
Abstract
Osteoarthritis (OA) is the most common type of arthritis, afflicting millions of people in the world. Elevation of inflammatory mediators and enzymatic matrix destruction is often associated with OA. Therefore, the objective of this study was to investigate the effects of conditioned medium from periodontal ligament-derived stem cells (PDLSCs) on inflammatory and catabolic gene expressions of chondrocytes, synoviocytes, and meniscus cells under in vitro inflammatory condition. Stem cells were isolated from human periodontal ligaments. Conditioned medium was collected and concentrated 20 × . Chondrocytes, synoviocytes, and meniscus cells were isolated from pig knees and divided into four experimental groups: serum-free media, serum-free media+interleukin-1β (IL-1β) (10 ng/mL), conditioned media (CM), and CM+IL-1β. Protein content and extracellular vesicle (EV) miRNAs of CM were analyzed by liquid chromatography-tandem mass spectrometry and RNA sequencing, respectively. It was found that the IL-1β treatment upregulated the expression of IL-1β, tumor necrosis factor-α (TNF-α), MMP-13, and ADAMTS-4 genes in the three cell types, whereas PDLSC-conditioned medium prevented the upregulation of gene expression by IL-1β in all three cell types. This study also found that there was consistency in anti-inflammatory effects of PDLSC CM across donors and cell subcultures, while PDLSCs released several anti-inflammatory factors and EV miRNAs at high levels. OA has been suggested as an inflammatory disease in which all intrasynovial tissues are involved. PDLSC-conditioned medium is a cocktail of trophic factors and EV miRNAs that could mediate different inflammatory processes in various tissues in the joint. Introducing PDLSC-conditioned medium to osteoarthritic joints could be a potential treatment to prevent OA progression by inhibiting inflammation.
Introduction
Osteoarthritis (OA)
Because of their chondrogenic potential [12,13], there are accumulating reports of cell-based therapy of damaged cartilage using mesenchymal stem cells (MSCs) [14 –17]. Recent studies have demonstrated that intra-articular injection of MSCs improves function and pain of osteoarthritic joints [18 –21]. Furthermore, it has been shown that various paracrine factors and extracellular vesicles (EVs) (eg, exosomes) secreted by MSCs affect a variety of cellular activities (eg, enhancing cell proliferation and inhibiting apoptosis and inflammation) [22,23] and mediate tissue repair, including cartilage repair in injury and OA [24 –26]. Recent studies demonstrated that MSC-conditioned medium can inhibit inflammatory processes of chondrocytes [27,28] and reduce pain and prevent cartilage degradation in a collagenase-induced OA model [29]. Thus, introducing stem cell-conditioned medium to diseased joints could be a potential treatment to prevent tissue degradation by inhibiting apoptosis and inflammation and promoting tissue regeneration.
The cranial neural crest (NC) cells play an important role in the development of teeth by contributing to the formation of condensed dental mesenchyme, dental papilla, odontoblasts, dentine matrix, pulp, cementum, and periodontal ligaments (PDLs) [30]. The PDL is a soft connective tissue embedded between the tooth root and alveolar bone socket. The main function of the PDL is to anchor the tooth to the bone, respond to mechanical force on the teeth, and contribute to homeostasis of tooth and bone and repair of damaged tissue. Recent studies have shown that the PDL contains multipotent NC stem cells [31 –33]. Previous studies also demonstrated the capacity of PDL-derived stem cells (PDLSCs) to undergo chondrogenic differentiation [34 –41]. However, the anti-inflammatory capability of PDLSC-conditioned medium has not been investigated as potential therapeutics for OA.
Since OA is an inflammatory disease of the whole joint [2], which may involve all intrasynovial tissues, including articular cartilage, synovial membrane, meniscal fibrocartilage, and subchondral bone, the objective of this study was to investigate (1) the anti-inflammatory effects of conditioned medium of human PDLSCs on chondrocytes, synoviocytes, and meniscus cells under in vitro inflammatory conditions and (2) the potential anti-inflammatory factors and EV miRNAs in the conditioned medium of human PDLSCs using liquid chromatography-tandem mass spectrometry and miRNA sequencing, respectively.
