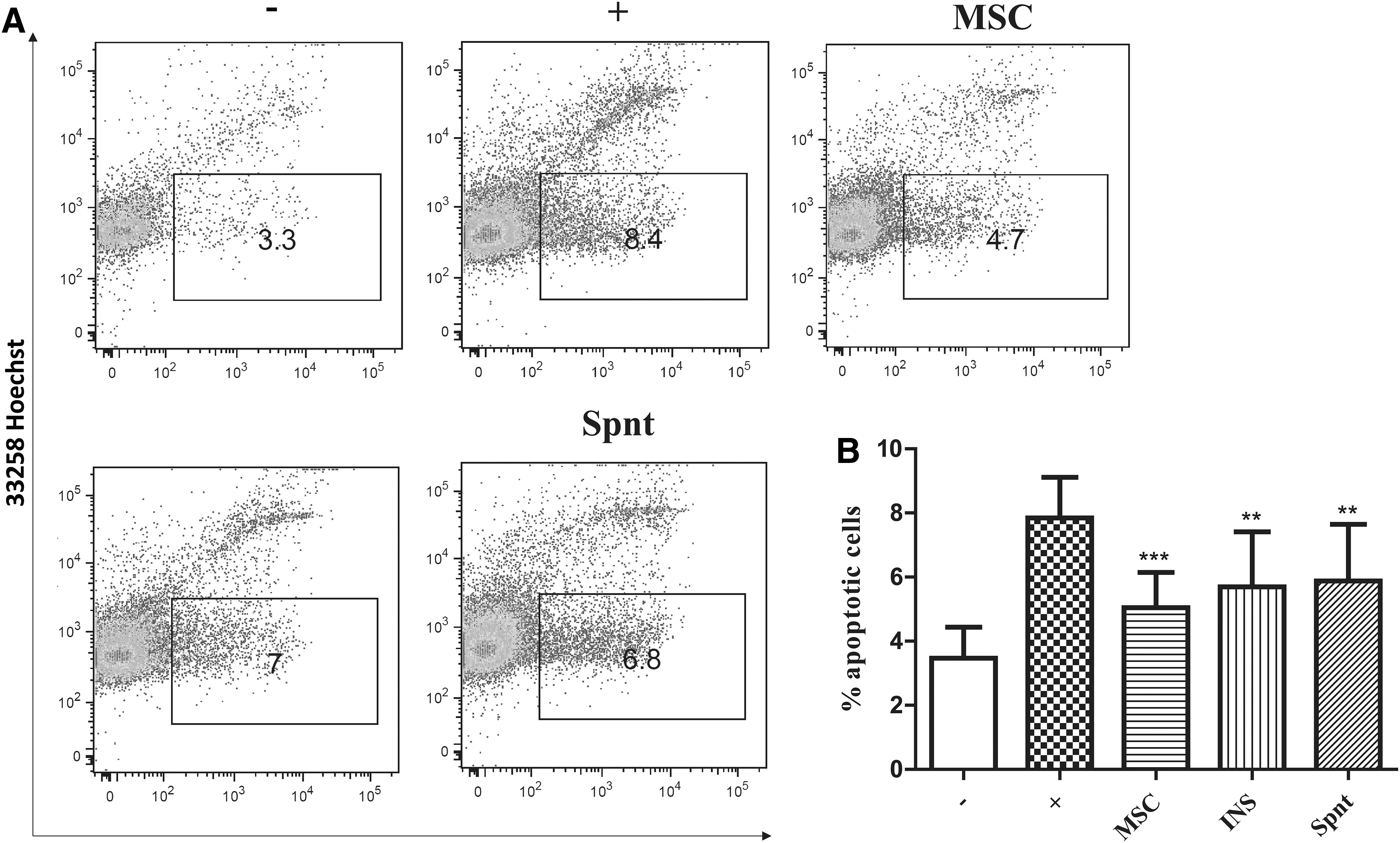
The inhibition of apoptosis in the cornea by MSCs. Excised corneas were cultured for 48 h unstimulated (−), stimulated with proinflammatory CYT IL-1β, TNF-α, and IFN-γ (+) in the presence of MSCs directly (MSC), or in insert (INS). Corneas were also cultured stimulated and treated by MSC supernatants (Spnt). Single-cell suspensions were prepared by enzymatic digestion from corneas and the percentage of apoptotic cells was measured using flow cytometry by detection of Annexin V.
Discussion
Corneal injuries or diseases are one of the main causes of a decreased quality of vision or even blindness. These disorders are associated with a local inflammatory reaction and with a loss of epithelial cells and keratocytes. Any damage of the cornea is accompanied by the production of stress proteins, secretion of proinflammatory cytokines and chemokines, and with infiltration with cells of the immune system. Increased levels of proinflammatory cytokines such as IL-1α, IL-1β, TNF-α, and IFN-γ were found in injured corneas in animal models and in patients [20 –22]. The proinflammatory cytokines can be produced by cells of the cornea and by infiltrating immune cells [1,2,23,24].
It has been shown in several in vitro and in vivo models that proinflammatory cytokines induce apoptosis of cells in the site of injury [3,4] and thus negatively influence the healing and regenerative processes. Therefore, a therapeutic strategy should involve the inhibition of an inflammation reaction and the prevention of apoptosis.
