
Abstract
Psoriasis is an autoimmune disease still lacking standard treatment, and it has been demonstrated that mesenchymal stem cells (MSCs) are capable of immunoregulation. The underlying mechanism might involve the secretion of soluble cytokines, such as hepatocyte growth factor (HGF). This study aims to investigate the therapeutic effect of HGF-overexpressed dental pulp stem cells (DPSCs) [DPSCs; HGF overexpressed DPSCs (HGF-DPSCs)] on imiquimod-induced psoriasis. DPSCs were isolated and transfected by adenovirus vector carrying HGF gene (Ad-HGF). The immunoregulatry abilities of DPSCs and HGF-DPSCs were investigated by coculture of the MSCs with peripheral blood mononuclear cells (PBMCs) under appropriated stimulation. The psoriatic mice were treated with saline control, DPSCs, or HGF-DPSCs. Then the mice spleens were collected and weighted. The psoriatic skin lesions were analyzed by Hematoxylin/Eosin and immunohistochemical staining for histopathological changes, and quantitative real-time polymerase chain reaction to detect the expression levels of CD4+ T cell-related transcription factors and cytokines. The mice blood serum was measured by MILLIPLEX analysis and enzyme-linked immunosorbent assay to evaluate the expression levels of inflammation cytokines. The coculture experiments showed HGF overexpression enhanced the immunoregulation abilities of DPSCs not by suppressing PBMCs' proliferation, but by downregulating T helper 1 (Th1), Th17 cells, and upregulating regulatory T (Treg) cells. In psoriatic skin lesions, the psoriasis-like erythema, scaling, and thickening were ameliorated; and the expression of cytokeratin 6 (CK6), and cytokeratin 17 (CK17) were downregulated by DPSCs and HGF-DPSCs treatment. HGF overexpression enhanced the decrease of spleen masses; enhanced the downregulation of the expression levels of interferon-gamma (IFN-γ), tumor necrosis factor-α, and interleukin (IL)-17A in the blood serums; enhanced the downregulation of T-box transcription factor 21 (T-bet), IFN-γ, retinoic acid-related orphan receptor-γt (RORγt), IL-17A, IL-17F, IL-23, and upregulation of Foxp3 and IL-10 in the psoriatic skin lesions. Therefore, HGF overexpression enhanced DPSCs' treatment effect on psoriasis mainly by reducing inflammatory responses. These findings might provide new immunoregulation strategies for psoriasis treatment.
Introduction
Psoriasis is an immune-mediated chronic skin disease, involving both local and systemic inflammation responses. The main pathological features of psoriasis are skin thickening, scales, erythema [1], and overexpression of inflammatory cytokines like interferon-gamma (IFN-γ), tumor necrosis factor-α (TNF-α), and interleukin 17 (IL-17) [2]. Psoriasis was initially regarded as a T helper 1 (Th1) cell-mediated skin disease [3]. Th1 cells and cytokines were overexpressed in psoriatic plaques and peripheral blood of psoriasis patients [4]. While following the identification of IL-17-secreting helper T (Th17) cells, numerous studies revealed that Th17 cells and their related cytokines like IL-17 and interleukin 23 (IL-23) played essential roles in psoriatic pathological processes as well [5]. In the last two decades, biologics, a kind of antibody medicine targeting the elevated inflammatory cytokines, was emerging as a new treatment for psoriasis [6]. However, severe adverse events had been reported in clinical trials using biologics, like severe infections (sepsis and tuberculosis), nonmelanoma skin cancers, multiple sclerosis, lupus, and congestive heart failure [7 –9]. The possible reason might be that the biologics neutralize all the targets they accessed, while the targeted cytokines participate not only in pathological changes of psoriasis, but also in other physiological processes, which might be affected by the neutralization [10,11]. Hence, new immunoregulation strategies for psoriasis treatment were needed.
