
Abstract
Lung aging alters the intrinsic structure of the lung and pulmonary surfactant system and increases the mortality and morbidity due to respiratory diseases in elderly individuals. We hypothesized that lung aging results from an insufficiency of type II alveolar epithelial cells (AECIIs) in the lung tissue. Sirtuin 3 (SIRT3) is a member of the sirtuin family of proteins that promote longevity in many organisms. Increased SIRT3 expression has been linked to an extended life span in humans. Hence, we speculated that the overexpression of SIRT3 may help to ameliorate lung senescence and improve AECII function. AECIIs were isolated from young and old patients with pneumothorax caused by pulmonary bullae. The expression of SIRT3, manganese superoxide dismutase, and catalase, as well as cell function and senescence indicators of young and old AECIIs, was measured before and after SIRT3 overexpression. After SIRT3 overexpression, the aged state of old AECIIs improved, and antiapoptotic activity, proliferation, and secretion were dramatically enhanced. Surfactant protein C (SPC), which is secreted by AECIIs, reduces alveolar surface tension, repairs the alveolar structure, and regulates inflammation. SPC deficiency in patients is associated with increased inflammation and delayed repair. SIRT3 deacetylated forkhead box O3a, thereby protecting mitochondria from oxidative stress and improving cell function and the senescent state of old AECIIs. These findings provide a possible direction for aging-delaying therapies and interventions for diseases of the respiratory system.
Introduction
In the modern world, respiratory diseases have become a main cause of mortality and morbidity among elderly people, surpassed only by cardiovascular diseases and cancer [1 –3]. Lung senescence may lead to the destruction of the alveolar structure, a decrease in lung compliance, impairment in the gas exchange function, and a reduction in the secretion of surfactant protein C (SPC), which will cause a further decrease in lung function and even respiratory insufficiency [4 –6]. Therefore, the prevention of respiratory failure and lung infection in elderly people is very important for improving their quality of life and prolonging their life span.
Because an ideal or effective therapeutic option that cures end-stage lung disease is currently unavailable (except for lung transplantation), strategies designed to reverse the senescence process of lung tissue and improve the function of senescent cells may represent key factors for reducing mortality in the elderly population [7].
As stem cells of the alveolar epithelium, type II alveolar epithelial cells (AECIIs) are able to proliferate and differentiate into type I alveolar epithelial cells (AECIs) during the process of alveolar homeostasis and self-repair after injury [8]. AECIIs are large squamous epithelial cells that constitute the air–blood barrier [9 –11]. Furthermore, AECIIs secrete SPC. The mature form of SPC is highly hydrophobic. It plays an important role in reducing the alveolar surface tension and maintaining the stability of the alveolar structure in the process of breathing [12 –14]. In addition to its biophysical roles, SPC also contributes to immune defense, to some extent [15]. Indeed, its deficiency is associated with increased inflammatory reactions and delayed repair [16,17].
To date, no study has reported a correlation between the modification of AECII function and age. This study mainly aimed to investigate methods to reverse the senescent status of AECIIs and to enhance their function.
Sirtuin 3 (SIRT3) is a deacetylase in the sirtuin family and is located in mitochondria [18,19]. It plays a key role in promoting the human life span, improving the status of senescent organs, and enhancing the function of stem cells [20 –22]. After deacetylation, forkhead box O3a (FoxO3a) can enter the cell nucleus and interact with the promoters of the genes that encode catalase (CAT) and manganese superoxide dismutase (MnSOD), hence reducing the level of reactive oxygen species (ROS). Moreover, the increased expression of SIRT3 has consistently been shown to upregulate the expression levels of CAT and MnSOD, which protect cells from ROS-induced oxidative damage [23,24].
In this study, the correlations between the SIRT3 expression level and lung senescence and cell function were investigated using lung tissue and AECIIs from patients with pneumothorax caused by pulmonary bullae. Our study is the first to show that upregulated SIRT3 expression may represent a new therapeutic target to combat respiratory diseases in the elderly population during the natural aging process.
Materials and Methods
Collection of lung tissue specimens
Specimens of human lung tissues were obtained from patients with pneumothorax undergoing lung surgery in the 2nd Affiliated Hospital of Harbin Medical University. Samples from 40 males and 40 females were used in the current study, which is based in China. All the subjects had provided written informed consent. All study protocols were approved by the Ethics Committee of Harbin Medical University to ensure that they were in compliance with the requirements of the Declaration of Helsinki. Senescent lung tissue and AECIIs were excised from patients over 65 years of age (mean age 70 ± 4.5 years), while young AECIIs were collected from patients between 18–35 years of age (mean age 27 ± 6.5 years). Characteristics of the donors are provided in Table 1.