Materials and Methods
Production of condition medium of PDL-derived stem cells
Stem cells were isolated from human PDL using the protocol described in our previous study [35]. Briefly, impacted third molars were obtained from healthy donors following routine medical procedures requiring their extraction at the University of Miami. The protocol was approved by University of Miami Institutional Review Board and conducted according to its guidelines. PDLs were enzymatically digested in a collagenase solution overnight and filtered through a 70 μm cell strainer to obtain single-cell suspensions. To isolate stem cells based on plastic adherence, cell suspension was plated in six-well plates at 1,000 cells per well and cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 100 U/mL penicillin/streptomycin (Invitrogen), and 0.1% v/v amphotericin B.
After 5 days of culture, nonadherent cells were discarded by changing medium. After 2 weeks of primary culture, stem cells were passaged into T175 flask at a density of 105 cells (Corning, New York, NY) and cultured at 37°C in a humidified atmosphere of 5% CO2. To produce conditioned media (CM), when reaching confluence, stem cells were cultured in 20 mL serum-free DMEM containing 1% antibiotic-antimycotic and 0.5% bovine serum albumin under the same environmental condition. After 48 h of culture, the CM were then collected. Following centrifugation at 1,000g for 5 min to remove unwanted cells, the CM were concentrated 20 × by centrifuging at 5,000g for 4 h using Macrosep Advance Centrifugal Devices (MWCO 3k; Pall Corporation, Port Washington, NY) and filtered through a 0.2 μm syringe filter. Concentrated serum-free media were made as a control. Concentrated media were stored at −80°C.
Anti-inflammatory experiment of chondrocytes, synoviocytes, and meniscus cells
Chondrocytes, synoviocytes, and meniscus cells were isolated from porcine knees [42]. Briefly, articular cartilage, meniscus, and synovial membrane tissues were obtained from the knee joints of 35–40 kg pigs obtained from the University of Miami Department of Surgery Tissue Sharing program (Institutional Animal Care and Use Committee approved source) within 2 h of death. Tissues were cut into small pieces and then digested in DMEM containing 10% FBS, 1% penicillin/streptomycin, 1 mg/mL collagenase (Worthington Biochemical, Lakewood, NJ), and 0.1 mg/mL protease (Sigma, St. Louis, MO) at 37°C for 16 h. After filtering cell suspension through a 70 mm cell strainer (BD Biosciences, San Jose, CA), cells were cultured in DMEM containing 10% FBS and 1% penicillin/streptomycin and expanded in monolayer culture at a seeding density of 105 cells in T-75 flask 37°C in a humidified atmosphere of 5% CO2.
During culture, media were changed every 2–3 days. For the treatment experiments, cells at passages 1 or 2 were plated in six-well plates with 2 × 105 cells per well and cultured for 48 h. To determine the effect of conditioned medium on gene expression, each cell type was divided into four groups: serum-free media (C), serum-free media+IL-1β (10 ng/mL) (C+), CM, and CM+IL1-β (CM+). For CM and CM+ groups, culture medium was supplemented with 0.25 mL of concentrated conditioned medium per mL culture medium. Each experimental group was cultured with specific treatments described above for 24 h. At the end of the experiment, gene expression of 18S, IL-1β, TNF-α, MMP13, and ADAMTS4 was examined using reverse transcription-polymerase chain reaction (RT-PCR).
RNA extraction and RT-PCR analysis of gene expression
Total RNA of cells was extracted using Tri-Reagent (Molecular Research Center, Cincinnati, OH) according to manufacturer's instructions. The RNA content of each sample was quantified by Qubit RNA HS assay kit (Life Technologies, Carlsbad, CA). Following RNA quantization, cDNA synthesis was performed by using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Grand Island, NY). Real-time PCR (StepOnePlus; Applied Biosystems) was performed utilizing a SYBRGreen kit to analyze gene expression of the following inflammatory and catabolic genes: 18S, IL-1β, TNF-α, MMP13, and ADAMTS4. The results were normalized by 18s gene as the endogenous control and calculated as relative expression to respective control group using the 2−ΔΔCt method. The primer sequences used in this study are summarized in Table 1.
Quantitative Real-Time Polymerase Chain Reaction Sequences
IL-1β, interleukin-1β; TNF-α, tumor necrosis factor-α.
