
Abstract
In women of reproductive age, severe injuries to the ovary are often accompanied by premature ovarian failure (POF), which can result in amenorrhea or infertility. Hormone replacement therapy has been used to treat POF; however, it has limited therapeutic efficiency and may cause several side effects. In this study, we aimed to fabricate a Matrigel scaffold loaded with human umbilical cord-derived mesenchymal stem cells (MSCs) and explore its potential to restore ovarian function and repair ovarian structures in vitro and in vivo. POF mouse models were established by injecting mice with cyclophosphamide for 15 consecutive days. Then, MSC/Matrigel was transplanted into the ovaries of the mice. Five weeks later, the morphology of the ovaries and follicles was observed by hematoxylin/eosin staining, and the tissue fibrosis ratio was measured using Masson's trichrome staining. The number of blood vessels was evaluated by α-smooth muscle actin and CD31 immunofluorescence, and Ki67 expression was used to determine the proliferation of granulosa cells. The expression of vascular endothelial growth factor (VEGF)-A was assessed by western blotting. The Matrigel scaffold regulated the expression of VEGF-A in vitro. Moreover, it promoted MSC survival and proliferation and prevented MSC apoptosis in vivo. After the transplantation of the MSC/Matrigel, the number of follicles was significantly increased in the mice with POF, and the tissue fibrosis ratio was reduced. Furthermore, the MSC/Matrigel significantly improved the proliferation rate of granulosa cells, increased the number of blood vessels, and upregulated the expression of VEGF-A. These findings demonstrate that MSC/Matrigel may support follicular development and help restore ovarian structures in vivo.
Introduction
Premature ovarian failure (POF), also known as early menopause, is diagnosed by complete amenorrhea lasting for >4 months and a hypoestrogenic-hypergonadotropic serum profile (follicle-stimulating hormone levels ≥40 mIU/mL on two occasions) before 40 years of age [1,2]. The features of POF include hypoestrogenism, hypergonadotropism, and amenorrhea, which contribute to female infertility and premenopausal syndrome [3]. Women with POF have a decreased quality of life because of hypoestrogenism, night sweats, and hot flashes, and may also experience negative effects on other health aspects, including increased risk of cardiovascular diseases, sexual dysfunction, diabetes mellitus type 2 [3], osteoporosis [3], and even early mortality [4].
To date, hormone replacement therapy (HRT) is typically used to treat POF; however, this treatment usually leads to severe adverse effects, such as cardiovascular diseases, osteoporosis, urogenital atrophy, and neurodegenerative diseases [5]. Effective therapeutic strategies with specific targets are therefore needed to improve the treatment efficacy of POF.
Depending on the cellular source, mesenchymal stem cells (MSCs) can be classified as bone marrow-derived [6], umbilical cord-derived (MSCs) [7], adipose-derived human amnion-derived mesenchymal stem cells (hADMSCs), and human menstrual blood-derived [8]. MSCs are considered a promising source for cell-based therapies because of their ease of collection, low immunogenicity, and high proliferative potential [9]. MSC therapy has regenerative potential in neural, myocardial, skin, liver, kidney, cartilage, bone, muscle, and other tissue injuries [10,11]. The therapeutic potential of MSCs is mainly attributed to their paracrine functions, by which they secrete a variety of soluble factors to exert immunomodulatory, angiogenic, anti-apoptotic, and anti-oxidative effects [12 –14]. In addition, MSCs have the potential to replace damaged tissue by differentiating into various cell lineages [15].
A growing body of preclinical and clinical evidence in the recent literature has suggested that MSCs could be used to promote fertility preservation as part of oncofertility preservation strategies, for other types of primary ovarian insufficiencies (POIs), and to prevent natural ovarian aging. Several studies have suggested that the major mechanism underlying the effects of MSC-based therapy on POI is its paracrine function [16 –20].
Liu et al. [21] suggested that hADMSCs could upregulate the expression of vascular endothelial growth factor (VEGF) and insulin-like growth factor-1 and inhibit ovarian granulosa cell apoptosis. Moreover, Cui et al. [22] showed that human MSC transplantation improves ovarian function by decreasing the expression of transforming growth factor (TGF)β-1 and the phosphorylation of mothers against decapentaplegic homolog 3 (SMAD3) and by reducing tissue fibrosis. Yang et al. [23] suggested that transplantation of membranous vesicles from MSCs could help recover ovarian function by upregulating angiogenic cytokines, such as VEGF, fibroblast growth factor (FGF), and angiogenin.
The ongoing challenges associated with developing MSC therapies include low MSC engraftment and determining whether MSC engraftment is needed for sustained benefits in various models of disease. Three-dimensional (3D) scaffolds are typically porous, biocompatible, and biodegradable materials that serve to provide structural support for cells and new tissue being formed, acting as a temporary extracellular matrix (ECM) to induce the natural processes of tissue regeneration and development [24].
Matrigel, a natural biomaterial and the main component of the ECM, has been widely used for tissue-enhanced engineering scaffolds [25] because it can provide physical support and a suitable microenvironment for transplanted stem cells [26].
Therefore, in this study, we investigated the effects of MSC transplantation on cell apoptosis, angiogenesis, and tissue fibrosis in a POF mouse model using MSC/Matrigel to perform orthotopic transplantation into mouse ovaries. We then assessed the therapeutic effects of MSCs on POF.
Materials and Methods
Experimental animals
Eight-week-old female-specific pathogen-free grade C57BL/6 mice were acquired from the Suzhou Institute of Biomedical Engineering and Technology, Chinese Academy of Sciences (Suzhou, China) and were fed a standard diet with free access to water. Vaginal smears were obtained daily. Only mice showing at least two consecutive normal 4- to 5-day vaginal estrus cycles were included in the experiments. The animal experimental protocol was approved by the University Ethics Committee for the Use of Experimental Animals and conformed to the Guidelines for the Care and Use of Laboratory Animals.
Establishment of the POF mouse model
To establish a chemotherapy-induced POF mouse model, mice were intraperitoneally injected with 100 mg/kg cyclophosphamide (Sigma-Aldrich, St. Louis, MO) on the first day and then with 10 mg/kg/day cyclophosphamide for 14 consecutive days. A control group was injected with 0.9% saline instead of cyclophosphamide.
Isolation and culture of MSCs and green fluorescent protein-positive MSCs
The umbilical cord sample was provided by the clinic of the First Affiliated Hospital of Soochow University (Suzhou, China). Written informed consent was obtained, and the procedure was approved by the research ethics committee with patient consent. All studies were done in accordance with the Ethical Guiding Principles on Human Embryonic Stem Cell Research (of the Ministry of Science and Technology and the Ministry of Health, People's Republic of China, 2003) and Helsinki Declaration. MSC isolation was performed as previously described [27]. After the vessels were mechanically removed, the umbilical cord was manually dissected into small sections, which were cultured in Dulbecco's modified Eagle medium/Nutrient Mixture F-12 (DMEM/F12; Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (Hyclone, Logan, UT), 100 IU/mL penicillin (Gibco, Waltham, MA), and 100 μg/mL streptomycin (Gibco). The first colony of MSCs was observed after ∼10 days of culture, and those obtained at passages 3–5 were used for subsequent experiments.
Some cells were infected with a lentivirus carrying green fluorescent protein (GFP) to obtain GFP-labeled cells that could be traced. Direct in vitro observation and anti-GFP immunofluorescence experiments verified the presence of GFP in the cells.
Flow cytometric analysis
The specific cell surface antigens of cultured MSCs and green fluorescent protein-positive (GFP+) MSCs were analyzed by flow cytometry using a LSRFortessa cytometer (Becton Dickinson, Franklin Lakes, NJ). Cells were harvested with 0.05% trypsin/ethylenediaminetetraacetic acid (EDTA; Gibco) and incubated with phycoerythrin- or fluorescein isothiocyanate-conjugated antibodies against human CD34, CD45, CD90, and CD105 (Biolegend, San Diego, CA).
