
Abstract
Prostate cancer (PCa) is a malignant tumor that originates in the male genitourinary system. Downregulation of death-associated protein kinase 1 (DAPK1) is closely related to PCa. Little is known about the functional role of DAPK1 in regulating cancer stem cell (CSC)-like characteristics of PCa cells, and we have conducted research on this topic. Compared with tumor-adjacent normal tissues, DAPK1 was severely downregulated in tumor tissues of PCa patients. DAPK1 expression was also reduced in PCa cell lines with respect to that in normal prostate cells. Moreover, we sorted PCa-CSCs (PCa-CD133+ cells) from PCa cells. PCa-CD133+ cells also exhibited a reduced DAPK1 level and elevated levels of stem cell markers (CD44, OCT4, and SOX2). DAPK1 knockdown promoted sphere formation and enhanced the proportions of PCa-CD133+/PCa-CD133− cells. Inhibition of DAPK1 also accelerated migration and invasion of PCa-CD133+ cells. In addition, DAPK1 interacted with zinc finger E-box-binding homeobox-1 (ZEB1) and repressed ZEB1 expression in PCa-CD133+ cells. DAPK1 suppressed Hippo/YAP signaling pathway by interacting with ZEB1. Finally, we generated a tumor xenograft model to verify the effect of PCa-CD133+ cells following DAPK1 overexpression on tumor growth of PCa. DAPK1 overexpression inhibited tumor growth of PCa and repressed the expression of ZEB1, YAP, and TAZ in the tumor tissues of PCa mice. In conclusion, reduced DAPK1 expression promoted stem cell-like characteristics of PCa cells through activating ZEB1 via Hippo/YAP signaling pathway. Taken together, this work sheds lights on the potential of DAPK1 as a target for PCa therapeutics from bench to clinic.
Introduction
Prostate cancer (PCa) is a type of malignant tumor that originates in the male genitourinary system [1]. Due to the high mortality and recurrence rate, PCa is considered as a serious threat to the health of men [2]. Given that patients with advanced PCa that cannot be treated surgically, current treatments mainly include endocrine and/or chemotherapy, that is, androgen deprivation therapy [3]. PCa and its therapeutics have shown adverse effects such as sexual dysfunction as well as urinary symptoms that negatively affect the psychological health of PCa patients [4]. However, in the advanced stage of PCa, androgen receptor is usually continuously activated in the absence of androgens [5]. To prevent and control the advanced and metastatic tumors, it is urgent to screen biomarkers for developing novel treatment strategies of PCa [6,7]. The inactivation or overactivation of tumor suppressor genes or oncogenes in various cancers, including PCa, is usually a vital factor in the development of various cancers.
In the past decades, cancer stem cells (CSCs) provide a novel perspective for cancer treatment [8,9]. CSCs possess the abilities of self-renewal and multidifferentiation and function as initiating cells to regulate the occurrence, spread, and recurrence of tumors [10]. CSCs exert a decisive role in tumorigenesis, maintaining tumor homeostasis, and increasing tumor invasion and metastasis [11]. CSCs have been reported in lung cancer, breast cancer, liver cancer, and many other types of tumors, which can be separated and purified by cell surface markers [12]. PCa CSCs have also been reported to be involved in the recurrence and metastasis of PCa, which induces chemoresistance [13,14]. Therefore, CSC theory may become a novel research direction for radical treatment of PCa.
A typical tumor suppressor gene named death-associated protein kinase 1 (DAPK1), a serine/threonate kinase, has been shown to be involved in the occurrence, development and metastasis of various cancers [15 –17]. Tong et al. have analyzed the PCa-related gene expression profiles, demonstrating that the methylation levels of DAPK1 are increased, and DAPK1 protein level is decreased in PCa [18]. The Cancer Genome Atlas database has verified that the difference in DAPK1 expression is statistically significant. Decreased DAPK1 expression is related to the resistance of cancer cells to cetuximab and erlotinib [19]. In addition, DAPK1 knockdown activates the transcription factor zinc finger E-box-binding homeobox-1 (ZEB1), thereby promoting epithelial-mesenchymal transition (EMT) and stem cell-like features in colon cancer [20]. DAPK1-ZEB1 jointly regulates TGF-β and WNT pathways. As an activator that regulates multiple signaling pathways, ZEB1 forms a complex with the effector YAP of the Hippo pathway, and thus activating the downstream genes of Hippo/YAP signaling pathway [21,22]. However, little is known about the functional role of DAPK1 and ZEB1 in regulating stem cell-like characteristics in PCa cells. Therefore, we carried out this study to investigate whether DAPK1/ZEB1 could regulate stem cell-like characteristics of PCa via Hippo/YAP signaling pathway.
Materials and Methods
Participants
A total of 60 PCa patients who underwent radical resection of PCa were recruited at the Affiliated Hospital of Nantong University. Sixty pairs of PCa tissues and tumor-adjacent normal tissue samples were collected from 60 PCa patients during the surgery. All PCa patients had not received any treatments before the surgery. The tumor tissues were diagnosed with PCa by pathological examination. The participants were informed and gave written consent. All protocols were carried out in accordance with the Declaration of Helsinki and authorized by the Ethics Committee of The Affiliated Hospital of Nantong University (Approval No. 2015-K005-01).
Cell culture
Human normal prostate cell line (RWPE2) and PCa cell lines (PC-3, DU145 and LNCaP) were purchased from China Center for Type Culture Collection (Wuhan, China). Cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Waltham, MA) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin (Sangon Biotech, Shanghai, China) at 37°C with 5% CO2.
Cell transfection
For DAPK1 or ZEB1 overexpression, full length of DAPK1 or ZEB1 was subcloned into pcDNA3.1 vector, generating the vectors named pcDNA3.1-DAPK1 or pcDNA3.1-ZEB1. The empty vector pcDNA3.1-NC was used as negative control (NC). Short hairpin RNA specifically targeting DAPK1 (sh-DAPK1) and scrambled shRNA were synthesized for DAPK1 knockdown. These vectors and oligonucleotides were obtained from GenePharma (Shanghai, China). Cells were transfected with vectors or oligonucleotides using Lip2000 (Invitrogen, Carlsbad, CA) following the instruction of manufacturer.