The Clinical Indicators of the Donors
Isolation and culture of AECIIs
The specimens of human lung tissue were sliced and rinsed with balanced salt solution (BSS) buffered with HEPES until the slices became clear. The slices were then placed in an aseptic container with 50 mL of BSS. After gentle mixing, the tissue-containing solution was filtered through a 150-μm mesh (BD Falcon, San Jose, CA). The remaining tissue was rinsed for a minimum of three times and incubated for 40 min in a 37°C water bath in a mixture of 40 mL of BSS, 3 mL of trypsin (Sigma, Saint Louis, MO), and 300 μL of elastase (Worthington, Lakerwood, NJ). The enzyme activity was inhibited with 30 mL of DMEM/F-12 (Invitrogen, Grand Island, NY), 10 mL of fetal bovine serum (FBS), and 1 mL of 10,000 U/mL DNase I. The solution was successively filtered through cell strainers (75- and 40-μm) to collect the suspension.
After centrifugation at 300 g for 10 min at room temperature, the cell pellet was resuspended in a mixture containing 22.5 mL of DMEM/F-12, 22.5 mL of small airway epithelial growth medium (SAGM; Lonza, Walkersville, MD), 5 mL of FBS, and 1 mL of 10,000 U/mL DNase I to separate the macrophages and fibroblasts. The suspended cells were then plated in 100-mm Petri dishes (10 mL per dish). After incubation at 37°C for 150 min, which is an ideal duration confirmed by our previous experiments, the macrophages and fibroblasts that adhered to the plates were isolated from the alveolar epithelial cells, as the latter require a longer time to attach to the plates. The AECII cells were then harvested and centrifuged at 300 g for 10 min at room temperature. The cell pellets were resuspended in 2 mL of BSS, and possible contaminants related to macrophages were removed using anti-CD14 microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany). The purified AECIIs were centrifuged at 300 g for ∼10 min at room temperature, resuspended in 2 mL of SAGM, and incubated in Petri dishes (60-mm diameter) at 37°C. The medium was replaced once every 2 days. The cells used in this study were cultured for 10 days [25].
Histological assessment
The lung tissues were fixed with paraformaldehyde (PFA), embedded in paraffin, and sectioned at a thickness of ∼5 μm. The sections were subjected to hematoxylin and eosin (H&E) staining, and six randomly selected fields were examined (100 × magnification) to evaluate lung morphology.
Gene expression level
The RNA was extracted from the lung tissue and AECIIs in six-well plates using TRIzol reagent (Life Technologies, Carlsbad, CA), followed by treatment with DNase I (Sigma-Aldrich, St. Louis, MO). Reverse transcription was carried out using the AccuPower RocketScript RT PreMix Test Kit (Bioneer, Alameda, CA). Gene expression levels in the sample were determined by real-time polymerase chain reaction (PCR) using the AccuPower 29 GreenStar qPCR Master Mix (Bioneer) Test Kit (Bio-Rad, Hercules, CA). The primers were purchased from Bioneer Corporation and are shown in Table 2.
Primers Used for Quantitative Real-Time Polymerase Chain Reaction
CAT, catalase; MnSOD, manganese superoxide dismutase; SIRT3, sirtuin 3; SPC, surfactant protein C.
Protein expression
The lung tissue homogenates were lysed with RIPA lysis and extraction buffer (Beyotime, Haimen, China) to extract the proteins. The protein expression levels of SIRT3, CAT, MnSOD, and FoxO3a were measured by western blot. Nuclear proteins were extracted with an extraction kit (Solarbio, Beijing, China). The protein contents of all samples were determined using a Bicinchoninic Acid Test Kit (Beyotime, Shanghai, China). Equal amounts of protein extracted from the samples were electrophoretically separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred onto polyvinylidene fluoride membranes (Millipore, Bedford, MA). The membranes were then blocked with 5% skim milk and incubated with specific primary antibodies for 12 h at 4°C.
The following antibodies were used: rabbit anti-SIRT3 monoclonal antibody (Cell Signaling Technology, Danvers, MA), rabbit anti-SOD2 monoclonal antibody (MnSOD; OriGene, Beijing, China), rabbit anti-CAT monoclonal antibody (Abcam, Cambridge, United Kingdom), rabbit anti-FoxO3a monoclonal antibody (Abcam), rabbit anti-SPC monoclonal antibody (Abcam), rabbit anti-histone H3 polyclonal antibody (Abcam), and mouse anti-GAPDH antibody (ZSGB-BIO, Beijing, China), which was used as a loading control. The membranes were incubated with the secondary antibody for 1 h at room temperature. The secondary antibodies used were goat anti-rabbit immunoglobulin G (IgG) conjugated with horseradish peroxidase (HRP; ZSGB-BIO) and goat anti-mouse IgG-HRP (ZSGB-BIO). The immunoblots were finally analyzed using Enhanced Chemiluminescence Detection Reagent (Beyotime, Shanghai, China).