Differentiation potential of MSCs
MSCs and GFP+ MSCs at passage 2 were assessed for multipotency by adipogenic and osteogenic differentiation assays. In brief, adipogenesis was induced using an adipogenic induction medium (Stemcell Technologies, Vancouver, Canada) for 23 days and confirmed by Oil Red O staining as an indicator of intracellular lipid accumulation. Osteogenesis was induced by culturing MSCs in an osteogenic induction medium (Cyagen, Santa Clara, CA) for 28 days. Calcium deposition was determined by Alizarin red staining.
Cell Counting Kit 8 assays
MSCs and GFP+MSCs were seeded at a density of 1 × 104 cells/well in a 96-well plate. After 1, 2, 3, or 4 days of culture, optical density value was evaluated using a Cell Counting Kit 8 (CCK8) kit (Beyotime, Shanghai, China) according to the manufacturer's protocol.
Coculture of MSCs in Matrigel
Third-generation MSCs were selected, and upon reaching 80%–90% confluence, cells were digested with 0.25% trypsin-EDTA, centrifuged, collected, and counted. The cell density was adjusted to 106 cells/mL. Under an ice bath, 50 μL of the cell suspension was mixed with 50 μL Matrigel hESC-Qualified Matrix (Cat. No. 354277; Corning®, Waltham, MA), placed in a 48-well plate, further cultured in DMEM/F12/10% fetal bovine serum/1% penicillin/streptomycin, and incubated at 37°C in a CO2 incubator.
Reverse transcription and real-time polymerase chain reaction
Total RNA was isolated using TRIzol reagent (Thermo Fisher Scientific). The quality of the RNA was evaluated by spectrophotometry and denaturing agarose gel electrophoresis. In brief, complementary DNA (cDNA) was synthesized from 1 mg purified total RNA using a Prime-Script RT reagent kit (Takara, Kusatsu, Japan) according to the manufacturer's instructions. Real-time polymerase chain reaction (RT-PCR) was then performed using the cDNA with a TB Green Premix Ex Taq (Takara) and a 96-Well Realtime PCR Detection System (Thermo Fisher Scientific). The amplification reaction was performed using 40 cycles as follows: denaturation at 95°C for 5 s and annealing at 60°C for 30 s. All gene expression levels were normalized to the level of the internal standard control, Gapdh, and analyzed using the 2−ΔΔCt method. The specific primers used in these experiments are given in Supplementary Table S1.
Transplantation of MSC/Matrigel
In total, 33 mice were randomly divided into 5 treatment groups: Control (n = 7), POF (n = 6), Matrigel (n = 6), MSC (n = 4), and MSC/Matrigel group (n = 4). The remaining four mice were used for tracing of transplanted MSCs. Mice were anesthetized with isoflurane and then shaved in the decubitus position. The ovaries of all mice were exposed through a back incision. Aliquot suspensions (5 μL) were injected into the core of the ovaries in a retrograde manner using 10 μL microliter syringes (Gaoge, Shanghai, China).
Each mouse was injected with a total of 5 μL of each suspension into each ovary. For the POF group, 5 μL of 0.9% saline solution (Baxter International, Deerfield, IL) was injected per ovary. For the Matrigel group, 5 μL of Matrigel, which was dissolved at 4°C in advance, was injected per ovary. For the MSC group, 5 × 105 GFP+ MSCs (passages 3–5) in 5 μL of saline solution were injected. For the MSC/Matrigel group, 5 × 105 GFP+ MSCs (passage 3–5) in 2.5 μL saline solution were mixed with 2.5 μL of Matrigel, and the suspensions were then injected. After pressing the injection site with sterile cotton buds for 3–5 min, the ovaries were placed back into the abdomen. The muscle and skin were sutured separately in an interrupted manner.
Stem cell tracking in vivo
Labeled MSC/Matrigel and MSCs were injected into the right and left ovaries of mice, respectively. The bilateral ovaries were removed at 5, 14, 21, and 28 days after transplantation, immersed in optimum cutting temperature compound (Sakura, Osaka, Japan), and frozen at −80°C. The ovaries were sectioned along the plane of the maximum cross-section and then serially sectioned at a thickness of 20 μm using a cryostat (Thermo Fisher Scientific).
Sections were fixed in 4% formalin solution at room temperature for 30 min and washed three times. After blocking by incubation in 5% bovine serum albumin for 1 h at room temperature, tissue sections were labeled with antibodies against GFP (1:500; Cat. No. ab112; Abcam, Cambridge, United Kingdom), Ki67 (1:100; Ebioscience, San Diego, CA), cleaved caspase 3 (CC3; 1:400; Invitrogen, Waltham, MA), or α-SMA and incubated overnight at 4°C. After washing, the sections were incubated with Alexa488/555/647-conjugated secondary antibodies for 2 h. Subsequently, the sections were washed three times in phosphate-buffered saline (PBS). Counterstaining was performed by incubating with 2-(4-amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI) for 15 min in the dark. Analyses were performed with a confocal microscope (laser scanning confocal microscopy [LSCM]; Nikon A1) at 200 × magnification.
Five random areas of the section were selected for counting, and the number of GFP+ cells was counted and summed from three sections for each sample. Bound by shuttle-shaped cells in the outer membrane of the follicle, the inside was considered to be the follicle, and the outside was considered to be the stromal region. The areas within two layers of cells outside the blood vessels (∼20 μm) were considered regions near blood vessels. Cells showing positive staining for GFP and Ki67 were considered Ki67+ MSCs. Cells showing positive staining for GFP and CC3 were considered CC3+ MSCs.
E2 level assays
After 5 weeks, the mice were killed, and blood was collected by cardiac puncture. Serum and blood cells were separated by centrifugation, and serum E2 levels were determined using a commercially available enzyme-linked immunosorbent assay (Nanjing Jiancheng Bioengineering, Nanjing, China).
Estrous cycle analysis
The estrous cycle was routinely assessed by vaginal smear, and the stage of the estrous cycle was determined as previously described [28].
Ovarian morphology and follicle counting
After 5 weeks, the mice were killed, and the ovaries were collected during diestrus. The ovaries were dissected, weighed, fixed overnight in 4% formalin solution, dehydrated in an ascending series of ethanol, and embedded in paraffin. Then, 8 μm thick sections were prepared, and every fifth section was used as a histologic slice and stained with hematoxylin and eosin. All sections were examined under a light microscope with a magnification of 200 × . The follicles were classified as previously described [29]. Only follicles with oocyte nuclei were counted. After counting every fifth section, the number of follicles was multiplied by five.
Masson's trichrome staining
All ovary sections were stained with Masson's trichrome technique, as previously described [30], in which collagen fibers were stained blue; cytoplasm, muscle fibers, and red blood cells were stained red; and nuclei were stained black [30]. ImageJ software (v1.51; National Institutes of Health, Bethesda, MD;
Immunohistochemistry
Paraffin-embedded sections of the ovary tissues were deparaffinized and rehydrated, and endogenous enzymes were blocked by incubation in 5% bovine serum albumin for 1 h at room temperature. The sections were then immunolabeled with antibodies against GFP (1:500; Cat. No. ab112; Abcam), Ki67 (1:100; Ebioscience), α-SMA (1:200; Cat. No. ab28364; Abcam), CD31 (1:500; SAB Biotherapeutics, Sioux Falls, SC), or CC3 (1:400; Invitrogen), incubated overnight at 4°C, washed three times in PBS, and incubated with Alexa488/555/647-conjugated secondary antibodies for 2 h. Subsequently, the sections were washed three times in PBS. Counterstaining was performed by incubating with DAPI for 15 min in the dark. Analyses were performed with a confocal microscope (Nikon) at 200 × magnification. Image-Pro Plus 6.2 software (Media Cybernetics, Bethesda, MD) was used to measure the mean optical density of immunostaining in follicles under a fluorescence microscope.