Liquid chromatography-tandem mass spectrometry
The CM were shipped to Bioproximity, LLC (Chantilly, VA) for analysis of global proteomic profile. Tandem mass spectra were acquired using a quadrupole-Orbitrap mass spectrometer (Q-Exactive; Thermo Fisher) as described previously [43]. SearchGUI [44] was utilized to identify peptides based on mass spectra using simultaneous OMSSA [45] and X!Tandem [46] searches against the human sequence library from UniProt. Protein identification and validation were performed at a 1.0% false discovery rate using PeptideShaker [47]. Relative protein abundance was estimated by normalized spectral abundance factor [48].
EV miRNA sequencing
The CM were shipped to System Biosciences (Palo Alto, CA) for analysis of EV miRNA Sequencing. Ten milliliters of CM was centrifuged at 1,500g for 5 min to remove residual cells and debris. The supernatant was transferred to a new 50 mL conical tube for EV isolation. ExoQuick-TC (EXOTC10A-1; System Biosciences) was added to the supernatant at 1:5 ratio (ExoQuick:Supernatant), mixed gently, and allowed to incubate overnight at 4°C. After overnight incubation, the admixture was centrifugated at 1,500g for 30 min to recover EVs for total RNA isolation. EVs isolated from the samples were processed for total RNA isolation using the SeraMir Exosome RNA Purification Column kit (RA808A-1; System Biosciences) according to the manufacturer's instructions.
For each sample, 1 μL of the final RNA eluate was used for measurement of small RNA concentration by Agilent Bioanalyzer Small RNA Assay using Bioanalyzer 2100 Expert instrument (Agilent Technologies, Santa Clara, CA). Small RNA libraries were constructed with the CleanTag Small RNA Library Preparation Kit (L-3206; TriLink Biotechnologies, San Diego, CA) according to the manufacturer's protocol. The final purified library was quantified with High-Sensitivity DNA Reagents (G2933-85004; Agilent Technologies) and High-Sensitivity DNA Chips (G2933-85004; Agilent Technologies). The libraries were pooled, and the 140 to 300 bp region was size selected on an 8% TBE gel (EC6215; Invitrogen). The size selected library was quantified with High-Sensitivity DNA 1000 Screen Tape (5067-5584; Agilent Technologies), High-Sensitivity D1000 reagents (5067-5585; Agilent Technologies), and the TailorMix HT1 qPCR assay (TM-505; SeqMatic, Fremont, CA), followed by a NextSeq High Output single-end sequencing run at SR75 using NextSeq 500/550 High Output v2 kit (FC-404-2005; Illumina, San Diego, CA) according to the manufacturer's instructions.
Statistical analysis
One-way ANOVA with Turkey's HSD post hoc test (SPSS Statistics; IBM) was performed to examine differences in gene expression between experimental groups for individual cell type. Similarly, one-way ANOVA was used to compare the effects of the conditioned medium of PDLSCs from different donors or passages on gene expression of chondrocytes after normalizing each experimental group by its respective control group. Statistical significance was considered with P value <0.05 in all analyses.
Results
Anti-inflammatory effects of conditioned medium of PDLSCs
The gene expression analysis showed that the IL-1β treatment upregulated the expression of IL-1β, TNF-α, MMP-13, and ADAMTS-4 genes in chondrocytes, synoviocytes, and meniscus cells (Fig. 1). The conditioned medium of PDLSCs repressed IL-1β-induced upregulation of gene expression in all three cell types (Fig. 1).

Effects of PDLSC-conditioned medium on the gene expression of IL-1β, MMP-3, TNF-α, and ADAMST-4 in chondrocytes, synoviocytes, and meniscus cells with and without the treatment of 10 ng/mL IL-1β. Four treatment group: serum-free media (C), serum-free media+IL-1β (10 ng/mL) (C+), CM and CM+IL1-β (CM+). *P < 0.05. CM, conditioned media; IL-1β, interleukin-1β; PDLSC, periodontal ligament derived stem cell; TNF-α, tumor necrosis factor-α.
Variations among cell subcultures and donors
To evaluate the variability in the efficacy of conditioned medium of PDLSCs due to cell subculture or donor variation, CM of PDLSCs at passage 2, 5, and 10 from one donor (22-year-old female) and at passage 2 from two other donors (18-year-old female and 23-year-old male) were compared for their effects on gene expression of IL-1β-treated chondrocytes. It was found that the CM of PDLSCs at different passages from three different donors significantly downregulated gene expression of IL-1β-treated chondrocytes (Figs. 2 and 3). There were no significant differences in the anti-inflammatory effects of conditioned medium among three different passages (passage 2, 5, and 10) from the same donor (22-year-old female) and among three donors (22-year-old female, 18-year-old female, and 23-year-old male) for the same passage (passage 2) (Figs. 2 and 3).