Evaluation of cell proliferation, differentiation, and secretion
The separated cells were plated in six-well plates (104 cells per plate) and cultured in SAGM. On the 10th day, the cells were harvested by trypsinization and counted to evaluate their proliferative ability. Cell counting was performed on the fifth day after transfection. Based on the results of the differentiation and secretion analyses, SPC was found to be the unique substance secreted by AECIIs, and recombinant aquaporin-5 (AQP-5) was found to be the unique substance secreted by AECIs. Cellular differentiation was detected by performing immunofluorescence staining for SPC and AQP-5, and secretion was detected by measuring the expression level of SPC mRNA and protein.
Enzymatic activity of CAT and MnSOD
Total proteins were extracted from the harvested cells to measure enzyme activity using MnSOD and CAT Test Kits (Beyotime, Shanghai, China) according to the manufacturer's instructions.
Immunofluorescence staining of lung tissue sections and AECIIs
The lung tissue was isolated, embedded, and frozen in liquid nitrogen. The tissues were then cut into 5-μm-thick pieces with a freezing microtome, and the slices were fixed with 4% PFA for 15 min and incubated with 5% bovine serum albumin (BSA) for 1 h to allow cell permeabilization and to block nonspecific protein–protein interactions. The tissue slices were then incubated with a primary anti-SPC antibody (Abcam) at a concentration of 5 μg/mL for 12 h at 4°C, followed by the secondary antibody (DyLight® 488 goat anti-rabbit IgG (H+L)) for 1 h. The nuclei were stained with 1.43 μM 4,6-diamidino-2-phenylindole (DAPI). AECIIs were plated at 1 × 104 cells/well in a 24-well plate, incubated for 24 h, and then fixed with 4% PFA for 10 min. The cells were incubated with 1% BSA for 1 h to allow cell permeabilization and block nonspecific interactions. The cells were then incubated with antibodies against SPC, p16 and, p21 (all Abcam) for 12 h at 4°C. The secondary antibody was used according to the manufacturer's instructions.
The cell nuclei (blue) were stained with 1.43 μM DAPI. The number of positively stained cells was recorded, and the average number of cells per microscopic field of view (0.4 mm2) in six fields per sample was calculated.
Senescence-associated β-galactosidase staining
Senescence-associated β-galactosidase (SA-β-gal) staining was performed using the SA-β-gal Test Kit (Beyotime, Shanghai, China). The number of positively stained cells was recorded, and the average number of cells per microscopic field of view (0.4 mm2) in six fields per sample was calculated.
Oxidative stress test
The cells were treated for 1 h with hydrogen peroxide (H2O2) diluted with SAGM (1 mmol/L). The SAGM was obtained from Lonza.
Cellular apoptosis and survival assay
Cellular apoptosis was detected using the terminal deoxyribonucleotidyl transferase-mediated dUTP-biotin nick-end labeling (TUNEL) technique (Roche, Indianapolis, IN). The nuclei of the apoptotic cells and nonapoptotic cells were stained green and blue, respectively. The number of apoptotic cells was recorded by calculating the average number of TUNEL-positive nuclei in six fields (0.4 mm2 per field) per sample. Cell Counting Kit-8 (CCK8; Dojindo, Kumamoto, Japan) was used to determine cell survival after undergoing oxidative stress.
Adenovirus vector construction
Adenoviruses encoding SIRT3 were purchased from Beijing SyngenTech Co., Ltd., China. SIRT3 was synthesized and transferred into the pAD-CMV-MCS-IRES-EGFP vector to obtain the pAD-CMV-Sirt3-IRES-EGFP plasmid. The adenoviruses were generated in the Ad293 cell line and purified by freeze–thaw cycles, followed by centrifugation in caesium chloride. The virus titer was measured by immunization with the coat protein. The Ad-GFP vector was used as the negative control.
Adenovirus transfection
The cells were randomized into four groups, namely, the old (O), O+Ad-GFP (Ad-GFP), O+Ad-SIRT3 (Ad-SIRT3), and young (Y) groups, to study the role of SIRT3 in the process of AECII senescence. After ∼1 week of culturing, the cells were dissociated using trypsin, reseeded into six-well plates at a density of ∼2,000 cells/cm2, and cultured at 37°C. After 2 days, the cells from the Ad-GFP and Ad-SIRT3 groups were rinsed thrice with phosphate-buffered saline and then treated at 37°C for 2 h with 200 pfu/cell of the virus in 1 mL of infection medium. Medium was added after infection, and the cells were cultured at 37°C in a humidified incubator containing 5% CO2 for 48 h before they were collected for RNA or protein extraction.