For α-SMA, five random areas of the section were selected for counting under 200 × magnification. Any red-stained vascular smooth muscle cells or cell clusters that were clearly separated from adjacent cells and other connective tissue elements were considered a single countable vessel. For CD31, five random areas of the section were selected for counting under 200 × magnification. For Ki67, five random areas of each section were selected for counting, and all secondary and preantral follicles in these areas were counted for the percentage of Ki67+ cells/DAPI+ cells. An oocyte wrapped around multiple layers of granulated cells was recognized as a secondary follicle. When granular cells began to gather on one side and cavities appeared in the follicles, they were considered to be preantral follicles.
Western blotting
For western blot analysis, protein lysates from fresh ovarian tissues were prepared, separated on 12% sodium dodecyl sulfate–polyacrylamide gels, and transferred to polyvinylidene difluoride membranes (Millipore, Burlington, MA). Membranes were blocked with 5% nonfat milk and individually incubated with the following specific antibodies at 4°C overnight: anti-VEGF-A (Abcam) and anti-β-actin (Abcam). After washing, the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (1:10,000; Abcam). Visualization of the blots was performed using a standard protocol for enhanced chemiluminescence (Thermo Fisher Scientific). The relative intensity of protein bands was quantified by digital densitometry using ImageJ software. Actin levels were used as a loading control.
Statistical analysis
The data were analyzed using Prism (GraphPad Software, San Diego, CA) and presented as means ± standard errors of the means. One-way analysis of variance was used, followed by post hoc Tukey's multiple comparison tests to analyze the data. Results with values of P < 0.05 were considered statistically significant.
Results
Characterization of MSCs
To confirm whether the cells isolated from human umbilical cords possessed the properties of MSCs, cell differentiation and flow cytometry analyses were performed. The differentiation potency of these cells was confirmed by observing osteogenesis and adipogenesis (Fig. 1B, C). Moreover, as given in Fig. 1D–G, the cultured MSCs expressed high levels of the MSC markers CD90 (97.30%) and CD105 (96.9%) and low levels of the hematopoietic markers CD34 (3.77%) and CD45 (1.79%).
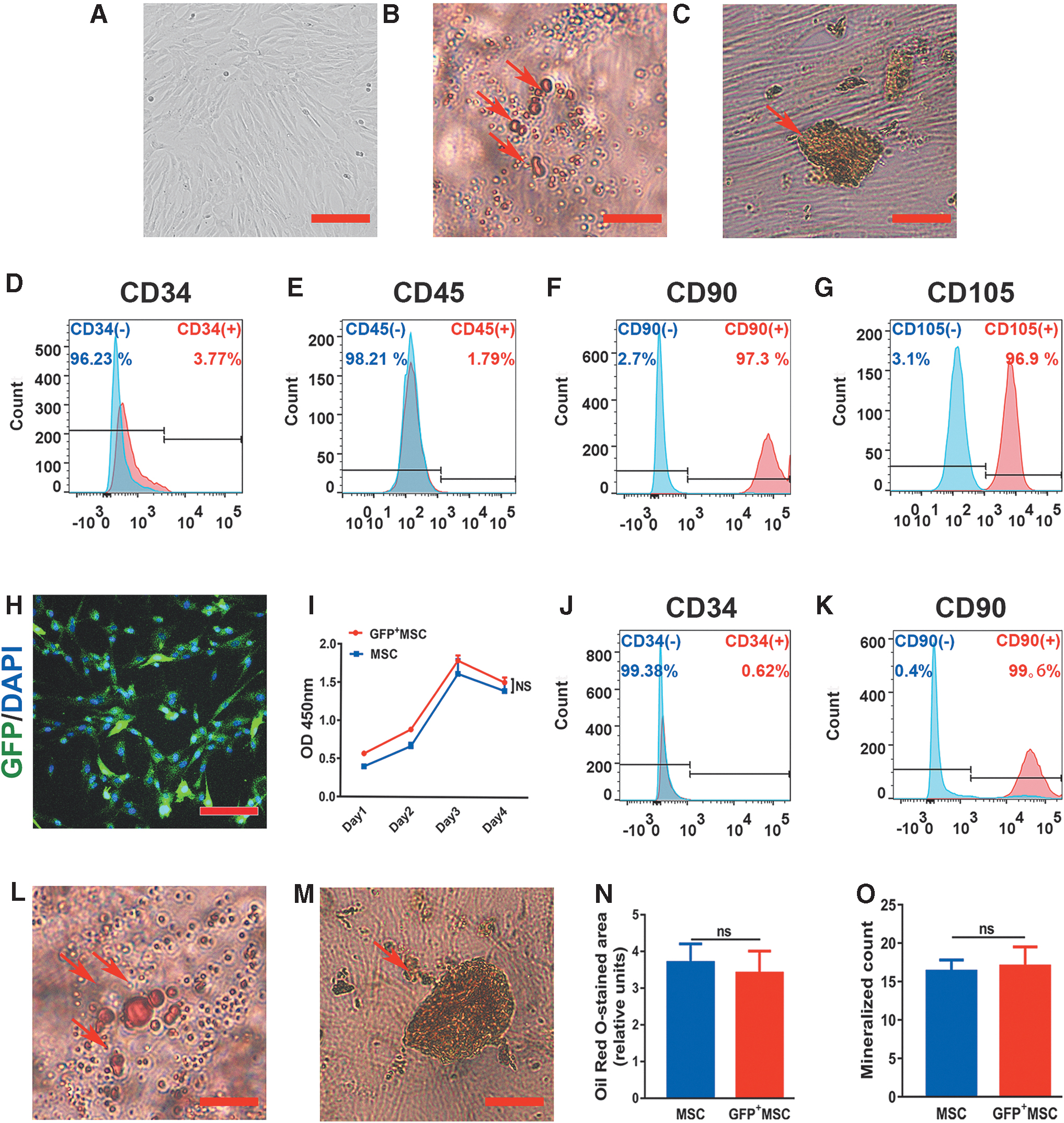
Characterization of MSCs and GFP+ MSCs.
To test whether GFP+ MSCs were altered compared with MSCs, we next examined their differentiation potency, cell proliferation, and MSC marker expression. CCK8 assays did not show significant differences between GFP+ MSC and MSC groups regarding cell proliferation capacity (Fig. 1I). Further evaluation by flow cytometric analysis showed that GFP+ MSCs expressed high levels of the MSC marker CD90 (99.6%) and low levels of the hematopoietic marker CD34 (0.62%), similar to untransfected MSCs (Fig. 1J, K). In addition, GFP+ MSCs could differentiate into osteogenic and adipogenic cells (Fig. 1L, M). Compared with the MSC group, there were no significant differences in the GFP+ MSC group in terms of mineralized count and Oil Red O-stained area (relative units; Fig. 1N, O). This demonstrated that GFP expression in MSCs had no impact on their stem cell properties.
Cell seeding and properties of MSCs in Matrigel scaffolds in vitro
To test whether MSC properties were modified by the Matrigel scaffold, RT-PCR and flow cytometry analyses were performed on MSC/Matrigel. After coculture with Matrigel for 24 h, MSCs resembled fibroblasts morphologically, suggesting that the structure of Matrigel could play an important role in cell adhesion and growth (Fig. 2A). Moreover, after coculture with Matrigel for 5 days, significantly lower expression of TGFβ-1 and higher expression of epidermal growth factor (EGF) and TGFβ-3 were observed in MSC/Matrigel (Fig. 2B). In addition, >99% of MSCs expressed CD90 and CD105, and few were positive for CD34 and CD45 (Fig. 2C–F), consistent with our previous data. These results indicated that MSC properties were not altered in Matrigel, although the secretion of growth factors in MSCs was regulated.