Cell sorting of CD133+ cells
PCa CSCs (PC-3-CD133+ and DU145-CD133+ cells) were sorted form PCa cells utilizing Miltenyi Biotec CD133 Cell Isolation Kit (Miltenyi Biotec, Auburn, CA) as previous study described [23]. PC-3 and DU145 cells (1 × 106 cells) were magnetically labeled with 1 μL CD133 (Miltenyi Biotec). Cell suspension was loaded onto the magnetic activated cell sorting column and then sorted on a MoFlo Cell Sorter (Beckman Coulter, Brea, CA). The effluent population of CD133+ and CD133- cells was collected, respectively. The sorted CD133+ cells were separated over a second column. Subsequently, the purity of CD133+ cells was examined by staining with CD133-FITC (1:20; #1-1339-42; Thermo Fisher Scientific, Carlsbad, CA) on a FACSCalibur (BD Biosciences).
Flow cytometry
PC-3 and DU145 cells were collected and washed with phosphate buffered saline (PBS) for three times. Cells at a concentration of 2.0 × 106/100 μL were incubated with CD133-FITC (1:20; #1-1339-42; Thermo Fisher Scientific) at 4°C in the dark for 30 min. After washing with PBS for several times, the proportions of CD133+ cells were analyzed on a FACSCalibur (BD Biosciences, San Jose, CA).
Quantitative real-time polymerase chain reaction
Total RNAs were extracted from cells and tumor tissues by TRIzol reagent (Invitrogen). RNA integrity was estimated by 1.0% agarose gel electrophoresis. Reverse transcription and polymerase chain reactions (PCRs) were carried out using TransScript® II Two-Step RT-PCR SuperMix (TransGen, Beijing, China) on a Real-Time PCR Instrument (Applied Biosystems, Carlsbad, CA). Relative expression of genes was analyzed through the 2−ΔΔCt method and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primer sequences (5′-3′) were designed as follows: DAPK1-F: 5′-ACAGCACGAGCACGTAGCA-3′; DAPK1-R: 5′-GGGGGTTTTCCCGGATC-3′. CD44-F: 5′-GACCACACAAAACAGAACCAGG-3′; CD44-R: 5′-GGGTTGAAGAAATCAGTCCAGG-3′. OCT4-F: 5′-CCGGAATTCGCCACCATGGACACCTGTTTCG-3′; OCT4-R: 5′-ATAAAAACATTCATAAACCACTTAACACA-3′. SOX2-F: 5′-CTCCGGGACATGATCAGC-3′; SOX2-R: 5′-CTGGGACATGTGAAGTCTGC-3′; GAPDH-F: 5′-GTGGACCTGACCTGCCGTCT-3′; GAPDH-R: 5′-GGAGGAGTGGGTGTCGCTGT-3′.
Western blot
Total proteins were extracted from cells and tissues using Total Protein Assay Extraction Kit (Solarbio, Beijing, China) as the protocol described. The protein concentration was examined using BCA Protein Assay Kit (Beyotime, Shanghai, China). Protein samples (25 μg) were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis, and transferred onto nitrocellulose membranes (Merck Millipore, Billerica, MA) for 1 h. The nitrocellulose membranes were blocked with 5% skimmed milk, and then incubated with primary antibody as indicated, including DAPK1 (1:1,000; #25136-1-AP; Proteintech), ZEB1 (1:1,000; #21544-1-AP; Proteintech), YAP (1:2,000; #13584-1-AP; Proteintech), TAZ (1:2,000; #66500-1-Ig; Proteintech), or CD133 (1:2,000; #66666-1-Ig; Proteintech,) at 4°C for 12 h. Membranes were then labeled with the secondary antibody horseradish peroxidase-conjugated IgG (HRP-IgG). β-actin antibody (1:2,000; #20536-1-AP; Proteintech) was used as an internal reference for normalization. The immunoblotting bands were analyzed by ImageJ software.
Immunohistochemical staining
Paraffin sections of PCa tissues and tumor-adjacent normal tissues were made through fixation and embedding. After deparaffinization and rehydration, the 4-μm paraffin sections were treated with Target Retrieval Solution (Dako, CA) for antigen retrieval. The sections were incubated with 0.3% H2O2 for 15 min, and then incubated with primary antibody as indicated, including DAPK1 (1:100; #25136-1-AP; Proteintech), ZEB1 (1:100; #21544-1-AP; Proteintech), YAP (1:100; #13584-1-AP; Proteintech), TAZ (1:500; #66500-1-Ig; Proteintech), or CD133 (1:1,000; #66666-1-Ig; Proteintech) at 4°C for 12 h. Subsequently, the secondary antibody HRP-IgG was labeled with the sections. Sections were then stained with 3, 30-diaminobenzidine and counterstained with hematoxylin. Finally, the sections were observed under an Olympus optical microscope (Olympus, Tokyo, Japan).
Sphere formation assay
Sphere formation assay was carried out according to previous report [24]. In brief, cells at a concentration of 500 cells/well were seeded into 6-well ultralow cluster plates (Corning, NY). Cells were cultured in DMEM supplemented with 2% B27 (Invitrogen), 20 ng/mL human epidermal growth factor (Sigma-Aldrich, St. Louis, MO), 20 ng/mL human basic fibroblast growth factor (Sigma-Aldrich), 5 μg/mL insulin (Sigma-Aldrich), and 0.4% bovine serum albumin (Sigma-Aldrich). Ten days of postculture, the spheres were observed under an optical microscope.
Transwell assay
To examine the ability of invasion, cells were seeded into the upper of the Transwell chambers precoated with Matrigel (BD Biosciences). The upper of Transwell chambers were cultured in FBS-free DMEM. The lower chamber contained DMEM (500 μL) with 10% of FBS. Twenty-four hours postcultivation, the noninvading cells on the upper side of chamber were gently removed. The invading cells were fixed with paraformaldehyde and then dyed with 0.1% crystal violet. The protocol of transwell migration assay was identical with the invasion assay, except that the chambers were not precoated with Matrigel. Finally, five visual fields were randomly selected to observe the invading and migrating cells under an inverted light microscope (Olympus).