Statistical analyses
Statistical analyses of the data were performed using GraphPad Prism 6.0 (GraphPad, La Jolla, CA). The results are presented as means ± standard deviations. Two-tailed Student's t-test was used to compare data between two groups, and one-way or two-way analysis of variance was used to evaluate the differences among multiple groups. P < 0.05 was considered a statistically significant difference.
Results
Differences in the alveolar cavity structure and cellular function between young and old lung tissues
Lung tissues collected from young and old patients were visualized using H&E staining to investigate the functions of young and old human AECIIs. The alveolar cavity of the senescent lung tissue was fused, the alveolar spaces were enlarged and mixed, and the alveolar septum was thinner in lung tissue from the old group (Fig. 1A). Immunofluorescence staining showed a decreased number of AECIIs in the old group (Fig. 1B, C). Furthermore, mRNA and protein levels of SPC were significantly downregulated in the old lung tissue (Fig. 1D, E).

Differences in the alveolar cavity structure and cell function between young and senescent lung tissues.
When the young and old AECIIs were cultured, the cells showed a cuboidal appearance and formed clusters that increased in size until forming a monolayer (Fig. 1F). No difference was observed in the appearance of young and old AECIIs after 10 days of culture. At the cellular level, the proliferation of old AECIIs was lower compared with the young cells on day 4, and the difference was still statistically significant on day 10 (Fig. 1G). The secretory function of the old cells (Fig. 1J, K) was also significantly lower. However, no differences in the differentiation time or ratio were observed between the two groups (Fig. 1H, I).
Changes in senescence indicators in young and old AECIIs
Senescence-related biomarkers were examined in AECIIs harvested from young and old lung tissues to determine whether old AECIIs were senescent. Higher SA-β-gal activity was detected in old AECIIs (Fig. 2A) than in young AECIIs. Immunofluorescence staining indicated greater numbers of p16- and p21-positive old AECIIs than in the young cells (Fig. 2B, C). Western blot analyses also showed greater expression of p16 and p21 in the old AECIIs (Fig. 2D, E).
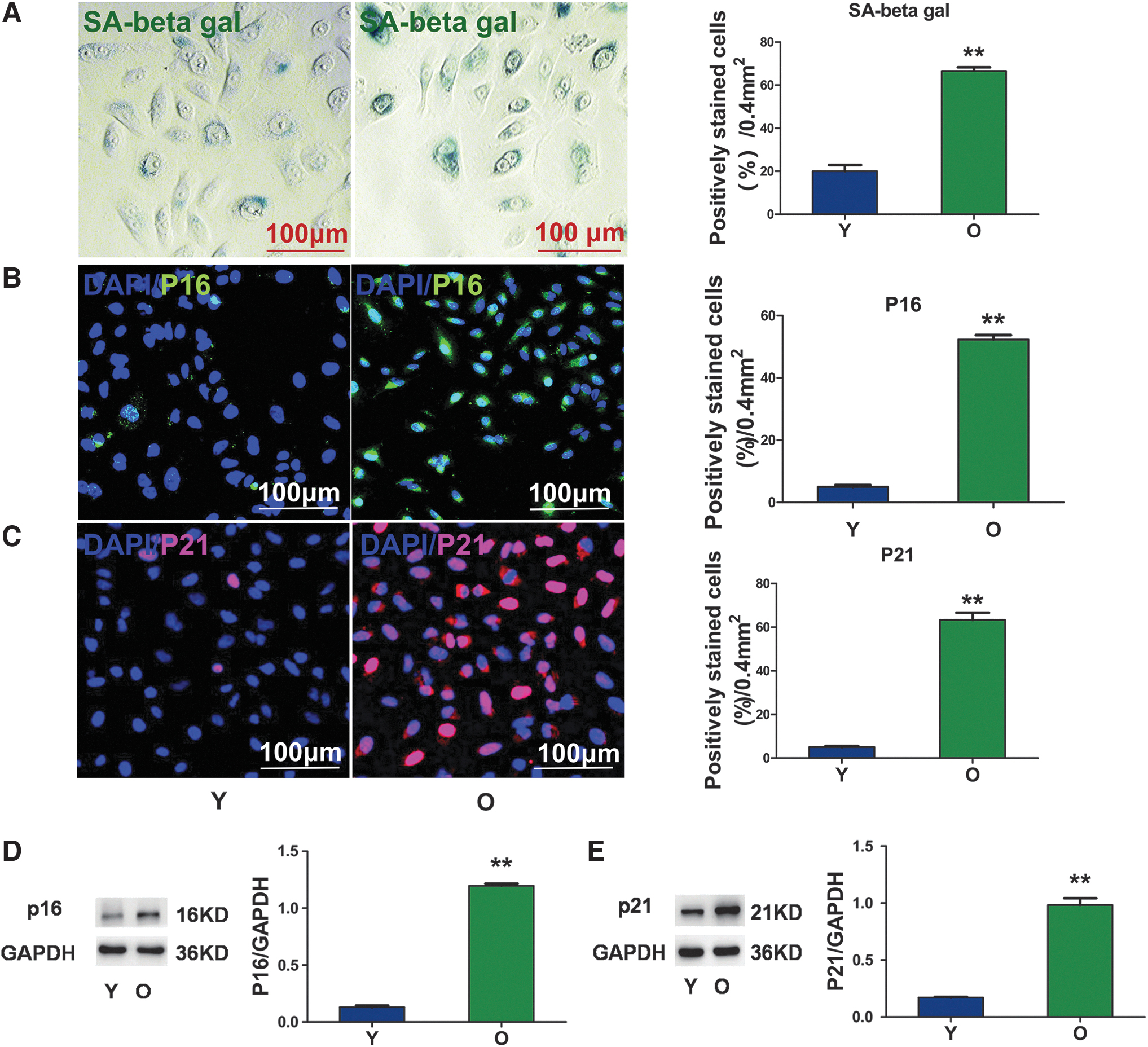
Changes in senescence indicators in young and senescent AECIIs.