Cell seeding and properties of MSCs in Matrigel scaffolds in vitro.
Matrigel scaffold promoted MSC survival and proliferation and reduced MSC apoptosis
To evaluate the distribution of GFP+ MSCs in ovaries, the ovaries were collected and sectioned at 5, 14, 21, and 28 days after cell transplantation, and anti-GFP immunofluorescence was performed (Fig. 3A–C and Supplementary Fig. S1). The results showed that the GFP+ MSCs in the MSC group were settled in the injection area (Fig. 3B), whereas those in the MSC/Matrigel group were distributed throughout the ovary (Fig. 3C). These results indicated that Matrigel promoted the survival of MSCs and may prevent insufficient settlement of MSCs in the ovaries. In addition, 5 days after injection, whether in follicles, near blood vessels, or in the stromal region, the number of GFP+ cells in the MSC/Matrigel group was higher than that in the MSC group (P < 0.05, Fig. 3D).
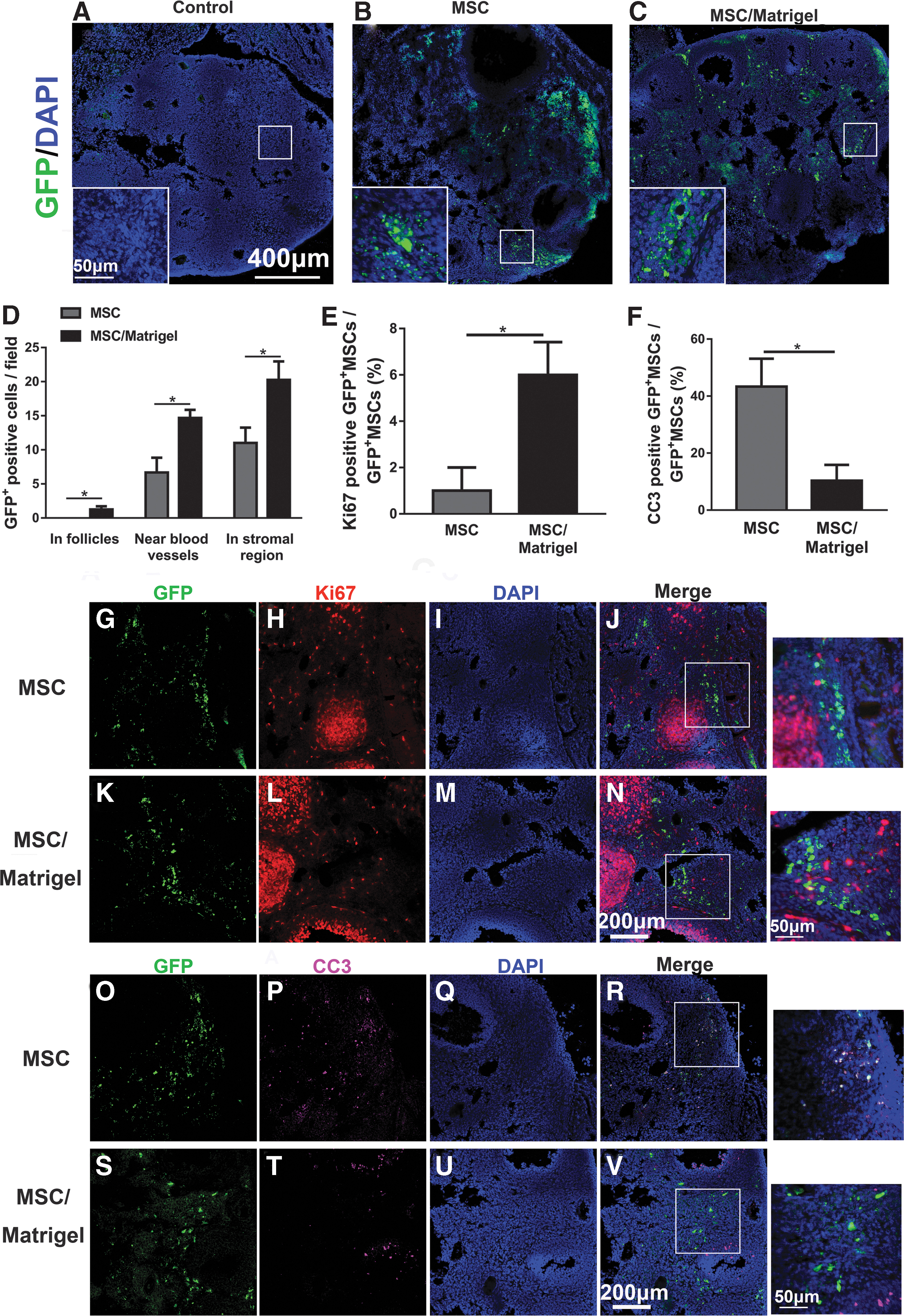
Matrigel scaffold promoted MSCs survival and proliferation, and reduced MSCs apoptosis 5 days after MSC transplantation.
In addition, 14, 21, and 28 days after injection, the number of GFP+ cells in the MSC/Matrigel group was also found to be greater than that in the MSC group at each time point, which indicated that Matrigel may promote MSC survival (Supplementary Fig. S1).
To determine whether the MSCs were proliferating or undergoing apoptosis in the MSC and MSC/Matrigel groups, colocalization of the GFP signal with Ki67 and CC3 markers was assessed (Fig. 3G–V). Compared with the MSC group, the percentage of Ki67+ MSCs was significantly higher in the MSC/Matrigel group (P < 0.05, Fig. 3E), and the percentage of CC3+ MSCs was significantly lower in the MSC/Matrigel group (P < 0.05, Fig. 3F). These results demonstrated that the Matrigel scaffold promoted MSC proliferation and prevented MSC apoptosis in vivo.
MSC/Matrigel increased the ovarian index and the number of follicles in POF model mice
Compared with those in the control group, the body weights of mice in the POF model group were significantly lower (P < 0.0001; Supplementary Fig. S2), as was the ovarian index (% ovarian weight/body weight; P < 0.0001; Fig. 4B). The number of total follicles in the POF group was decreased compared with that in the control group (P < 0.05, Fig. 4H). Moreover, the collagen volume fraction in the POF group was significantly increased compared with that in the control group (P < 0.0001; Fig. 5F). The number of blood vessels per field (P < 0.05; Fig. 6B) and the relative expression levels of VEGF-A (P < 0.0001; Fig. 6E) and Ki67 (P < 0.0001; Fig. 7P) in the POF group were decreased significantly compared with those in the control group. These results indicated that ovarian function was severely impeded in the POF mouse model.