Coimmunoprecipitation assay
The interaction between DAPK1 and ZEB1 in the PC-3-CD133+, DU145-CD133+, PC-3, and DU145 cells was estimated through coimmunoprecipitation (Co-IP) assay following transfection of pcDNA3.1-DAPK1 (PC-3-CD133+-DAPK1, DU145-CD133+-DAPK1, PC-3-DAPK1, and DU145-DAPK1 cells). Cells were washed with PBS for three times, and then lysed in RIPA Lysis Buffer (Beyotime). Cell lysates were collected and incubated with the primary antibodies, including DAPK1 (#MA1-24696; Thermo Fisher Scientific) or ZEB1 (#A301–922A; Thermo Fisher Scientific) at 4°C for 3 h. Then, Protein A/G Plus-Agarose (Santa Cruz Biotechnology, Santa Cruz, CA) was incubated with the cell lysates. The immunoprecipitates were washed with PBS, along with a portion of the cell lysates (Input), and were analyzed by western blot (WB) assay using DAPK1 (1:1,000; #25136-1-AP; Proteintech) and ZEB1 (1:1,000; #21544-1-AP; Proteintech) antibodies.
Xenograft mouse model
Male BALB/c nude mice aged 4 weeks (weighting 18 ± 2 g) (SLAC, Shanghai, China) were housed under specific pathogen-free conditions at 18°C–25°C and a 12/12 h light/dark cycle. All animal protocols were performed in accordance with the Guide for the Care and Use of Laboratory Animals, and authorized by the Ethics Committee of The Affiliated Hospital of Nantong University (Approval No. S20191118). Mice were subcutaneously injected with 200 μL (2.0 × 106/mL) of pcDNA3.1-DAPK1-transfected PC-3-CD133+ cells/pcDNA3.1-DAPK1-transfected DU145-CD133+ cells (DAPK1 group), pcDNA3.1-NC-transfected PC-3-CD133+ cells/pcDNA3.1-NC-transfected DU145-CD133+ cells (NC group), or untransfected PC-3-CD133+ cells/DU145-CD133+ cells (Control group) through the right fossa axillaris. Mice were euthanized to separate the tumor xenografts; the weight and volume of tumor xenografts were measured every week for 4 weeks. Tumor volume (mm3) = 1/2 × width2 (mm2) × length (mm).
Statistical analysis
Every assay was repeated for three times. Data were analyzed using SPSS 20.0 software (SPSS, Inc., Chicago, IL) and expressed as the mean ± SD. Statistical differences between two groups (or among multiple groups) were analyzed using the Student's t-test (or one-way analysis of variance). Difference P < 0.05 was considered statistically significant.
Results
DAPK1 was downregulated in the tumor tissues of PCa patients and PCa cell lines
We initially assessed the mRNA and protein expression levels of DAPK1 between tumor tissues and tumor-adjacent normal tissues of PCa patients. Data obtained from quantitative real-time PCR (qRT-PCR), WB, and immunohistochemical (IHC) assays showed that DAPK1 expression was severely decreased in the tumor tissues of PCa patients with respect to those in tumor-adjacent normal tissues (Fig. 1A–C). We also found that PC-3, DU145, and LNCaP cells exhibited a reduced DAPK1 mRNA and protein levels compared with those in control prostate cell line RWPE2 (Fig. 1D, E). These data indicated that DAPK1 expression was decreased in the tumor tissues of PCa patients and in PCa cell lines.
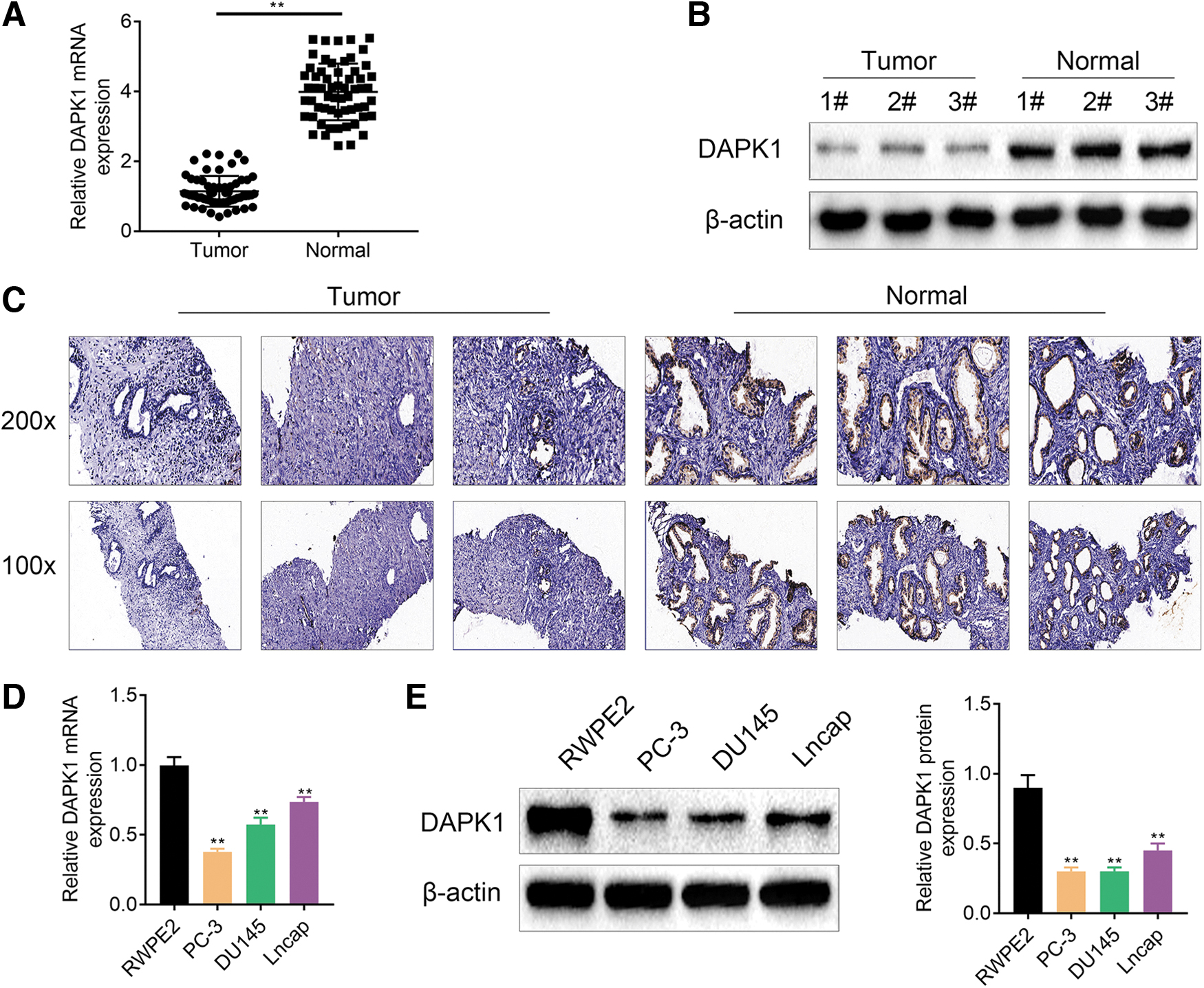
DAPK1 expression was decreased in PCa cell lines and tissues. The mRNA and protein expression of DAPK1 in the PCa tissues and tumor-adjacent tissues (N = 60) was assessed by qRT-PCR
DAPK1 expression was inhibited in PCa CSCs
To determine whether DAPK1 could affect stem cell-like characteristics in PCa, we sorted PCa CSCs from PC-3 and DU145 cells. Then, we assessed the mRNA and protein expression of DAPK1 and stem cells-related genes (CD44, OCT4, and SOX2) by qRT-PCR and WB. Figure 2A showed that DAPK1 mRNA and protein were significantly downregulated in the PC-3-CD133+ and DU145-CD133+ cells. Compared with those in CD133− PCa cells, the mRNA and protein levels of CD44, OCT4, and SOX2 were obviously enhanced in the PC-3-CD133+ and DU145-CD133+ cells (Fig. 2A, B). To sum up, PCa CSCs displayed a downregulation of DAPK1.