Antiapoptotic activity of old AECIIs
The antiapoptotic activity induced by H2O2 (1 mM) was tested to examine the association between the SIRT3 level and oxidative stress (Fig. 3A). The cellular apoptotic rate of the old group was greater compared with the young group (Fig. 3B). The number of surviving cells in the old group was lower compared to that in the young group, as detected by CCK8 assay (Fig. 3C). mRNA and protein levels of SIRT3, MnSOD, and CAT were measured in young and old lung tissues and AECIIs using real-time PCR and western blotting. The expression levels of SIRT3 (Fig. 3D, E, D1, E1), MnSOD (Fig. 3F, G, F1, G1), and CAT (Fig. 3I, J, I1, J1) were significantly lower in old lung tissue and AECIIs compared to young lung tissue and AECIIs. The enzymatic activities of MnSOD and CAT (Fig. 3H, K, H1, K1) were also lower in old lung tissue and AECIIs.
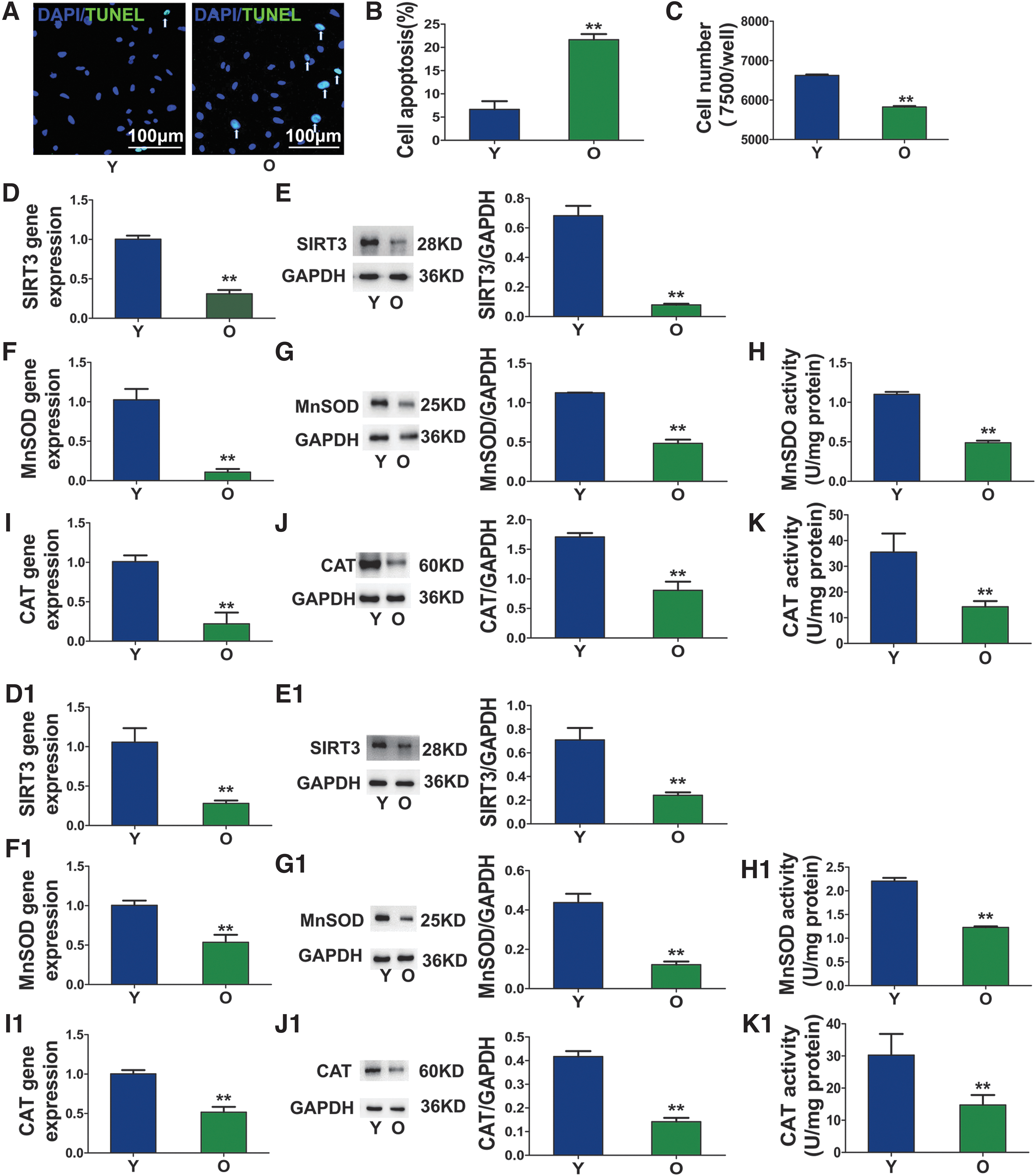
Antiapoptotic activity of senescent AECIIs.
SIRT3 upregulation improved the function of senescent AECIIs
Adenovirus (Ad)-SIRT3 was used to infect old AECIIs and to explore the effect of SIRT3 on cell function. Immunofluorescence staining suggested that the transduction efficiency was ∼65% (Fig. 4A). The antiapoptotic (Fig. 4B–D), proliferation (Fig. 4E), and secretion (Fig. 4F, G) activities of the Ad-SIRT3-infected old AECIIs were significantly improved compared to the uninfected control and Ad-GFP-infected groups, although a small difference was noticed between the uninfected group and the Ad-GFP-infected group.

Overexpression of SIRT3 improved the function of senescent AECIIs.
Changes in senescence indicators in old AECIIs after Ad-SIRT3 transduction