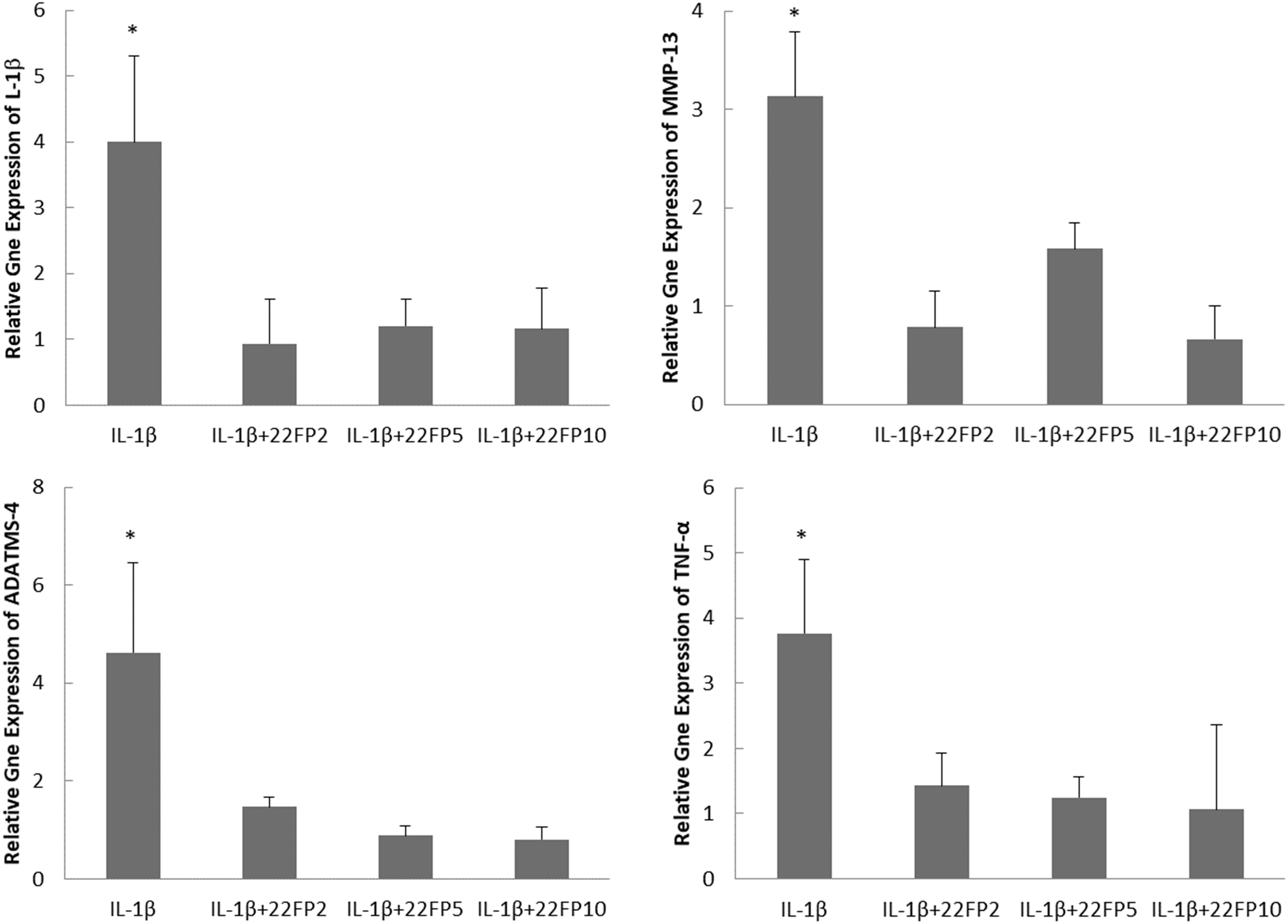
The effects of CM obtained from PDLSCs at passage 2 (22FP2), 5 (22FP5), and 10 (22FP10) from a 22-year-old female donor on the gene expression of IL-1β, MMP-3, TNF-α, and ADAMST-4 in chondrocytes under the treatment of 10 ng/mL IL-1β. *P < 0.05.