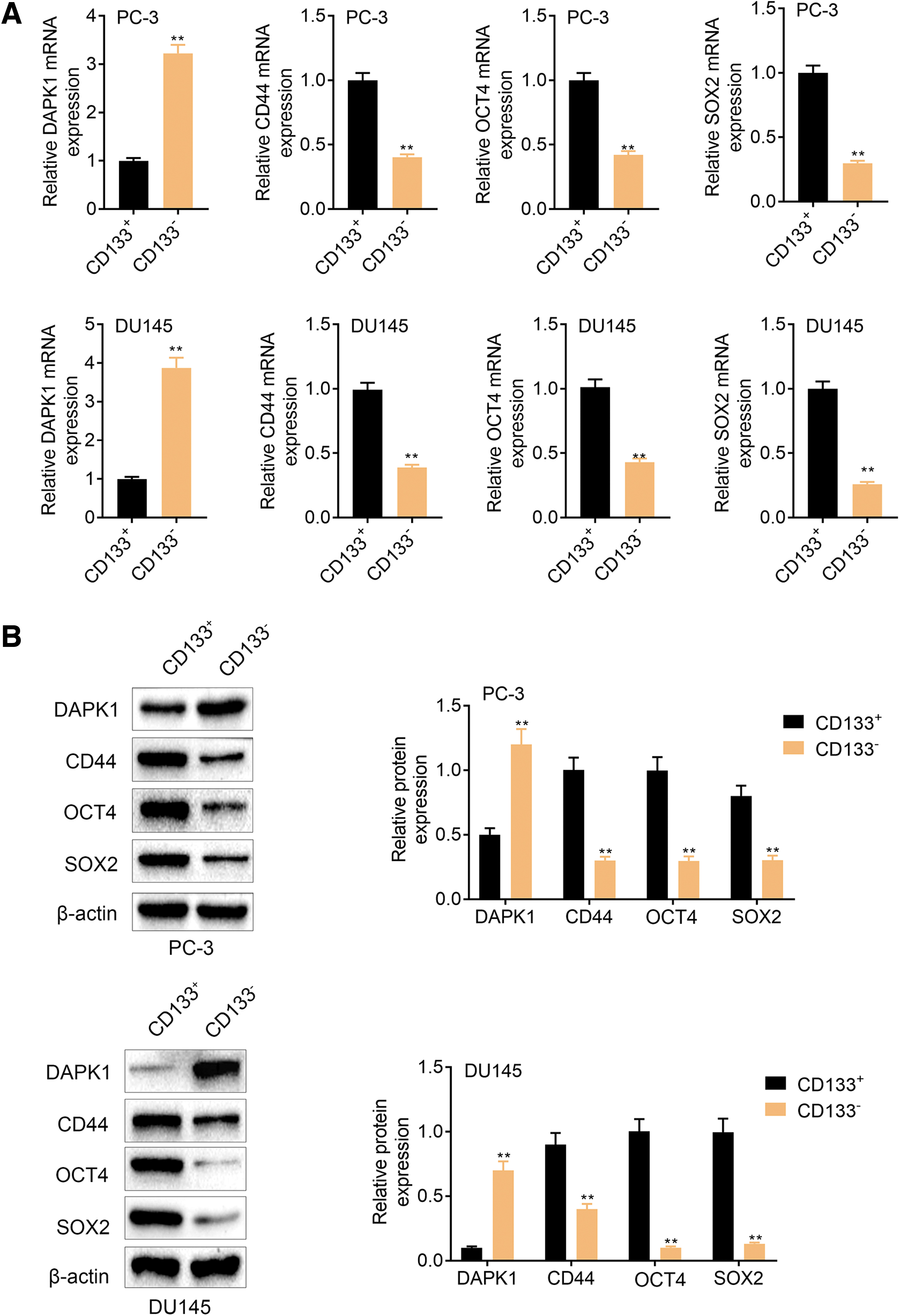
DAPK1 expression was decreased in PCa CSCs. The qRT-PCR
DAPK1 knockdown promoted stem cell-like phenotype in the PCa cells
Next, we overexpressed or silenced DAPK1 in PC-3-CD133+ and DU145-CD133+ cells. The qRT-PCR and WB results showed that DAPK1 mRNA and protein expression was notably increased in the PC-3-CD133+ and DU145-CD133+ cells expressing pcDNA3.1-DAPK1. Reversely, the expression of DAPK1 mRNA and protein was severely decreased in the PC-3-CD133+ and DU145-CD133+ cells following transfection of shDAPK1 (Fig. 3A, B). Meanwhile, DAPK1 overexpression caused reduced levels of CD44, OCT4, and SOX2, whereas DAPK1 deficiency led to an upregulation of CD44, OCT4, and SOX2 in the PC-3-CD133+ and DU145-CD133+ cells (Fig. 3A, B). Moreover, sphere formation assays assessed the impact of DAPK1 on sphere formation in PCa cells, showing that DAPK1 overexpression inhibited sphere formation in PC-3 and DU145 cells. However, DAPK1 silencing promoted sphere formation in the PC-3 and DU145 cells (Fig. 3C, D). We also found that DAPK1 overexpression decreased the CD133+ cell proportions in the PC-3 and DU145 cells. However, DAPK1 deficiency boosted the proportions of PC-3-CD133+ and DU145-CD133+ cells in PC-3 and DU145 cells (Fig. 3E). Therefore, DAPK1 silencing accelerated stem cell-like phenotype in the PCa cells.