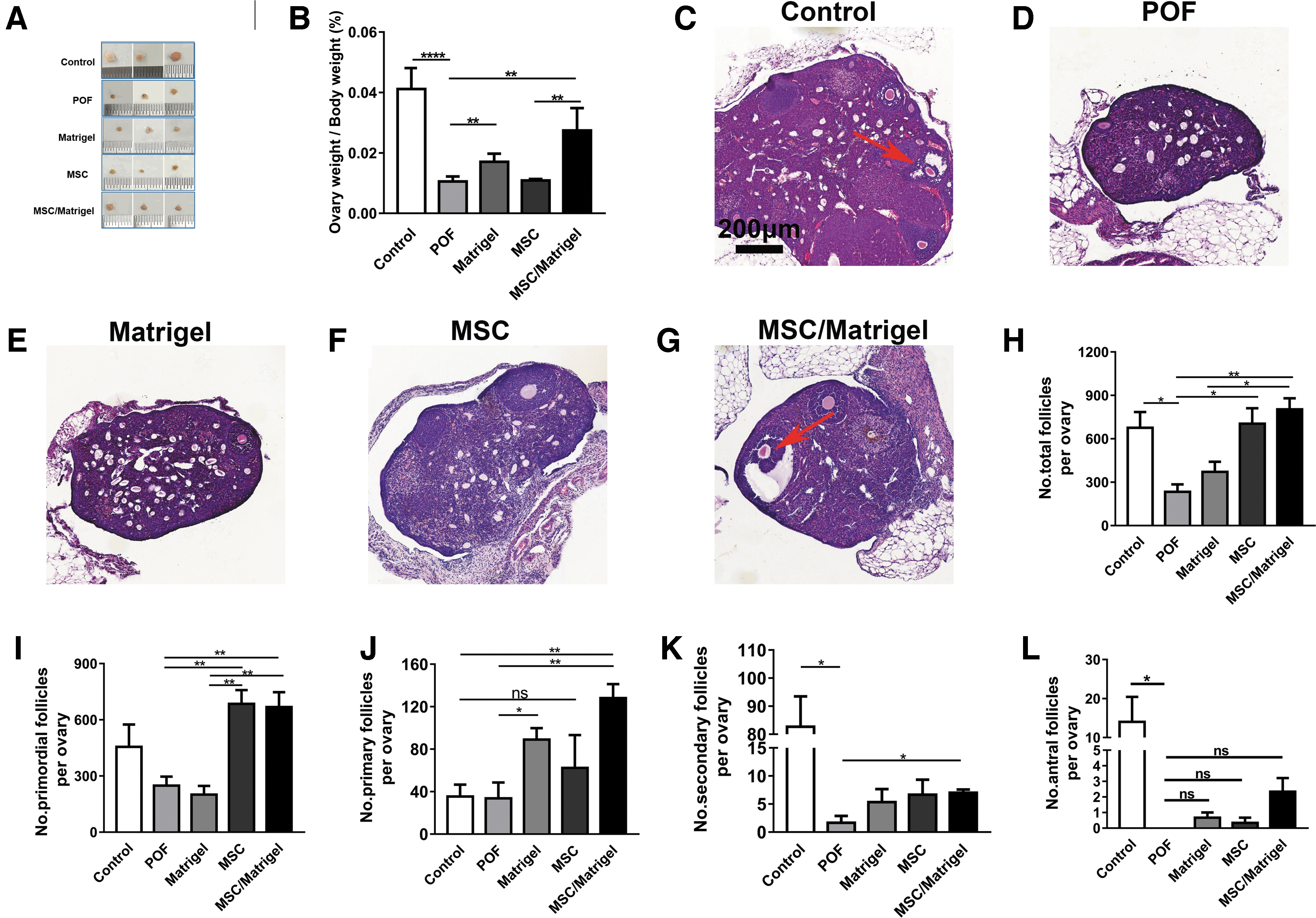
MSC/Matrigel improved ovarian index and the number of follicles in POF mouse model 5 weeks after MSC transplantation.
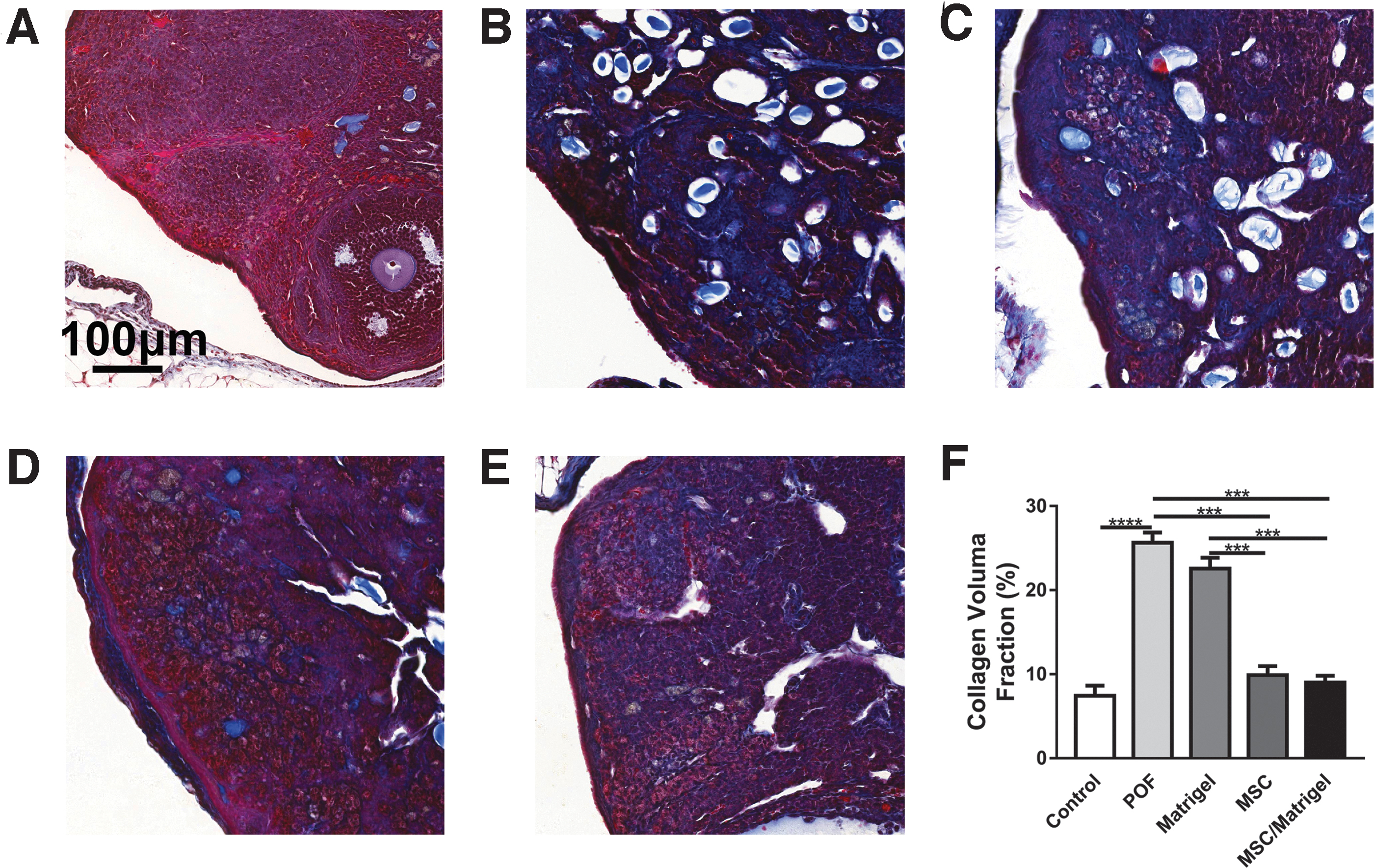
MSC/Matrigel decreased the tissue fibrosis ratio in POF mouse model 5 weeks after MSC transplantation.