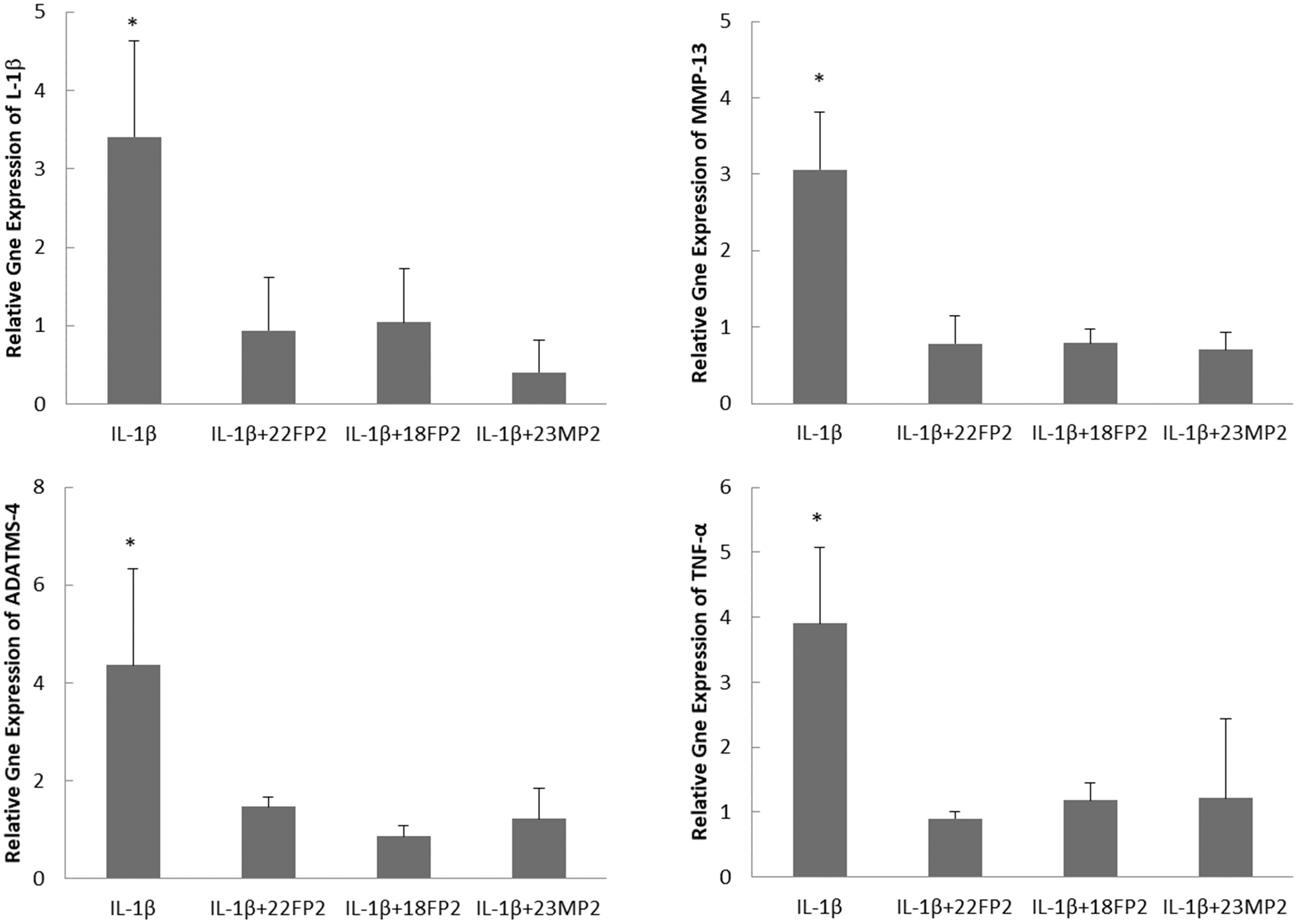
The effects of CM of PDLSCs at passage 2 from 3 donors (22-year-old female: 22FP2; 18-year-old female: 18FP2; and 23-year-old male: 23MP2) on the gene expression of IL-1β, MMP-3, TNF-α, and ADAMST-4 in chondrocytes under the treatment of 10 ng/mL IL-1β. *P < 0.05.