After transduction with Ad-SIRT3, the senescent status of AECIIs was dramatically reversed. The activity of SA-β-gal and immunofluorescence staining revealed the presence of fewer SA-β-gal-positive cells (Fig. 5A) and fewer p16- and p21-positive cells (Fig. 5B, C) in the Ad-SIRT3 group compared with the uninfected group or the Ad-GFP group. Significantly lower levels of p16 and p21 proteins were also detected in the Ad-SIRT3 group than in the untreated old group or the Ad-GFP group (Fig. 5D, E).
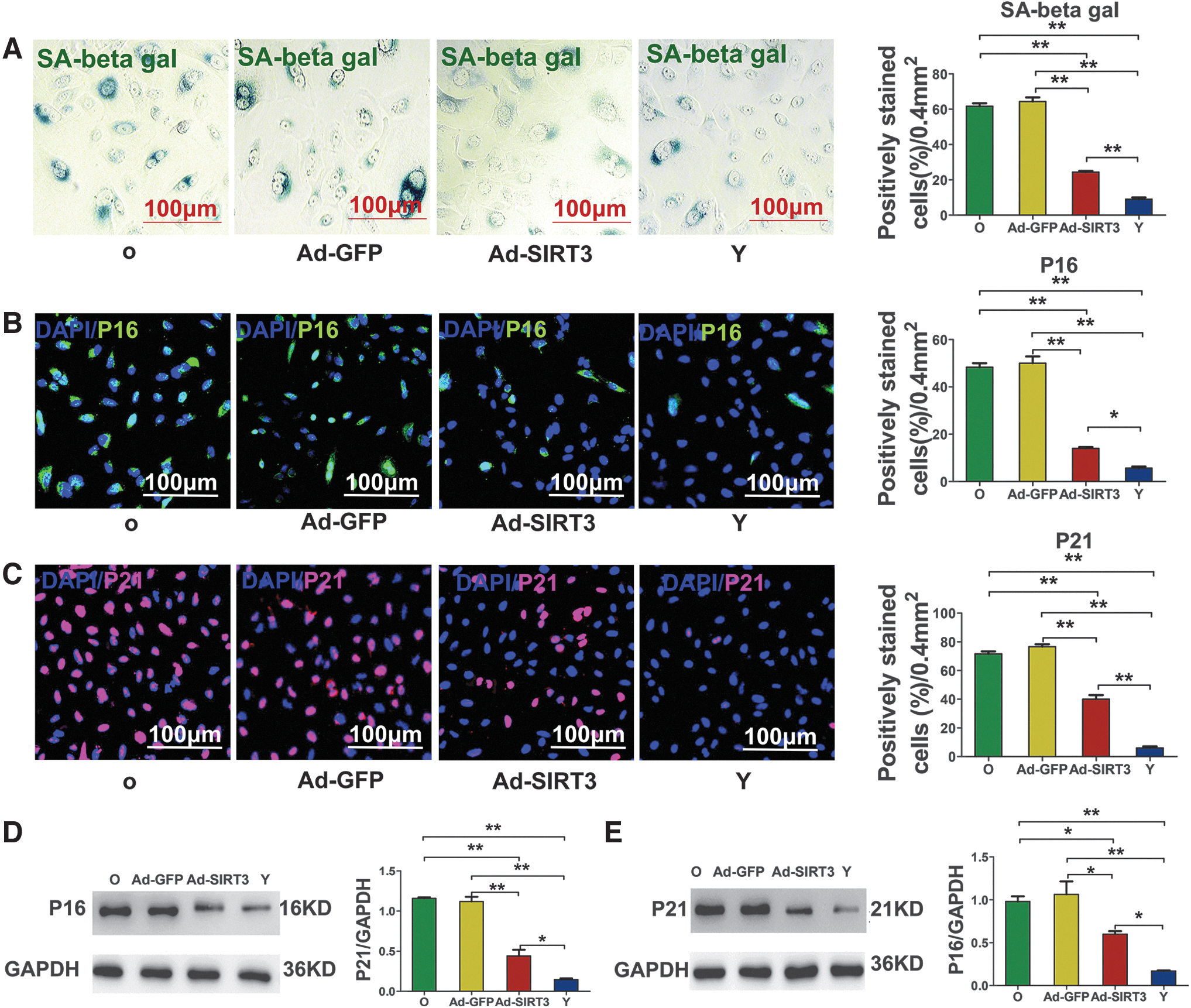
Changes in senescence indicators in senescent AECIIs after transfection.
Mechanism of action of SIRT3 in improving the cellular function of senescent AECIIs
After Ad-SIRT3 transduction, the expression of the FoxO3a protein was significantly upregulated in the nuclei following the upregulation of SIRT3 in the Ad-SIRT3 group (Fig. 6A–C). The mRNA and protein expression levels and the enzymatic activities of CAT and MnSOD in the Ad-SIRT3 group were higher than those in the uninfected old group and the Ad-GFP group (Fig. 6D–I).

The mechanism by which SIRT3 improves the functions of senescent AECIIs.
Discussion
Many diseases originate from aging, such as cardiovascular disease, Parkinson's disease, and lung diseases caused by the senescence of tissue stem cells and microenvironmental changes [26,27]. Aging is related to interactions between degenerative factors generated by senescent cells, such as free radicals, proteases, cytokines, and ROS [28]. Stem cell loss-of-function during the aging process is accompanied by the loss of the ability of the tissue or organ to regenerate. Accumulating evidence indicates that the accumulation of senescent cells in aging tissues may contribute to the development of age-related pathological changes, such as atherosclerosis, osteoarthritis, and lung disease.