DAPK1 knockdown regulated stem cell-like phenotype in the PCa CSCs cells. The qRT-PCR
DAPK1 knockdown promoted migration and invasion of PCa CSCs and PCa cells
We then estimated the functional role of DAPK1 in regulating migration and invasion of PCa cells by transwell assays. Figure 4A revealed that migration and invasion of PC-3-CD133+ and DU145-CD133+ cells were suppressed under DAPK1 overexpression, which could be accelerated by DAPK1 silencing. Moreover, DAPK1 overexpression suppressed the migration and invasion abilities in PC-3 and DU145 cells, whereas DAPK1 deficiency led to an increase of migration and invasion in PC-3 and DU145 cells (Fig. 4B). Thus, DAPK1 knockdown promoted migration and invasion of PCa CSCs and PCa cells.

DAPK1 knockdown promoted migration and invasion of PCa cells and PCa CSCs.
Reduced DAPK1 level induced ZEB1 expression in PCa CSCs and PCa cells
To further determine the role of DAPK1 and ZEB1 in PCa, we verified the relationship between DAPK1 and ZEB1 by Co-IP assay. Figure 5A showed that DAPK1 interacted with ZEB1 in PC-3-CD133+ and DU145-CD133+ cells. There were also binding sites between DAPK1 and ZEB1 in PC-3 and DU145 cells (Fig. 5B). In addition, DAPK1 overexpression reduced ZEB1 protein expression, whereas DAPK1 inhibition enhanced ZEB1 protein level in the PC-3-CD133+ and DU145-CD133+ cells (Fig. 5C). DAPK1 overexpression caused a decreased ZEB1 protein level, whereas deficiency of DAPK1 led to an increase of ZEB1 protein in PC-3 and DU145 cells (Fig. 5D). To sum up, these data illustrated that DAPK1 deficiency activated ZEB1 expression in PCa CSCs and PCa cells.
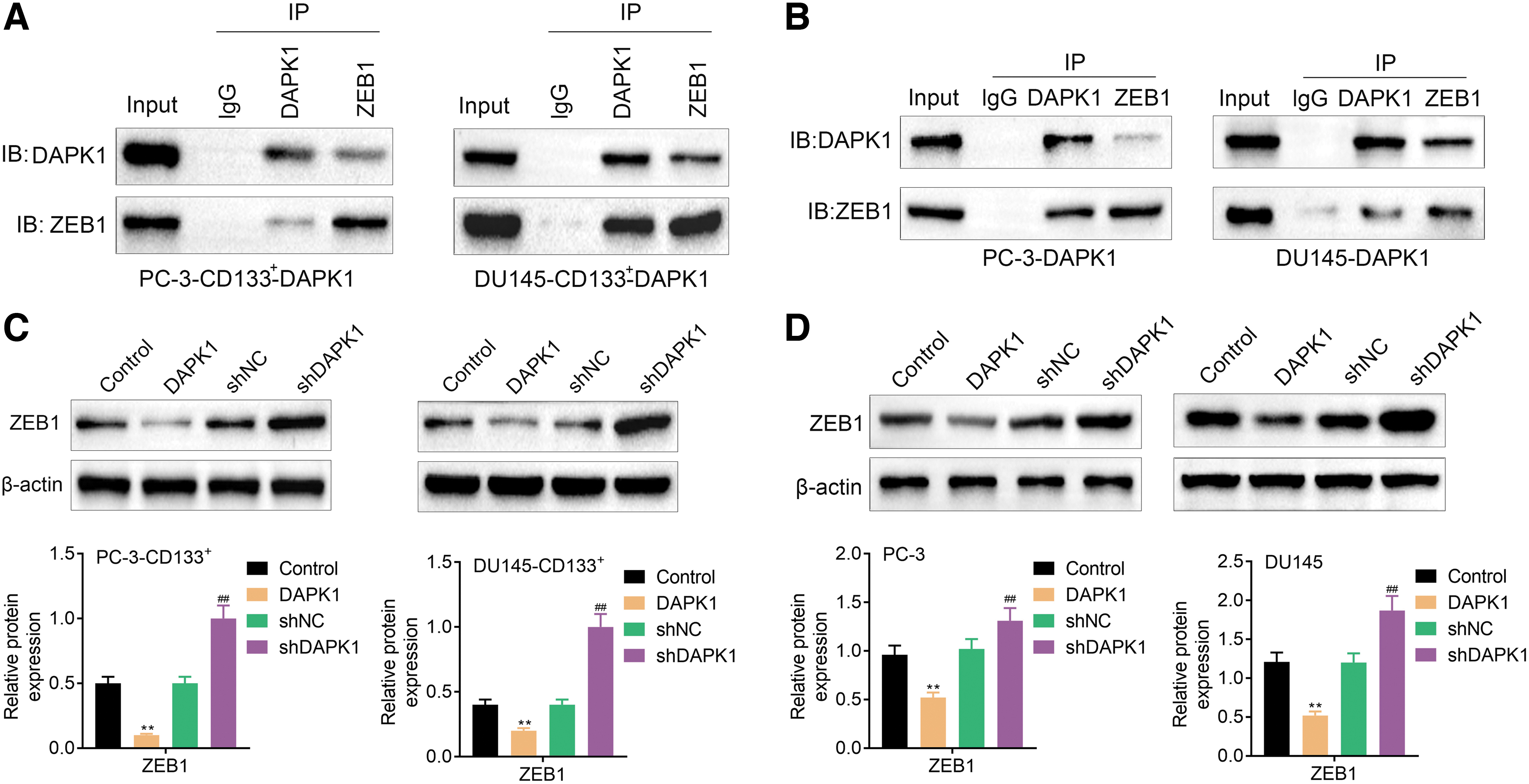
DAPK1 knockdown promoted ZEB1 expression by interacting with ZEB1 in PCa CSCs and PCa cells.