It was demonstrated that mesenchymal stem cells (MSCs) possessed immunoregulation abilities [12]. MSCs could sense the status of the immune system and regulate it into a homeostatic state [13,14]. MSCs can restrain inflammation when the immune system is overactivated [15]. Some studies have revealed that human MSCs could treat psoriatic mice effectively by their immunoregulation abilities, such as human umbilical cord blood-derived MSCs, umbilical cord-derived MSCs, and embryonic stem cell-derived MSCs [16 –20]. What is more, it was demonstrated that human MSCs treatment reduced the severity of psoriasis in several clinical trials as well [21]. The major mechanism underlying the immunoregulation by MSCs involves the secretion of soluble factors, such as hepatocyte growth factor (HGF), indoleamine 2,3-dioxygenase (IDO), prostaglandin E2 (PGE2), and transforming growth factor beta 1 (TGF-β1) [15]. HGF is a kind of scatter factor, which is primarily secreted by mesenchymal cells [22]. It was found that HGF had therapeutic effects in various inflammatory diseases, including collagen-induced arthritis [23], graft-versus-host disease [24], and experimental autoimmune encephalomyelitis (EAE) [25].
Dental pulp stem cells (DPSCs) are a kind of MSCs like postnatal stem cells derived from ectoderm [26], which exhibit immunomodulatory abilities [27 –30]. Previously, the authors' group had found that overexpressing HGF in DPSCs can enhance the DPSCs' inhibition effect on TNF-α in osteoporotic mice [31]. Hence, we hypothesize that overexpressing HGF can promote the immunoregulation abilities of DPSCs, which may be beneficial to psoriasis treatment.
In this study, HGF-overexpressed DPSCs (HGF-DPSCs) were fabricated and whether their immunoregulation abilities were enhanced were tested. Then their treatment effect on psoriasis were investigated in a psoriasis mouse model.
Materials and Methods
Isolation, culture, and characterization of human DPSCs
Human DPSCs were isolated, cultured, and characterized as per previously described methods [32].
Isolation and culture: Dental pulp tissues were obtained from healthy human orthodontic molars of adult patients (19–29 years old) at Beijing Stomatological Hospital of Capital Medical University. Ethics approval was obtained from the Research Ethics Committee of Capital Medical University, China. All patients gave their written informed consent to participate in the study. The pulp tissues were cut into 1 mm3 and digested in a solution of 3 mg/mL collagenase type I enzyme (Worthington Biochem, Lakewood, NJ) and 4 mg/mL dispase-type αenzyme (Boehringer Mannheim, Germany) for 1 h at 37°C with vigorous shaking. Afterward, the digested tissue precipitations by centrifugation were seeded into culture flasks (NEST, Pacific Grove, CA) with α-MEM supplemented, 20% fetal bovine serum (Every Green, China), 100 U/mL of penicillin and streptomycin (Sigma, Ronkonkoma, NY), and then incubated in 5% CO2 incubator at 37°C. Upon reaching 80% confluence, hDPSCs were detached with 0.05% trypsin (Sigma), and passed with α-MEM supplemented with 10% fetal bovine serum (Every Green). The cells at passage 6 (P6) were used in this study, whereas extra cells were cryopreserved in liquid nitrogen for further use (−196°C).
Adipogenic differentiation: Human DPSCs (P6) were seeded in six-well plates (NEST) at a density of 1 × 105 cells/well. Then, cultured for 14 days with adipogenic induction medium, supplemented with 0.5 mM 3-isobuty-1-1-Methylxanthine, 1 μM dexamethasone, and 0.1 mM indomethacin (Sigma). Then, the cells were fixed in 4% (v/v) phosphate-buffered paraformaldehyde and stained with 0.5% Oil Red O (Sigma) solution. To examine the adipogenic differentiation, the stained cells were observed and photographed using a phase-contrast inverted microscope (Olympus, Japan).
Osteogenic differentiation: Human DPSCs (P6) were seeded in six-well plates (NEST) at a density of 1 × 105 cells/well. Next, cells were cultured for 21 days with an osteogenic induction medium, supplemented with 100 nM dexamethasone, 50 nM ascorbic acid, and 10 mM β-glycerophosphate (Sigma). Then the cells were fixed in 4% (v/v) phosphate-buffered paraformaldehyde and stained with 1% Alizarin Red solution. To examine the osteogenic differentiation, the stained cells were observed and photographed using a phase-contrast inverted microscope (Olympus).