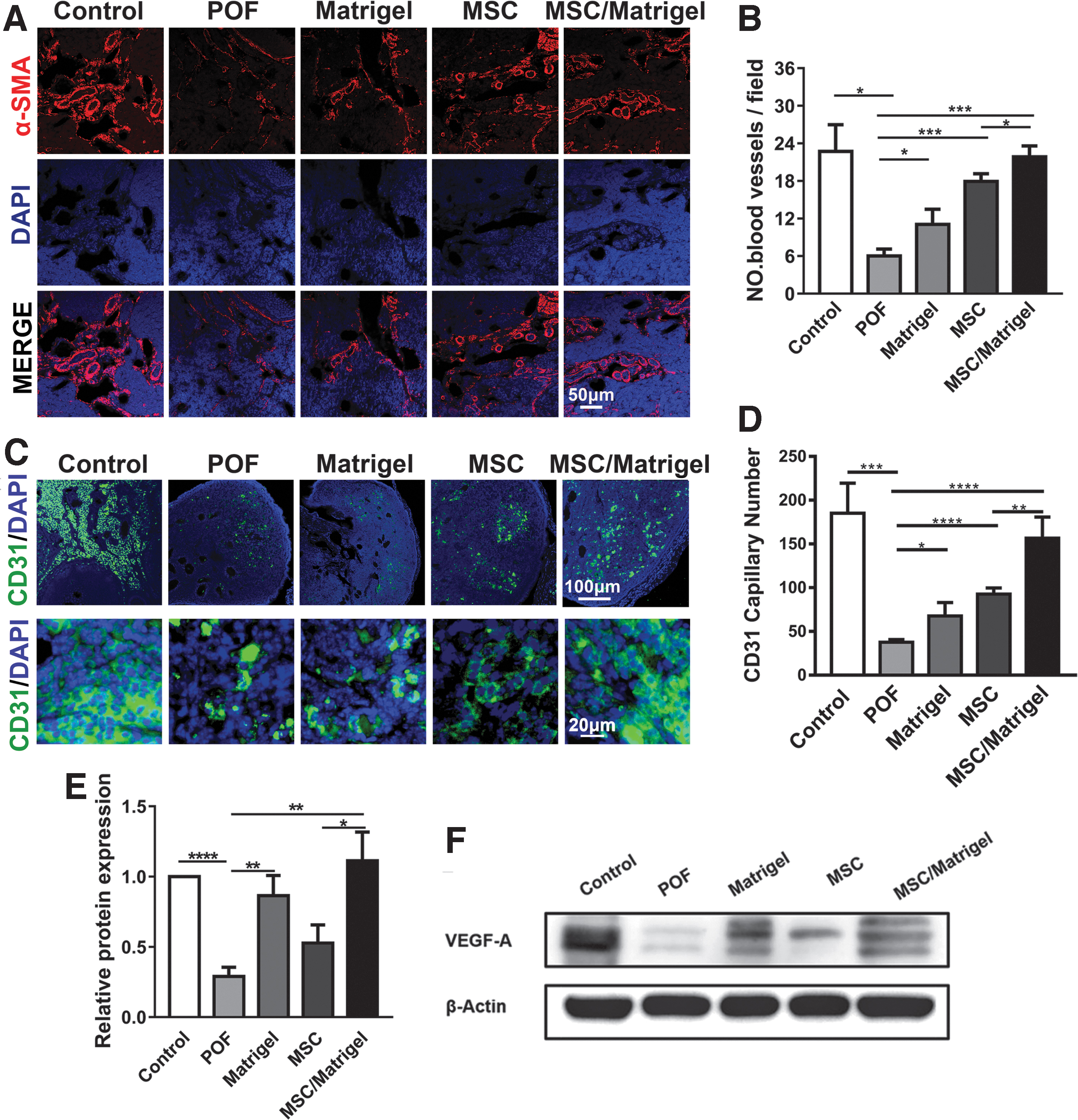
MSC/Matrigel transplantation increased the number of blood vessels and the expression of VEGF-A in the POF mouse model 5 weeks after MSC transplantation.
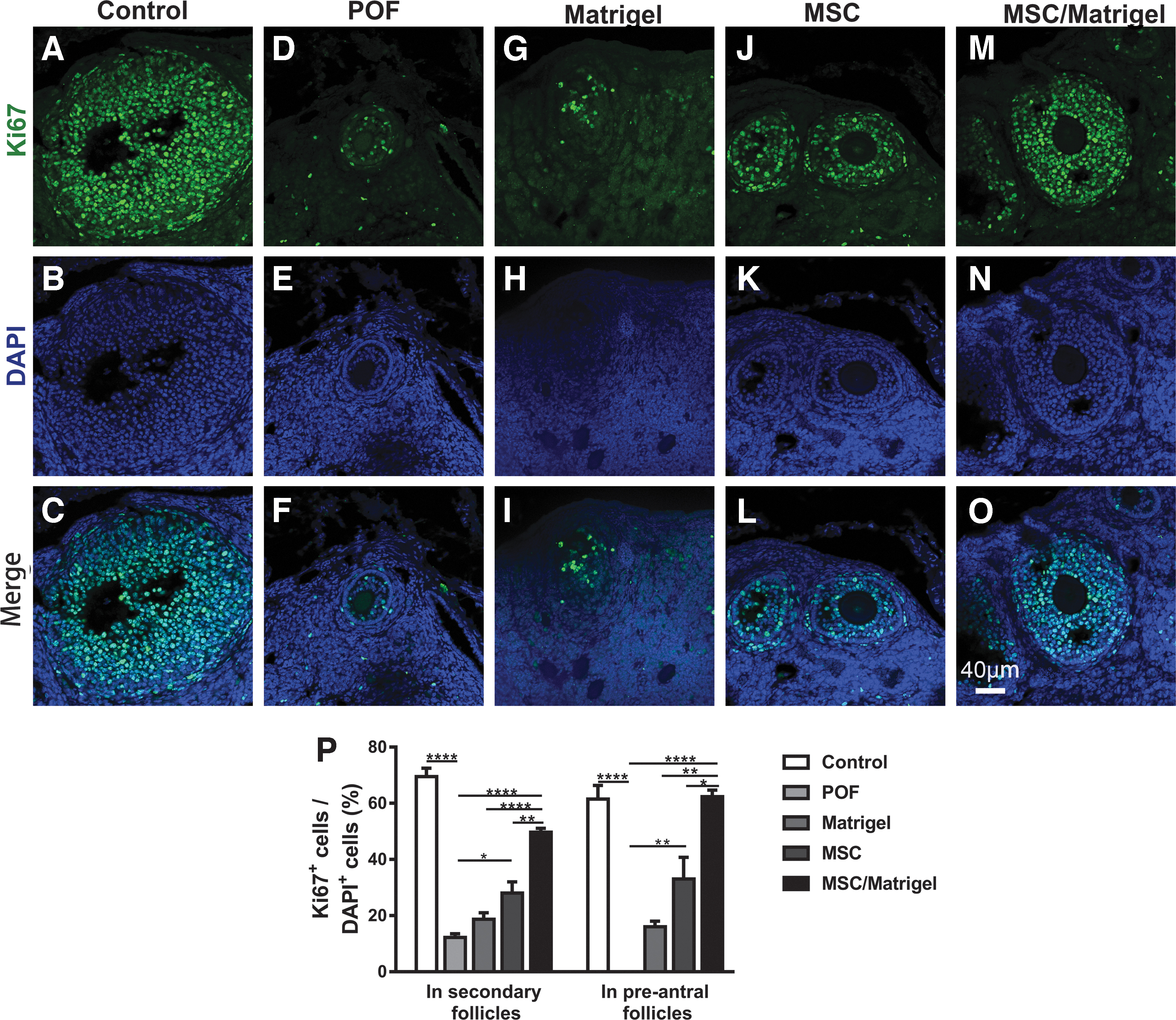
MSC/Matrigel transplantation improved the proliferation rate of granulosa cells in the POF mouse model 5 weeks after MSC transplantation.
To better understand the potential therapeutic effects of MSC/Matrigel, we compared the body weights, ovarian indices, and follicle numbers in mice at 5 weeks after cell transplantation. The results indicated that body weights in the Matrigel, MSC, and MSC/Matrigel groups were not significantly different compared with those in the POF group (Supplementary Fig. S2). By contrast, the ovarian indices of mice in the Matrigel (P < 0.01) and MSC/Matrigel groups (P < 0.01, Fig. 4B) were significantly higher than those of mice in the POF group. In addition, the ovarian indices of mice in the MSC/Matrigel group were significantly higher than those of mice in the MSC group (P < 0.01, Fig. 4B). These results suggested that MSCs and Matrigel transplantation could prevent ovarian weight loss.