The self-repair ability of senescent alveolar epithelial cells through either self-renewal or endogenous progenitor cells is lower than in younger cells, resulting in a smaller gas exchange surface area and reduced barrier function. These changes in senescent lung epithelial cells may promote the expansion of the mesenchymal compartment in the lung. Due to the limited number of therapeutic options to reverse or correct these losses, strategies designed to improve the function of senescent lung cells and delay the senescent process have become key factors for reducing the related mortality, which may represent a new paradigm in the treatment of parenchymal pulmonary diseases [29].
The integrity of the pulmonary alveolar epithelium and its dynamic interaction with the surrounding mesenchyme is crucial for efficient gas exchange and the function of host defenses, as the lung has a large epithelial surface area that is highly vascularized [9,10]. Each alveolus contains cuboidal AECIIs expressing high levels of SPC, as well as very thin AECIs that are in close apposition to capillaries [11,12]. AECIIs are the stem cells of lung epithelial cells. These cells repair the damaged alveolar epithelium and secrete SPC, which in turn reduces the surface tension during the breathing cycle and regulates inflammatory reactions [17].
In several pathological conditions, the delicate architecture of the alveoli is disrupted, resulting in severe respiratory diseases, such as the well-known chronic obstructive pulmonary disease (COPD) [30] and idiopathic pulmonary fibrosis [31]. These pathological processes are triggered, in part, by defects in the alveolar epithelium, such as SIRT3 protein deficiency. This deficiency increases the risk of pulmonary fibrosis and COPD by increasing mitochondrial oxidative DNA damage and apoptosis in the airway epithelium [32], along with mutations associated with abnormal surfactant protein processing.