Immunophenotype analysis: To characterize the immunophenotype of DPSCs, their expression of surface markers was analyzed by flow cytometry. Briefly, the tested cells were trypsinized and washed with phosphate-buffered saline (PBS). Around 2 × 105 cells/tube were harvested and incubated with various antibodies, including human CD73, CD90, CD105, CD45, CD19, CD34, CD11b, and HLA-DR in the dark for 30 min at room temperature; mouse IgG1 FITC and mouse IgG1 PE were used as isotype controls (BD Bioscience, Franklin Lakes, NJ). The analysis was performed with a FACSCalibur flow cytometer (BD Bioscience) and FlowJo software (FlowJo, Ashland).
Transfection of DPSCs by adenovirus vector
Adenovirus vector lack of exogenous genes (Ad-Null) and adenovirus vector carrying HGF gene (Ad-HGF) were used in this study, and both vectors were constructed as described previously [33]. The DPSCs were infected with 150 multiplicities of infection of Ad-Null or Ad-HGF. The Ad-Null transfected DPSCs (Null-DPSCs) and HGF-DPSCs were collected 48 h posttransfection for use. HGF expression was analyzed by quantitative real-time polymerase chain reaction (Q-PCR) and enzyme-linked immunosorbent assay (ELISA).
Peripheral blood mononuclear cell isolation
Human peripheral blood mononuclear cells (PBMCs) were isolated from the buffy coats of healthy volunteers by density gradient centrifugation using Ficoll-Paque (human lymphocyte separation medium, specific gravity 1.077, TBD sciences, China). The buffy coats were provided by the Beijing Red Cross Blood Center at Beijing. Briefly, the buffy coat samples were diluted with an equal volume of PBS, then slowly layered upon human lymphocyte separation medium, and centrifuged at 600 g for 20 min. PBMCs were collected from the interphase and washed with PBS supplemented with 5% fetal bovine serum (Every Green). The isolated PBMCs were cultured in RPMI-1640 medium (Gibco, Germany) supplemented with 10% fetal bovine serum (Every Green), 100 U/mL of penicillin, and streptomycin (Sigma).
MSCs/PBMCs cocultures
To investigate the immunomodulatory abilities of DPSCs and HGF-DPSCs, DPSCs or HGF-DPSCs were cocultured with PBMCs under appropriate stimulation, then the CD4+ T cell differentiation assay and PBMC proliferation assay were conducted.
CD4+ T cell differentiation assay: Briefly, 2 × 105 DPSCs or HGF-DPSCs were plated in six-well plates (NEST), and treated with 25 μg/mL Mitomycin C (Selleck Chemicals, Houston, TX) for 30 min at 37°C, then washed with RPMI-1640 medium (Gibco) twice. PBMCs were suspended in RPMI-1640 medium supplemented with 10% fetal bovine serum (Every Green), 15 μL/mL CD3/CD28 Fab-Strep (IBA Lifesciences, Germany), 200 U/mL interleukin 2 (IL-2; Roche, United States), 100 U/mL penicillin and streptomycin (Sigma) at a concentration of 1 × 105 cells/mL. The PBMCs were added to six-well plates with or without DPSCs or HGF-DPSCs at a ratio of 1:1, then incubated for 5 days.