To evaluate the possible mechanisms through which MSCs increase ovarian weight, serum E2 levels and days of estrous cycle were determined (Supplementary Fig. S3), and the ovaries were collected for histological analysis and assessment of the number of follicles at different stages of development (Fig. 4C–H). The E2 level in the MSC/Matrigel group was significantly higher than that in the POF group (P < 0.05; Supplementary Fig. S3A). In addition, the days of estrous cycle in the MSC/Matrigel group were significantly lower than those in the POF group (P < 0.05; Supplementary Fig. S3B), and in the MSC group, the numbers of total follicles (P < 0.05; Fig. 4H) and primordial follicles (P < 0.01, Fig. 4I) were significantly higher than those in the POF group.
Of note, in the MSC/Matrigel group, the numbers of total follicles (P < 0.01; Fig. 4H), primordial follicles (P < 0.01; Fig. 4I), primary follicles (P < 0.01; Fig. 4J), and secondary follicles (P < 0.05; Fig. 4K) were significantly higher than those in the POF group. Moreover, the POF group did not have antral follicles, whereas MSC/Matrigel transplantation increased the formation of antral follicles by up to 20% compared with that in the control group (Fig. 4L). In the Matrigel group, only the number of primary follicles was higher than that in the control group (P < 0.05; Fig. 4J). These results suggest that Matrigel or MSCs alone increased the numbers of different types of follicles, although there were no synergistic effects of combining Matrigel and MSCs.
MSC/Matrigel decreased the tissue fibrosis ratio and promoted angiogenesis in POF model mice
To investigate the underlying mechanisms through which MSCs promote follicular development, the tissue fibrosis ratio, number of blood vessels, and expression of the angiogenic factor VEGF-A were analyzed. Masson's trichrome staining was performed on ovarian sections to determine the collagen volume fraction of the ovaries (Fig. 5A–E), revealing marked suppression in the MSC/Matrigel (P < 0.001) and MSC groups (P < 0.001) compared with that in the POF group (Fig. 5F). In addition, it was also remarkably suppressed in the MSC/Matrigel (P < 0.001) and MSC groups (P < 0.001) compared with that in the Matrigel group (Fig. 5F). However, the collagen volume fraction in the Matrigel group remained high. These results suggested that MSC transplantation decreased tissue fibrosis.
To test the number of blood vessels per field, we performed α-SMA and CD31 immunofluorescence staining in each group of mice (Fig. 6A, C). A significantly increased number of blood vessels/field was observed in the ovary sections of the Matrigel (P < 0.05), MSC (P < 0.001), and MSC/Matrigel groups (P < 0.001) compared with the POF group (Fig. 6B), and the number of blood vessels per field in the MSC/Matrigel group was higher than that in the MSC group (P < 0.05; Fig. 6B).
In addition, compared with the POF group, the CD31+ capillary number was significantly higher in the Matrigel (P < 0.05), MSC (P < 0.0001), and MSC/Matrigel groups (P < 0.0001; Fig. 6D). In particular, the MSC/Matrigel group had more CD31+ capillaries than those in the MSC group (P < 0.01; Fig. 6D), suggesting that Matrigel and MSC transplantation may support angiogenesis.
In addition, to make sure GFP did not cause false positive while detecting Alexa 488-labeled cells in immunofluorescence assay, we performed negative controls in ovarian sections that replaced the primary antibody against CD31 with PBS (Supplementary Fig. S4). Results indicated that no spontaneous immunofluorescence of GFP was visible.
Moreover, the content of VEGF-A was significantly upregulated in the Matrigel (P < 0.01) and MSC/Matrigel (P < 0.01) groups compared with that in the POF group (Fig. 6F), further suggesting that Matrigel and MSC transplantation may increase the number of blood vessels and the expression of VEGF-A in POF mice.
MSC/Matrigel promoted granulosa cell proliferation in POF model mice
To further investigate the MSC-mediated mechanisms underlying follicular development, the proliferation rate of granulosa cells was analyzed. Immunohistochemical staining for Ki67 was performed in secondary and preantral follicles (Fig. 7A–O). In the MSC/Matrigel group, the percentage of Ki67+ cells in secondary follicles was significantly greater than those in the POF (P < 0.0001), Matrigel (P < 0.0001), and MSC groups (P < 0.01; Fig. 7P). Moreover, the percentage of Ki67+ cells in secondary follicles was also higher in the MSC group than that in the POF group (P < 0.05; Fig. 7P).
In addition, the percentage of Ki67+ cells in preantral follicles was significantly greater in the MSC/Matrigel group than those in the POF (P < 0.0001), Matrigel (P < 0.01), and MSC groups (P < 0.05, Fig. 7P), and the percentage of Ki67+ cells in preantral follicles was higher in the MSC group than that in the POF group (P < 0.01; Fig. 7P). These findings indicate that MSC/Matrigel promoted granulosa cell proliferation in secondary and preantral follicles. In addition, to further understand the significant advantage of using MSC/Matrigel together, the summary table (Table 1) provides the statistical differences of the above indicators between the MSC group and the MSC/Matrigel group.
The Summary Table of the Statistical Differences Between the Mesenchymal Stem Cell Group and the Mesenchymal Stem Cell/Matrigel Group
CC3, cleaved caspase 3; DAPI, 2-(4-amidinophenyl)-6-indolecarbamidine dihydrochloride; GFP+, green fluorescent protein positive; MSC, mesenchymal stem cell; POF, premature ovarian failure; VEGF, vascular endothelial growth factor.
Discussion
Traditionally, most patients with POF are treated with HRT. However, long-term HRT may increase the risk of cardiovascular disease, osteoporosis, urogenital atrophy, and neurodegenerative diseases [31,32]. In recent years, the therapeutic effects of stem cells on POF have been reported [33 –35]. In addition, 3D scaffolds combined with stem cells can exhibit improved therapeutic efficacy owing to enhanced cell survival and retention [36,37]. In this study, we fabricated Matrigel scaffolds loaded with MSCs, which demonstrated potential to decrease the tissue fibrosis ratio and promote the proliferation of granulosa cells and ovarian vasculature in mouse models of POF.
Ovarian fibrosis is primarily triggered by different factors, such as surgery, chemotherapy, radiotherapy, inflammation, and immune abnormalities [38,39]. Fibrosis is characterized by excessive proliferation of fibrous connective tissue and deposition of ECM [40]. Diffuse ovarian fibrosis can lead to decline of ovarian function [38,39]. A previous study demonstrated that MSC transplantation contributed to the inhibition of ovarian tissue fibrosis and restoration of ovarian function in POI rats through the TGF-β/SMAD3 signaling pathway [22]. In this study, the results showed that the tissue fibrosis ratio in the MSC and MSC/Matrigel groups was significantly decreased compared with those in the POF and Matrigel groups, indicating that MSCs may promote the repair of ovarian tissue fibrosis and restore ovarian function.
Angiogenesis is essential for ovarian development and function [41,42]. Quantification of endothelial cell proliferation shows that angiogenesis commences in the preantral follicle, increases with follicular development, and becomes intense in the early corpus luteum [43]. VEGF-A, which promotes the proliferation and differentiation of endothelial cells and remodels the vascular interstitial matrix, is important to angiogenesis [44]. In addition, VEGF-A plays an important role in the maintenance of capillary structures during final follicle maturation as well as in corpora lutea development and function [41]. Previous studies have indicated that ovarian angiogenesis can be suppressed by inhibition of VEGF [43].
Matrigel is a basement membrane protein mixture that contains ECM components and growth factors, such as basic FGF and EGF, and VEGF in Matrigel (Corning) is generally present at a concentration of 5–7 ng/mL. Previous studies have shown that Matrigel can promote angiogenesis [45]. Ou et al. [46] suggested that Matrigel promotes angiogenesis in the myocardium after ischemic injury. Herein, the results showed that the number of blood vessels and the expression of VEGF-A in the Matrigel and MSC/Matrigel groups were higher than those in the POF group, demonstrating that Matrigel and MSCs may promote angiogenesis by increasing VEGF-A expression.