Ideal or effective therapeutic options are currently unavailable for treating end-stage lung disease, except for lung transplantation, which is considered the most effective method for addressing terminal-stage lung disease. However, the shortage of donor organs and postoperative rejection limit the long-term survival of recipients. SIRT3 is categorized as a class III histone deacetylase and is a member of the highly conserved sirtuin family. It is the only member of the seven sirtuin proteins related to longevity and plays important roles in DNA repair, cell cycle progression, and life span prolongation. Two isoforms of human SIRT3 have been identified, fl-SIRT3 and sh-SIRT3. The short isoform of SIRT3 is considered to have higher functional significance.
As shown in our previous studies, SIRT3 protects senescent human mesenchymal stem cells from the damage caused by oxidative stress by upregulating antioxidant enzymes, including CAT and MnSOD [33,34]. This finding encouraged us to explore the relationship between sh-SIRT3 and lung aging, as well as the senescent status of AECIIs. Although SIRT3 protein has been linked to lung disease, the relative expression of SIRT3 in young and old healthy individuals had not been explored before our study. In this study, we clearly showed the differential expression of SIRT3 in young and old groups and highlighted the potential of SIRT3 to serve as a novel therapeutic option for treating lung diseases, which are characterized by progressive and irreversible damage to airway and alveolar tissues.
In this study, all the samples were obtained from patients with pneumothorax caused by pulmonary bullae, and the effects of pulmonary fibrosis, COPD, and other diseases on the experimental results were excluded to observe the changes in the function of AECIIs and the improvement in SIRT3 function under the state of natural aging.
The alveolar spaces were enlarged and mixed, and the alveolar septum was thinner in the old group than in the young group. In addition, the number and secretory activity of AECIIs in old lung tissues were dramatically lower compared with AECIIs from the young tissues, and the proliferation, secretion, and antiapoptotic activity of the old AECIIs were also dramatically lower compared with the young tissues. However, no differences in the differentiation time and ratio were observed between the two groups. Furthermore, p16, p21, and SA-β-gal staining were markedly increased in the old AECIIs compared to cells in the younger group. These results suggest that the function of AECIIs might be improved by ameliorating the senescent status of the lung tissue, thus potentially reducing the morbidity and mortality due to lung diseases.
In relation to the mechanism, we observed substantially lower expression levels of SIRT3, CAT, and MnSOD mRNA and protein in the old lung tissues and AECIIs compared to the young tissues and AECIIs. The enzymatic activities of CAT and MnSOD in the old AECIIs were also lower than those in their young counterparts.
Due to its low pathogenicity, high-efficiency gene transfer, and easy manipulation, we used an adenovirus vector to transfer the SIRT3 gene into AECII cells. Although SIRT3 expression in the Ad-SIRT3-infected old group did not reach the level of the young group, SIRT3 expression was dramatically upregulated and improved the antiapoptotic, proliferation, and secretory activities of these older tissue-derived cells. A slight reduction was noted in some parameters in the Ad-GFP-infected control group compared with the old group. The intrinsic immune response and local inflammatory reactions may be the main causes for these variations [25,35]. Moreover, the senescent status of the AECIIs was also markedly ameliorated.
Based on the results of our study, FoxO3a expression in the nucleus is closely associated with the expression and activities of SIRT3, CAT, and MnSOD. The upregulation of SIRT3 expression enhanced the antioxidant function through the FoxO3a-MnSOD-CAT pathway in AECIIs. Therefore, the upregulation of SIRT3 expression might be a new therapeutic target to treat respiratory diseases in the elderly population. In addition, we have studied and compared levels of these samples between male and female patients and found that sex had no significant effect on the results.
Due to constraints related to the experimental conditions, the study had some limitations. Although the specimens were obtained from patients with pneumothorax due to bulla rupture, a CT scan did not reveal obvious COPD or pulmonary fibrosis in these patients. However, the older patients may have had mild pulmonary fibrosis and a higher prevalence of COPD than the younger patients. Thus, detailed lung tissue histology and pulmonary function tests are required to exclude those possibilities. In our future study, we will investigate whether SIRT3 upregulation can improve lung function and the antioxidant activity of alveolar epithelial cells using in vivo models.
Conclusions
SIRT3 improves the function of senescent AECIIs by upregulating the expression of FoxO3a in the nucleus to positively regulate the enzymatic activities of the antioxidants MnSOD and CAT. This eventually improves the ability of AECIIs to resist oxidative stress and ameliorates the senescent state of these cells.
Footnotes
Author Disclosure Statement
The authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
This study was funded by the National Natural Science Foundation of China (Nos. 81770347 and 81270188) and the National Natural Science Foundation of China (Youth Fund Project) (No. 81500268).