DAPK1 inhibited Hippo/YAP signaling pathway by targeting ZEB1 in PCa CSCs and PCa cells
Furthermore, we verified whether DAPK1/ZEB1 can regulate Hippo/YAP signaling pathway in PCa CSCs and PCa cells. WB results verified that the protein expression of ZEB1, YAP, and TAZ was severely decreased in PC-3-CD133+ and DU145-CD133+ cells upon DAPK1 overexpression. ZEB1 overexpression significantly enhanced the protein levels of ZEB1, YAP, and TAZ in PC-3-CD133+ and DU145-CD133+ cells. However, ZEB1 overexpression reversed the impact of DAPK1 on ZEB1, YAP, and TAZ expression in PC-3-CD133+ and DU145-CD133+ cells (Fig. 6A). We also found that ZEB1, YAP, and TAZ proteins were downregulated in PC-3 and DU145 cells in cells expressing pcDNA3.1-DAPK1. ZEB1 overexpression caused increased protein levels of ZEB1, YAP, and TAZ in the PC-3 and DU145 cells. DAPK1 upregulation-mediated elevated ZEB1, YAP, and TAZ expression in PC-3 and DU145 cells was reversed by ZEB1 elevation (Fig. 6B). Taken together, these data showed that DAPK1 inhibited Hippo/YAP signaling pathway by targeting ZEB1 in PCa CSCs and PCa cells.
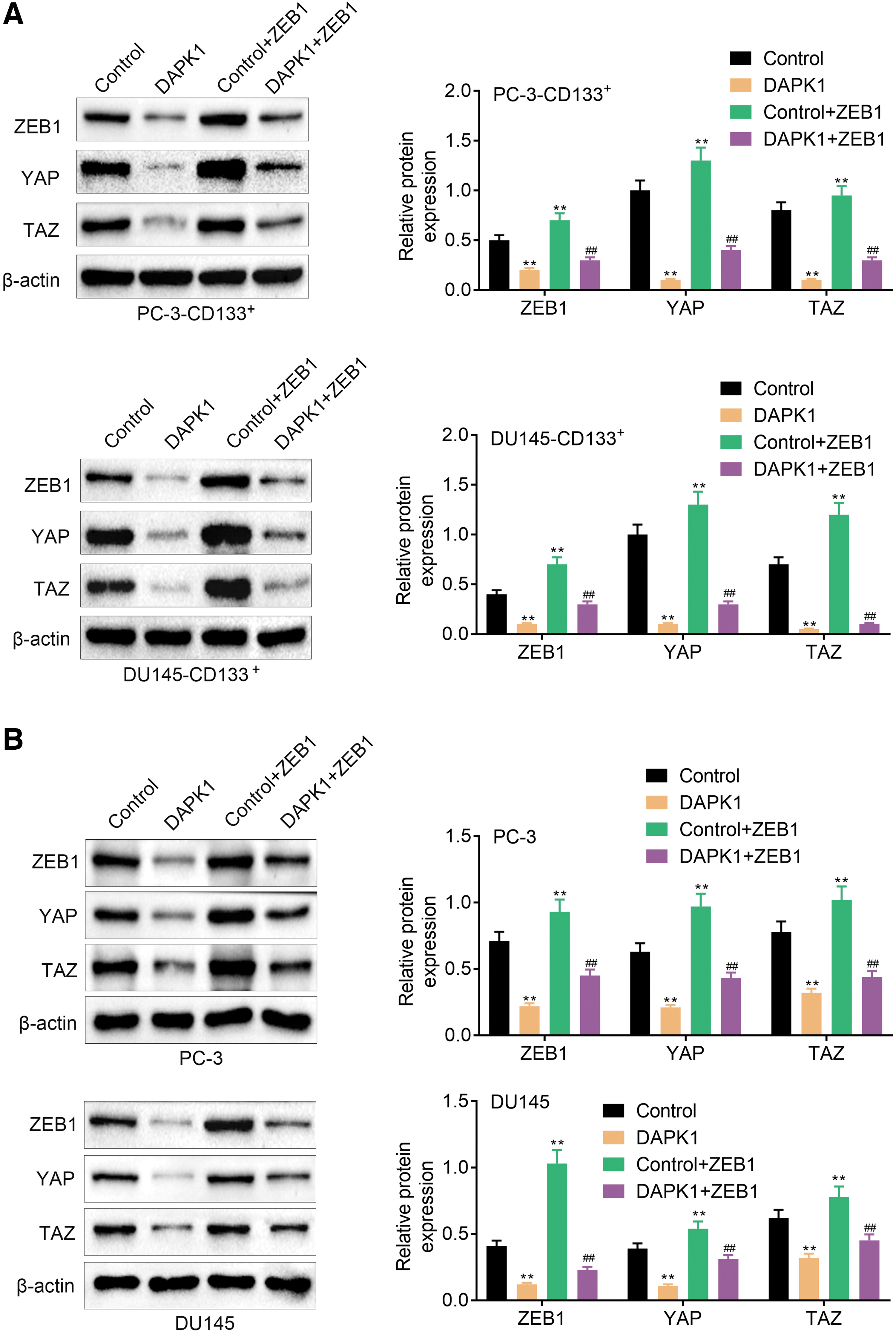
DAPK1 inhibited Hippo/YAP signaling pathway by repressing ZEB1 expression.
DAPK1 overexpression inhibited tumor growth in mice
Finally, we constructed a tumor xenograft mouse model through inoculation of DAPK1-overexpressed PCa-CD133+ cells to verify the influence of DAPK1 overexpression on tumor growth in vivo. We found that inoculation of DAPK1-overexpressed PC-3-CD133+ cells significantly repressed the PCa tumor growth in mice (Fig. 7A). Inoculation of DAPK1-overexpressed DU145-CD133+ cells also severely suppressed the tumor volume and weight in PCa mouse model (Fig. 7B). In summary, in vivo data demonstrated that DAPK1 overexpression inhibited tumor growth in PCa mouse model.