The supernatants were collected to measure the production of CD4+ T cell-related cytokines, including IFN-γ, Interleukin-4 (IL-4), TGF-β1, and IL-6 by ELISA. The PBMCs were harvested, and the percentages of CD4+IFN-γ+ Th1 cells, CD4+IL-4+ Th2 cells, CD4+IL-17A+ Th17 cells, and CD4+CD25+CD127− regulatory T (Treg) cells were analyzed by flow cytometry. To detect Th1, Th2, and Th17 cells, the PBMCs were stimulated for 4 h with a leukocyte activation cocktail in the presence of GolgiStop (BD Bioscience). Then, the PBMCs were incubated with BB700-conjugated anti-CD4 antibody (BD Bioscience) for 30 min at 4°C in the dark. Afterward, the cells were fixed and permeabilized, and stained with FITC-conjugated anti-IFN-γ antibody, APC-conjugated anti-IL-4 antibody, and PE-conjugated anti-IL-17A antibody (BD Bioscience) for 30 min in the dark at 4°C. To detect Treg cells, the PBMCs were stained with BB700-conjugated anti-CD4 antibody, BB515-conjugated anti-CD25 antibody, and Alexa Fluor 647-conjugated anti-CD127 (BD Bioscience) for 30 min in the dark at 4°C. The stained cells were analyzed by FACSCalibur flow cytometer (BD Bioscience) and FlowJo software.
PBMCs proliferation assay: Briefly, 2 × 105 DPSCs or HGF-DPSCs were plated in six-well plates (NEST), and treated with 25 μg/mL Mitomycin C (Selleck Chemicals) for 30 min at 37°C, then washed with RPMI-1640 medium (Gibco) twice. PBMCs were labeled with 5 μM Dye 670 (Thermo Fisher Scientific, Waltham, MA) for 10 min at 37°C, and suspended in RPMI-1640 medium supplemented with 10% fetal bovine serum (Every Green), 10 μg/mL PHA (Sigma), 100 U/mL of penicillin and streptomycin (Sigma) at a concentration of 1 × 105 cells/mL. The PBMCs were added to six-well plates with or without DPSCs or HGF-DPSCs at a ratio of 1:1, then incubated for 3 days. The proliferation index of PBMCs was analyzed by FACSCalibur flow cytometer (BD Bioscience) and ModFit software (Verity Software House, Topsham, ME).
Mice and treatment
Twenty female BALB/C mice (18–22 g, 8 weeks old) were used in this study. All animal experiments were approved by the Ethics Committee of the Beijing Institute of Radiation Medicine. The mice were held in a specific pathogen-free animal laboratory house, and all procedures were carried out in accordance with the relevant guidelines and regulations.
The back skin of all mice was shaved, then the mice were divided into four groups. The Control group, DPSC group, and HGF-DPSC group received a daily topical dose of 62.5 mg of imiquimod (IMQ) cream (IMQ, 5%, Med Shine Pharmaceutical, China) on the shaved back for 6 days to induce psoriasis-like symptoms as previously described [34]. The Blank group did not receive IMQ. Twenty-four hours before IMQ application, all mice received an intravenous tail injection of 200 μL normal saline (Shijiazhuang No.4 pharmaceutical, China) suspension, respectively. The Blank group and Control group received only normal saline. The DPSC group received normal saline suspension of 2 × 106 DPSCs. The HGF-DPSC group received normal saline suspension of 2 × 106 HGF-DPSCs. The mice were euthanized and sacrificed on day 6. The blood serums, psoriasis-like back skin lesions, and spleens were collected for further analysis.
The determination of the spleen index
Mice were weighed before sacrifice, then their spleens were excised and weighted immediately after sacrifice. The spleen indexes were calculated according to the following formula:
Spleen index (mg/g) = spleen weight (mg)/body weight (g)
Histological and immunohistochemical analysis of the back skins
Sections from the mice back skins were stained with Hematoxylin/Eosin (H&E) stain for histological evaluation. Briefly, the skin samples were fixed and embedded in paraffin. Tissue sections of 5 μm thickness were deparaffinized with xylene and rehydration with a graded ethanol solution, then stained with H&E. All sections were observed and photographed under a phase-contrast inverted microscope (Olympus), and the epidermal thickness of psoriasis-like back skin lesions were measured by ImageJ software (National Institutes of Health, Bethesda, MD).
The skin sections as above were analyzed by immunohistochemical stains. Briefly, the sections were incubated with PBS or the following primary antibodies at 4°C overnight: rabbit anti-mouse CD8, CD83, Cytokeratin 6 (CK6), and Cytokeratin 17 (CK17) (Bioss, China). The biotinylated goat anti-rabbit secondary antibodies were conjugated to streptavidin-peroxidase, and DAB solution was used to observe the immunoreactivity (Bioss, China). All the sections were observed and photographed using a light microscope (Olympus).