Using a model of corneal damage by alkali burn, we hereby demonstrated that the injury induces an enhanced expression of genes for proapoptotic molecules Bax and p53 and a decrease in the expression of gene for antiapoptotic molecule Bcl-2. These molecules have been used in multiple studies of apoptosis to determine the impacts of antiapoptotic therapy [25,26], and in the study of apoptosis in a model of dry eye in mice [27]. We also showed that the changes in the expression of genes for molecules associated with apoptosis were significantly inhibited by the treatment of injured corneas with MSCs. The ability of MSCs to inhibit a proinflammatory reaction and a harmful inflammation in the damaged cornea has been described [13,14]. Thus, MSCs turned out to be a perspective cell type, not only for the suppression of an inflammatory reaction but also for the attenuation of apoptosis.
To analyze the mechanism of the antiapoptotic effect of MSCs, we established a model of co-cultivation of corneal explants with MSCs in vitro. The apoptosis was induced in the explants by their cultivation for 48 h in the presence of proinflammatory cytokines IL-1β, TNF-α, and IFN-γ. As shown previously [3,4], these cytokines induce changes in the expression of genes associated with apoptosis (Bcl-2, Bax, and p53) and can induce apoptosis of corneal cells. Furthermore, we showed that the cultivation of corneal explants with proinflammatory cytokines induced the enhanced expression of genes for molecules Atf4, Bip, and p21, which are associated with ER stress. It has been shown that ER stress induced by proinflammatory cytokines enhanced the expression of the Bip gene in human corneas [5]. The level of Atf4 was also elevated in the study of keratoconus in stress conditions, and in the model of induced oxidative stress [28,29]. P21 plays a role in cell survival, but its expression is elevated in induced ER stress, while protecting cells against apoptosis and can be used as an ER stress marker [30,31]. We showed that the expression of genes for all these molecules is enhanced in corneal explants cultivated with proinflammatory cytokines, and that the increase is inhibited in the presence of MSCs.
To characterize the antiapoptotic effect of MSCs in more detail, corneal explants were cultured with proinflammatory cytokines and MSCs either directly or MSCs were separated in the inserts. Furthermore, MSCs were replaced by a supernatant obtained after the stimulation and cultivation of MSCs. The results showed that the antiapoptotic effect of MSCs was preserved if the direct contact of MSCs and explants was prevented, or if the MSCs were replaced by their supernatant. These observations suggest that antiapoptotic effects of MSCs are mediated by the paracrine action of MSCs.
To extend the study on the gene expression level, the corneal explants were cultured with proinflammatory cytokines in the absence or presence of MSCs, and the number of apoptotic corneal cells was determined by flow cytometry. We found that proinflammatory cytokines significantly increased the number of apoptotic cells and that MSCs inhibited this decrease. Again, the effect of MSCs was mediated by paracrine action.
The immunosuppressive properties of MSCs and their ability to modulate immune reactions have been well documented [6 –8]. To characterize molecules that could play a role in the antiapoptotic effects of MSCs in the cornea, we cultured MSCs unstimulated or stimulated with proinflammatory cytokines. We observed an elevated expression of genes for IDO, COX-2, PD-L1, and IL-6 after a 48-h stimulation. MSCs also produce several growth factors, such as HGF, TGF-β, basic FGF (bFGF), IGF-1, or EGF, which could be involved in their antiapoptotic and therapeutic effect. The expression of some of these factors (TGF-β, HGF, and bFGF) was decreased after stimulation with proinflammatory cytokines, while the expression of IGF-1 was increased. It has been shown that IDO, COX-2, and PD-L1 play a role in the inhibition of an immune response [32]. This effect might also be responsible for the inhibition of proapoptotic cytokines. IGF-1 plays a role in corneal regeneration and can inhibit apoptosis in corneal cells [33]. IL-6 is a multifunctional molecule, which is involved in both inflammatory and anti-inflammatory reactions, and is able to inhibit apoptosis through signal transducer and activator of transcription 3 activation [34]. IL-6 can also contribute to the inhibition of inflammation by increasing secretion of prostaglandin E2 [35] or by the induction of regulatory T cells in combination with TGF-β [16]. HGF facilitates corneal epithelial cell migration and proliferation, and protects corneal cells against apoptosis [36]. HGF also inhibits a myofibroblast phenotype in the cornea [37], and it has been shown that HGF enhances cell survival under oxidative stress in myocardial infarction [38]. Therefore, numerous factors produced by MSCs spontaneously or in an inflammatory environment can contribute to the antiapoptotic effects of these cells.
Conclusion
In conclusion, we showed that corneal injury or the incubation of corneal explants with proinflammatory cytokines induces the enhanced expression of proapoptotic genes and increases the number of apoptotic corneal cells. These effects can be inhibited by MSCs that attenuate the expression of proapoptotic genes in a paracrine way. We suggest that the inhibition of apoptosis is therefore one of the mechanisms of therapeutic action of MSCs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the project from the Grant Agency of the Charles University no. 1516218, grant no. 19-02290S from the Grant Agency of the Czech Republic, and the Charles University programs SVV 260435 and 20604315 PROGRES Q43.