However, the VEGF-A levels in the MSC group were not significantly different compared with those in the POF group. The VEGF content in Matrigel (Corning; 5–7 ng/mL) may mask the expression of VEGF-A in MSCs. Although the VEGF-A levels in the MSC group did not increase significantly compared with those in the POF group, MSCs increased the number of blood vessels in the POF group. This difference may be related to the involvement of various growth factors in angiogenesis. VEGF-A only promotes the growth of blood vessels; 50% of these growing blood vessels may wither and fail to grow into mature blood vessels. Thus, MSCs may secrete various cytokines to promote vascular stability.
Granulosa cells are critical to the development of follicles [47,48], with apoptosis of granulosa cells being the first event in follicular atresia [49]. Cyclophosphamide mainly acts on granular cells in the division and proliferation stages, thereby inducing their apoptosis [50,51].
According to previous reports, MSCs can secrete VEGF-A [52], which inhibits granulosa cell apoptosis by suppressing the downstream activities of caspase-3 independent of the mitochondrial pathway. Moreover, inhibition of VEGF-A activity appears to produce an increase in ovarian apoptosis, leading a larger number of follicles to atresia [53]. In stem cell transplantation, Matrigel can provide 3D support to transplanted cells [54] and promotes stem cell survival in vivo [55]. In our study, the results showed the expression of VEGF-A in the MSC/Matrigel group was significantly higher than that in the POF group, which indicates that MSC/Matrigel may promote the proliferation of granular cells by enhancing the upregulation of VEGF-A.
In our study, MSC/Matrigel transplantation recovered the reserve of primordial and growing follicles, but antral follicle numbers failed to reach those of the control group, consistent with a previous report [17,51,56,57]. Yang et al. [57] indicated that transplantation of 2 × 105 umbilical cord-derived mesenchymal stem cells (UC-MSCs) increased the numbers of primary and secondary follicles in POF mice. Moreover, Song et al. [51] transplanted 1 × 105 UC-MSCs (100 μL, at a concentration of 1 × 106 cells/mL) into a POF rat model and found that UC-MSC transplantation increased the number of secondary follicles and alleviated disruptions in hormone secretion in POF model rats. Although MSCs were involved in the observed responses in these experiments, the numbers of antral follicles in the MSC-treated group did not show a significant increase compared with those in the POF group.
In our study, we transplanted 5 × 105 MSCs combined with Matrigel, and the results that showed in the MSC/Matrigel group, the numbers of total follicles, primordial follicles, primary follicles, and secondary follicles were significantly higher than those in the POF group. Of note, the POF group did not have antral follicles, whereas MSC/Matrigel transplantation increased the formation of antral follicles by up to 20% compared with that in the control group. These results indicated that >5 × 105 MSCs may be needed to achieve complete repair. Further studies are needed to test the threshold amount that is sufficient to achieve beneficial effects. In addition, more studies must be performed to enhance stem cell function, such as the addition of growth factors and heat shock pretreatment of MSCs.
In addition, the numbers of primordial and primary follicles were increased in the MSC group, although no significant differences were observed between the POF and MSC groups. The reason for this occurrence may be related to the major differences between individual samples. Because the ovaries of mice are small, ovarian hematomas can occur during cell transplantation; this can lead to impaired ovarian structure and function. By contrast, MSC transplantation into the ovary in humans usually proceeds through a vaginal puncture guided by a vaginal mirror, which is less likely to cause hematoma in the ovaries.
A previous study reported that a biomimetic matrix 3D scaffold could induce MSCs to secrete factors [58]. Our study showed that the Matrigel scaffold promoted the expression of EGF in MSCs. EGF is a growth factor that stimulates cell migration and proliferation [59]; thus, EGF treatment promotes the expansion and paracrine activity of MSCs in vitro and MSC proliferation in vivo [60].
In addition, TGFβ-1 and TGFβ-3 have been implicated in the pathogenesis of fibrosis [40]. Cui et al. [22] reported that MSCs decreased ovarian fibrosis by reducing the expression of TGFβ-1. In turn, Zhang et al. [61] demonstrated that treatment with TGFβ-3 reduced the degree of fibrosis and the deposition of collagen fibers in the liver. Our results showed that the coculture of Matrigel in vitro decreased the expression of TGFβ-1 and increased the expression of TGFβ-3, which may have contributed to the reduced degree of fibrosis observed after MSC/Matrigel transplantation.
Matrigel supports MSC survival in vivo. Kaiser et al. [54] found that the survival rate of human neural progenitor cells on Matrigel-coated surfaces was better than that on surfaces without the supportive matrix. Wang et al. [62] transplanted neural stem cells/Matrigel subcutaneously into nude mice and found that the Matrigel supported the survival of these stem cells in vivo. Similarly, in our studies, the data demonstrated that Matrigel promoted MSC survival in ovarian tissues. The mechanisms of this supportive effect of Matrigel may include promotion of MSC proliferation and reduction of cell apoptosis.
In addition, the Matrigel scaffold can enhance the effects of MSC transplantation [63]. In this study, MSC/Matrigel significantly increased the ovarian index, restored the ovarian structure (increased the number of blood vessels), and enhanced the proliferation rate of granulosa cells, compared with the treatment with the MSCs alone.
Nevertheless, Matrigel is an animal-based product and is, therefore, not clinically viable. To overcome this limitation, researchers have focused on developing alternative scaffold materials. For example, fibrin and/or alginate matrices or 3D-printed crosslinked sharp-angled gelatin fibers can be used as scaffolds for reproduction [64 –68]. In addition, previous studies have indicated that decellularized ECM scaffolds can promote the tissue-specific differentiation of MSCs and be transplanted to improve or restore tissue function [69,70].
Although some studies have indicated that collagen scaffolds increase the short- and long-term retention of MSCs in tissues, no significant differences have been reported between MSCs and MSC/collagen treatments [37,57,71]. Additional studies are warranted to select a more effective scaffold for MSC transplantation. Furthermore, the potential underlying mechanisms through which such scaffolds affect MSC proliferation in vivo also requires further investigation.
In general, the novel findings of this study are as follows. First, the MSCs were labeled with lentiviral-GFP to enable tracing of the transplanted cells after injection in the ovary, and we tested the fate and biological behavior of the MSCs in vivo. In addition, we used Matrigel (Corning) as a scaffold for MSC transplantation to promote the effects of MSCs on ovarian structure and function and tested the effects of Matrigel on MSC survival in vivo. Third, our results indicated that Matrigel containing 5–7 ng/mL VEGF promoted vessel growth. This indicated that a scaffold combined with extra VEGF could better promote the repair of ovarian function in a POF model. Fourth, we scanned the entire cross-section of the ovary to observe MSC distribution, and the results showed that GFP+ MSCs in the MSC group were settled in the injection site, whereas the GFP+ MSCs in the MSC/Matrigel group were distributed throughout the ovary; the latter promoted MSC uptake in the whole ovary. The reason for this occurrence may be that Matrigel facilitates the migration of MSCs in vivo.
However, there were no significant differences between the MSC and MSC/Matrigel groups with regard to the tissue fibrosis ratio, number of follicles, body weight, and E2 levels. Thus, further studies are needed to explore the reasons for these results.
Conclusion
In this study, we demonstrated that the transplantation of MSC/Matrigel increased the number of follicles, decreased the tissue fibrosis ratio, improved the proliferation rate of granulosa cells, and increased the number of blood vessels in a mouse model of POF.
Footnotes
Acknowledgments
The authors thank all members of Prof. Zhang's laboratory for the valuable discussions.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This work was supported by National Natural Science Foundation of China (Nos. 81772773, 81672560), Jiangsu Provincial Medical Youth Talent (No. QNRC2016753), Suzhou Clinical Key Technology Project (No. LCZX201705), Gusu Medical Youth Talent (No. GSWS2019034), and The Project of Jiangsu Provincial Maternal and Child Health Association (No. FYX201709).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.