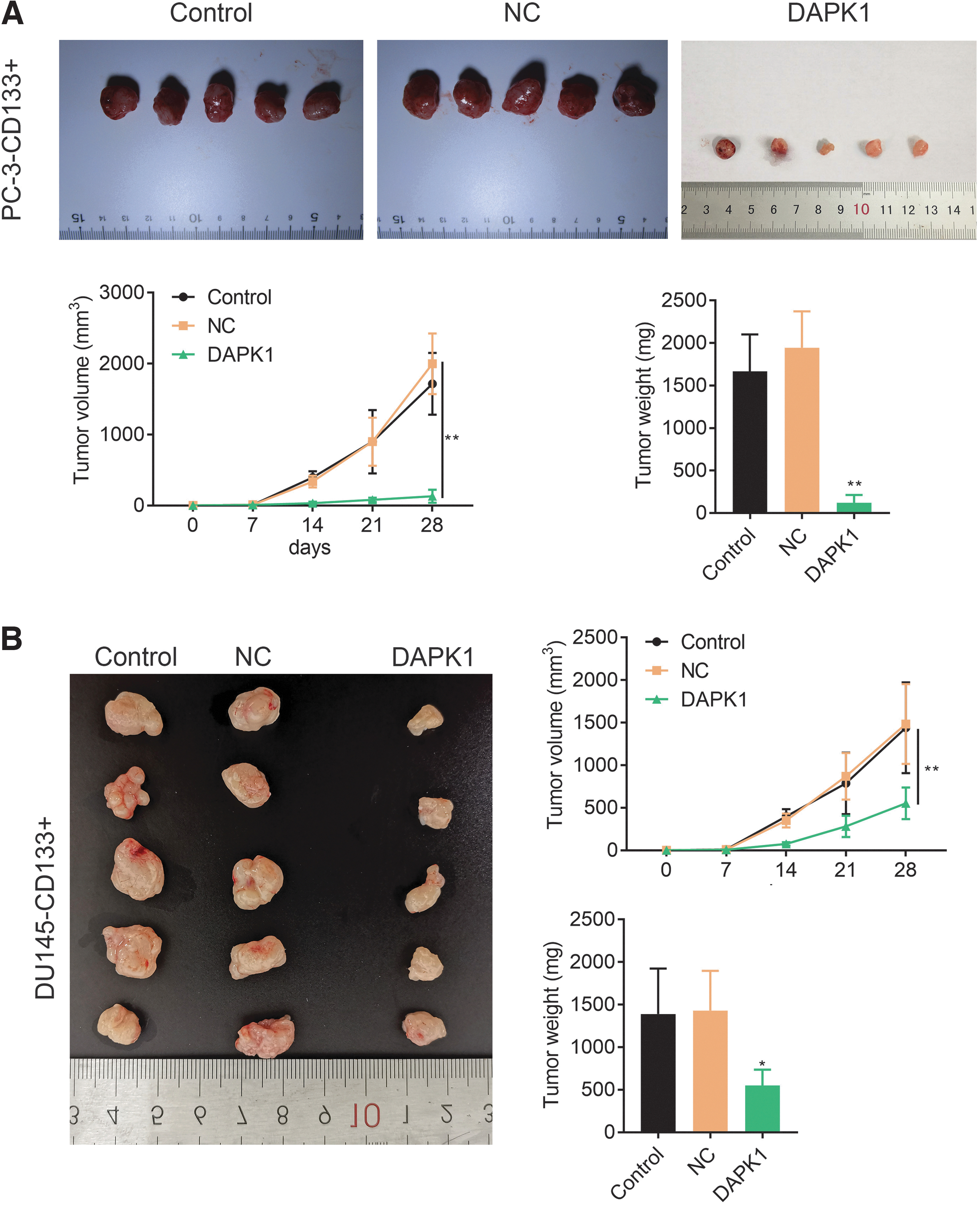
DAPK1 overexpression inhibited tumor growth of PCa in mice.
DAPK1 overexpression repressed the expression of ZEB1, YAP, and TAZ in the tumor tissues of PCa mice
In addition, we examined the expression of DAPK1, ZEB1, YAP, and TAZ in the tumor tissues of PCa mice through IHC and WB assays. As shown in Fig. 8A and B, DAPK1 level was significantly increased, while the expression of ZEB1, YAP, and TAZ was suppressed in the tumor tissues of mice in the presence of DAPK1-overexpressed PC-3-CD133+ cells. DAPK1-overexpressed DU145-CD133+ cells of inoculation also enhanced the expression of DAPK1, and reduced the expression of ZEB1, YAP, and TAZ in the tumor tissues of PCa mice (Fig. 8A, B). Thus, DAPK1 overexpression repressed the levels of ZEB1, YAP, and TAZ in the tumor tissues of PCa mice.