Proteomic analysis of PDLSC-conditioned medium
Mass spectrometry analysis of proteins was used to examine the content of abundant proteins in the conditioned medium of PDLSCs to identify potential anti-inflammatory factors. A total of 589 different proteins were identified in 5 CM of PDLSCs obtained from 3 different passages and 3 different donors as described in the previous section, including ECM proteins, enzymes, growth factors, and cytokines. For the CM of PDLSCs at passage 2, 5, and 10 from a 22-year-old female donor, there are 174 proteins in common (Fig. 4A). The CM of PDLSCs at passage 2 from 3 different donors had 52 proteins in common (Fig. 4A). Only 44 proteins were common to all 5 samples (Fig. 4A and Table 2), while 29 out of 44 common proteins were secretory proteins (Fig. 4B), most of which were ECM components (eg, FN, COL1A1, COL1A2, DCN, and COL6A3). Several secretory proteins (ie, MFG-E8, PTX3, and THBS1) have been reported to exhibit immunomodulatory actions [49 –51].

Common Proteins Detected in Five Samples (Three Different Passages and Three Different Donors) by Mass Spectrometry Analysis
EV miRNA sequencing
To examine whether exosomes in the conditioned medium of PDLSCs potentially exhibit anti-inflammatory effects, expression of EV miRNAs in the CM of PDLSCs at passage 2 from three donors (18-year-old female, 22-year-old female, and 23-year-old male) was analyzed. Overall, a total of 677 miRNAs were detected in exosomes from three samples. Top 25 miRNAs consisted of ∼80% of total miRNA reads for all samples (Fig. 5). Among the top 25 miRNAs identified from 3 samples, there are 14 common miRNAs, which are let-7a-5p, let-7b-5p, let-7g-5p, miR-21-5p, miR-24-3p, miR-25-3p, miR-92a-3p, miR-100-3p, miR-143-3p, miR-199a-3p/miR-199b-3p, miR-221-3p, miR-320a-3p, miR-423-5p, and miR-493-5p.
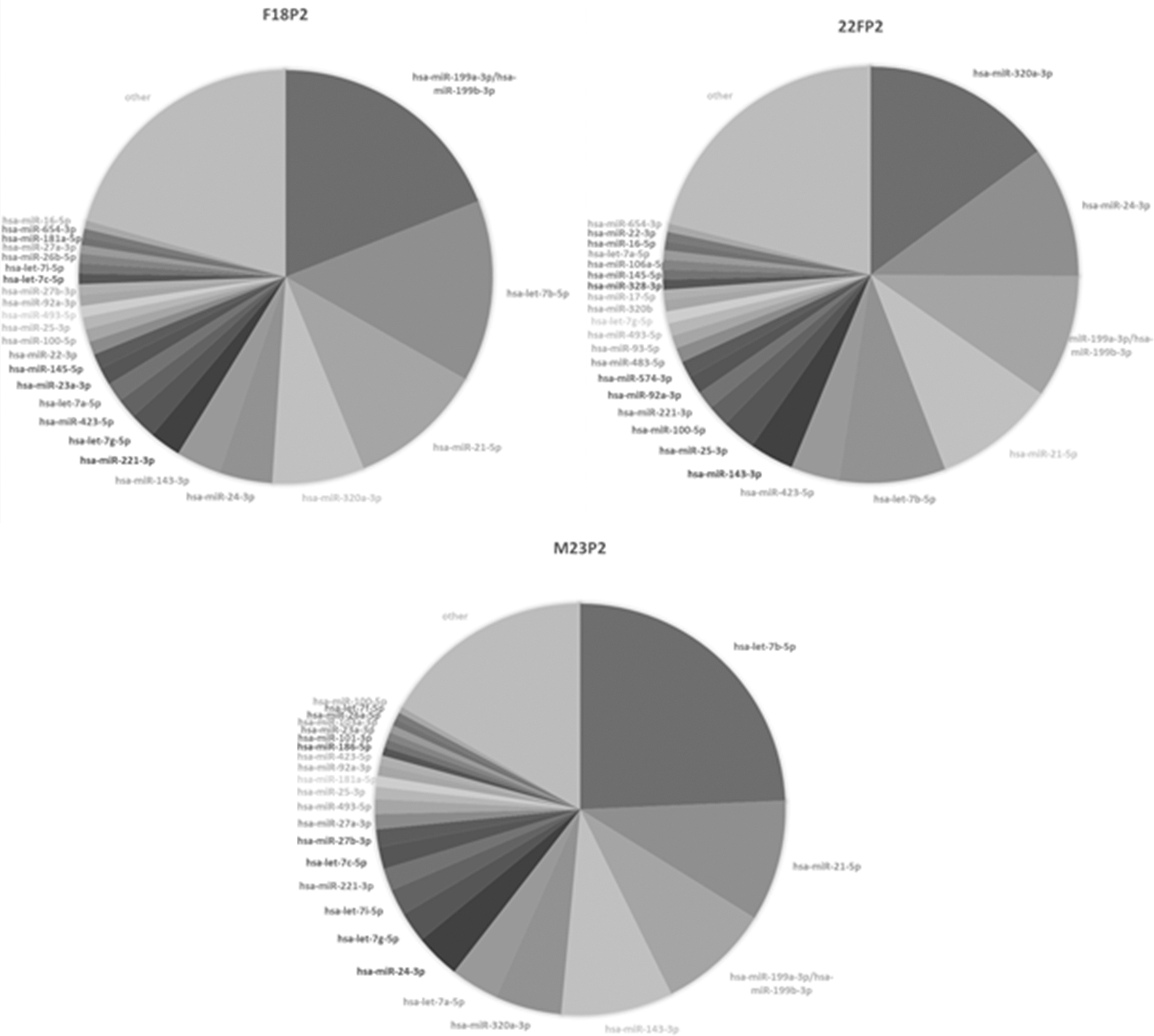
Pie diagrams of top 25 EV miRNA identified in the CM of PDLSCs from three different donors by RNA sequencing. The labels 22FP2, 18FP2, and 23MP2 represent the CM of PDLSCs at passage 2 from a 22-year-old female donor, 18-year-old female donor, and a 23-year-old male donor, respectively. EV, extracellular vesicle.