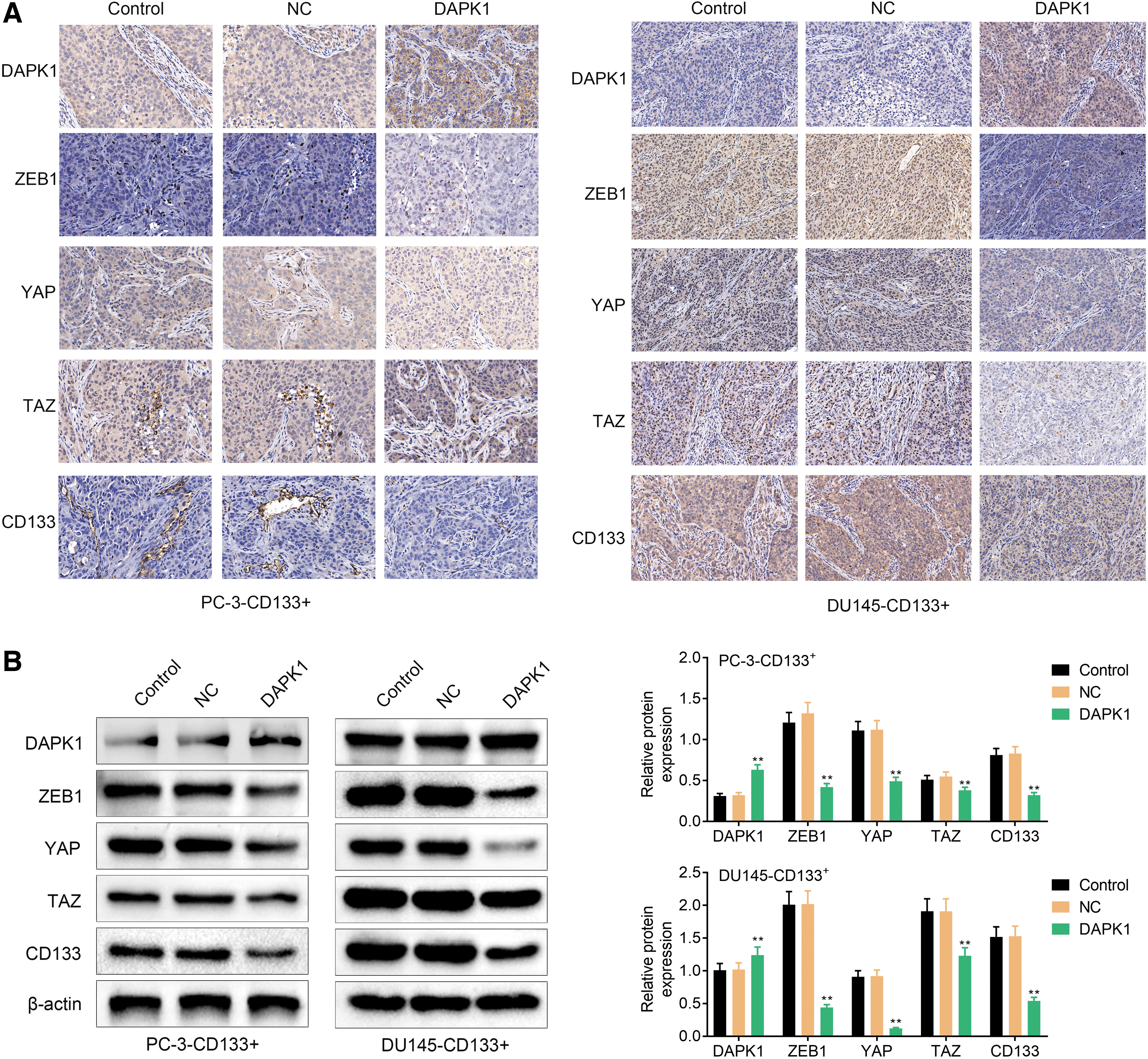
DAPK1 overexpression repressed the expression of ZEB1, YAP, and TAZ in the tumor tissues of PCa mice.
Discussion
DAPK1 exerts a vital role in the multiple types of biological functions, such as autophagy, apoptosis, immune response, autoimmune diseases, neuronal aging, wound repair, and progression of various cancers [25 –27]. At the same time, DAPK1 functions as a tumor suppressor gene to inhibit cell canceration in the early stage of tumor development [28]. For example, DAPK1 mRNA level is decreased in liver cancer, and low expression of DAPK1 is reported to be associated with the shorter survival of liver cancer patients [29]. DAPK1 inhibits the invasion and migration of pancreatic cancer cells through the miR-82/ROCK-1/RhoA signaling pathway, thereby exerting an antitumor effect in pancreatic cancer [30]. DAPK1 possesses antimetastasis functions in colorectal cancer, and DAPK1 deficiency promotes tumor invasion in colorectal cancer cells [17]. Kilinc et al. have confirmed that DAPK1 methylation levels are significantly enhanced in PCa tissues, indicating DAPK1 methylation may participate in the progression of PCa [31]. This study further explored the functional role of DAPK1 in PCa. We found that DAPK1 expression was obviously decreased in PCa cells and the tumor tissues of PCa patients, suggesting that DAPK1 knockdown was closely associated with PCa progression.
As the progenitor cells of tumor cells, CSCs play a crucial role in various malignant tumors. CD133 is confirmed as the most common marker of CSCs [32], which is closely associated with the proliferation of PCa cells [33]. Kanwal et al. have used CD133 as a CSC biomarker to isolate CD133+ cells from PCa cells [23]. Consistently, we separated PCa CSCs from PC-3 and DU145 cells to investigate whether DAPK1 could regulate stem cell-like characteristics in PCa. CD44, OCT4, and SOX2 are important markers of stem cells, and these molecules have been used to estimate the subpopulations of CSCs in cancer researches [34,35]. In the present work, our data revealed that DAPK1 expression was reduced in PCa CSCs. PCa CSCs exhibited an increase level of stem cell markers (CD44, OCT4, and SOX2). Moreover, DAPK1 knockdown positively regulated CD44, OCT4, and SOX2 expression, and notably promoted the ability of sphere formation in PCa CSCs. DAPK1 deficiency also boosted the proportions of CD133+ cells in PCa CSCs. DAPK1 knockdown accelerated migration and invasion of PCa CSCs and PCa cells. Thus, data obtained from this study demonstrated that DAPK1 knockdown promoted stem cell-like phenotype and accelerated the progression of PCa.
ZEB1 has been reported to take part in the progression of PCa. Wang et al. have found that the increased ZEB1 level is closely associated with EMT and phenotypes of CSCs in PCa [36]. ZEB1 drives EMT to mediate docetaxel-resistance in PCa [37]. Song et al. have confirmed that ZEB1 expression is elevated in PCa cells, and inhibition of ZEB1 accelerates apoptosis and restrains proliferation and invasion in PCa cells by repressing ERK1/2 signaling pathway [38]. This study deeply investigated the mechanism of action of DAPK1 and ZEB1 in the progression of PCa. We found that DAPK1 interacted with ZEB1, and thus inhibited ZEB1 expression in PCa CSCs and PCa cells. ZEB1 activated Hippo/YAP signaling pathway. DAPK1 suppressed Hippo/YAP signaling pathway by interacting with ZEB1. In addition, inoculation of DAPK1-overexpressed PCa CSCs suppressed tumor growth in PCa mouse model. DAPK1 overexpression also repressed ZEB1, YAP, and TAZ levels in the tumor tissues of PCa mice. A previous study has revealed that FRMD6 represses tumor growth of PCa by inhibiting Hippo/YAP signaling pathway [39]. Thus, these data demonstrated that DAPK1 inhibited Hippo/YAP signaling pathway by repressing ZEB1 expression, which contributed to inhibit the progression of PCa.
Conclusion
This work illustrated that DAPK1 knockdown promoted stem cell-like characteristics of PCa cells through activating ZEB1 via the Hippo/YAP signaling pathway, thereby accelerating the progression of PCa, shedding lights on DAPK1 as a potential biomarker for monitoring the progression and drug development for PCa treatment from bench to clinic.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the China Jiangsu Province Postdoctoral Science Foundation (grant no. 2014M139951).