Discussion
This study demonstrated that the conditioned medium from PDLSCs showed promising effects on reducing inflammatory and catabolic responses of chondrocytes, synoviocytes, and meniscus cells to IL-1β. Our findings are consistent with those in recent studies, which demonstrated that MSC-conditioned medium can inhibit inflammatory processes of chondrocytes [27,28,52]. OA has been suggested as an inflammatory disease in which the elevated release of inflammatory mediators (eg, IL-1β) by intrasynovial tissues increases metalloproteinase synthesis, causing cartilage degradation [2,53]. IL-1β can sustain catabolic pathways in chondrocytes through autocrine and paracrine signaling [9]. Since our findings suggested that the conditioned medium obtained from PDLSCs can inhibit both autocrine and paracrine signaling of IL-1β in cartilage, synovium, and meniscus, the PDLSC-derived conditioned medium could be used to treat OA as a whole joint disease.
This study is the first to demonstrate that PDLSC CM contains several highly secretory anti-inflammatory and immunomodulatory proteins (ie, MFG-E8, PTX3, and THBS1) that have not been reported previously for PDLSCs and are consistently released by PDLSCs across donors and cell subcultures. In this study, the liquid chromatography-tandem mass spectrometry analysis revealed some abundant proteins in PDLSC-conditioned medium, which may have therapeutic potentials. One of them detected is milk fat globule-epidermal growth factor 8 (MFG-E8). Recent studies have shown an association between MFG-E8 and different inflammatory/autoimmune diseases such as rheumatoid arthritis, sepsis, ischemia/refusion injury, and Alzheimer's disease [54 –57]. The function of MFG-E8 was initially found to facilitate phagocytosis of apoptotic cells by binding with phosphatidylserine on apoptotic cells and αvβ3 integrin on phagocytes [58]. It has been recently demonstrated that MFG-E8 can suppress inflammation and reduce the expression of proinflammatory cytokines (eg, IL-1β and TNF-α) in several inflammatory diseases [56,57,59 –63]. Those previous studies support the findings of anti-inflammatory effects of PDLSC-conditioned medium in this study.
Macrophages, the major component of the mononuclear phagocyte system, may play a critical role in OA pathogenesis [64]. Macrophages can polarize into two subsets: proinflammatory M1 macrophages and anti-inflammatory M2 macrophages. The imbalance of macrophages of M1 and M2 subtypes in synovium is associated with the severity of OA [64]. It has been shown that MFG-E8 can reprogram macrophages from an M1 (proinflammatory) to an M2 (anti-inflammatory) phenotype [49]. In addition to MFG-E8, among the abundant proteins identified in PDLSC-conditioned medium, thrombospondin-1 and pentraxin-3 have also been shown to reduce inflammatory activities of macrophages [50,51]. Therefore, the PDLSC-conditioned medium could mitigate the progression of OA by targeting macrophages and macrophage-associated inflammatory pathways in synovium.
In addition to trophic factors, conditioned medium also contains EVs that have been shown to exhibit therapeutic potentials in OA. EVs are nano-sized vesicles that contain a variety of bioactive molecules, such as DNA, mRNAs, miRNAs, proteins, and lipids [22], which can be delivered into target cells either by direct fusion with the plasma membrane or activation of membrane-bound receptors. EV miRNAs have been shown to contribute to the therapeutic effects of EVs [22]. Among the highly expressed EV miRNAs detected in PDLSC-conditioned medium, miR-92a, miR-199a, miR-221, and miR-320a have been suggested as therapeutic targets for OA [65 –68]. It was found that OA cartilage exhibits a lower expression of miR-92a-3p and miR-221-3p than normal cartilage, while overexpression of miR-92a-3p or miR-221-3p in chondrocytes significantly suppressed IL-1β-upregulation of catabolic gene expression (eg, ADAMTS-4/5 and MMP-13) [65,68].
In OA chondrocytes, miR-199a directly suppressed IL-1β-induced overexpression of COX-2, which strongly contributes to the inflammation and cartilage degeneration in OA [66]. In addition, miR-24 has been recognized as one of anti-inflammatory miRNAs [69], which could inhibit apoptosis and promote proliferation of OA chondrocytes [70]. Furthermore, subchondral bone remodeling is also involved in OA development. It has been shown that expression of miR-199a-3p decreased in human OA subchondral bone, while overexpression of miR-199a-3p rescued the abnormal osteoblast gene expression and mineralization in OA subchondral bone [71]. Therefore, the PDLSC-conditioned medium may also exhibit therapeutic effects on OA joint through EV miRNAs.
This study demonstrated consistency in anti-inflammatory effects of PDLSC CM across donors and cell subcultures, which was supported by the consistent production of trophic factors and EV miRNAs by PDLSCs under the experimental conditions. It suggested that PDLSCs are a reliable stem cell source for producing therapeutic CM. Furthermore, OA has been recognized as a disease with chronic low-grade inflammation by which all tissues in the entire joint, including cartilage, subchondral bone, and synovium, are affected [72]. This study showed the anti-inflammatory effects of PDLSC CM on three different cell types in the knee joint, suggesting that PDLSC-conditioned medium is a cocktail of trophic factors and EV miRNAs that could mediate different inflammatory processes in various tissues in the joints.
One limitation of this study is that the protein assays were not performed to confirm the gene expression results since the anti-inflammatory effects of stem cell-conditioned medium have been verified previously at both mRNA and protein levels [28,73]. However, analysis of proteins expression will be included in the future studies when investigating the in vivo therapeutic effects of PDLSC CM. Since exosomes, a subtype of EV, could be responsible for the therapeutic effect of stem cell secretomes [22,23], the characterization and anti-inflammatory activities of exosomes in the PDLSC-conditioned medium will also be examined in our future studies. Moreover, the content of secretome in conditioned medium is affected by the culture conditions [74]. Thus, optimizing culture condition may help produce more effective conditioned medium.
Conclusion
The conditioned medium derived from PDLSCs demonstrated anti-inflammatory effects on various cell types in the joints. It was also found that PDLSCs consistently produced the anti-inflammatory factors and EV miRNAs at high levels, which make them a reliable stem cell source to produce a therapeutic cocktail for treating inflammatory diseases such as OA.
Footnotes
Acknowledgments
The authors thank Victoria Amirniroumand, Daniela Cusco, Michael Santiago-Castro, and Rhea Kumar for their assistance with cell experiments.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the College of Dentistry Alumni Endowment Fund Award from the University of Tennessee Health Science Center and partially funded by the Dominican Republic Ministry of Higher Education Science and Technology (National Fund for Innovation, Scientific and Technological Development; code 2016-2017-2A7-002).