ELISA analysis
The concentrations of HGF in the culture supernatants of DPSCs, Null-DPSCs, and HGF-DPSCs; the concentrations of IFN-γ, IL-4, TGF-β1, and IL-6 in the coculture supernatants of PBMCs/DPSCs and PBMCs/HGF-DPSCs; and the concentrations of HGF and TGF-β in the mice blood serums were quantitated by the anti-human ELISA Kits according to the manufacturer's instructions (Neobioscience Biotech, China). Briefly, all samples were centrifuged at 400 g for 10 min, then diluted or stimulated, respectively. The pretreated samples were added to the test wells and incubated for 90 min at 37°C to capture the antibodies. Afterward, the test wells were incubated with biotinylated antibodies and avidin HRP successively. The color was developed and stopped using tetramethylbenzidine substrate solution and stop solution. The plates were read by a microplate manager (Bio-Rad, Hercules, CA) at 450 nm.
MILLIPLEX analysis
The concentrations of inflammatory cytokines IFN-γ, TNF-α, IL-17A, and IL-6 in the mice blood serums were measured by using the MILLIPLEX Kit, according to the manufacturer's instructions (Mouse Th17 Magnetic Bead Panel; Merck Millipore, Burlington, MA). Briefly, the background, standard, or sample were mixed with chemically dyed antibody-bound beads in the plate wells, incubated with agitation overnight at 4°C, and then incubated with biotinylated detection antibodies for 1 h and Streptavidin/Phycoerythrin reporter molecules for 30 min at room temperature. Data were collected by MAGPI and analyzed with MILLIPLEX Analyst software (Merck Millipore).
Q-PCR analysis
The gene expression of HGF in DPSCs, Null-DPSCs, and HGF-DPSCs; and the gene expression of T-box transcription factor 21 (T-bet), IFN-γ, GATA-binding protein 3 (GATA3), IL-4, Retinoic acid-related orphan receptor-γt (RORγt), IL-17A, Interleukin 17F (IL-17F), IL-23, Forkhead box protein 3 (FOXP3), and Interleukin 10 (IL-10) in the mice skin lesions were measured by Q-PCR analysis. Briefly, total ribose nucleic acid (RNA) from all samples was extracted using TRIzol Reagent (Invitrogen, Waltham, MA), according to the manufacturer's instructions. First-strand complementary deoxyribonucleic acid (cDNA) was synthesized, and the quantification of messenger RNA (mRNA) expression was performed on ABI PRISM 7500 FAST sequence detector (Applied Biosystems). The expression of the above genes was evaluated by Q-PCR using the human primers listed in Table 1 and mouse primers listed in Table 2, with GAPDH as internal control, respectively. The relative gene expression levels were calculated by the 2−ΔΔCT method.
The Human Primers for Quantitative Real-Time Polymerase Chain Reaction
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HGF, hepatocyte growth factor.
The Mouse Primers for Quantitative Real-Time Polymerase Chain Reaction
GATA3, GATA-binding protein 3; IFN-γ, interferon-gamma; IL, interleukin; FOXP3, Forkhead box protein 3; RORγt, retinoic acid-related orphan receptor-γt; T-bet, T-box transcription factor 21.
Statistical analysis
Data were graphed and statistically analyzed by using GraphPad Prism 6.0 (GraphPad Software, San Diego, CA). All data were expressed as the mean ± SD. The unpaired t-test was used to compare the mean of two independent groups, and the level of significance was set as P < 0.05.
Results
Characterization of DPSCs
DPSCs were obtained from human dental pulp. Spindle-like cells were observed around the digested pulp tissue on day 10 (Fig. 1A). The cells were digested with trypsin and passaged. At P6, the cells were spirally arranged and had the same typical spindle shape as MSCs (Fig. 1B). After 14 days of adipogenic induction and Oil Red stain, lipid droplets were clearly observed at the internal side of the cells (Fig. 1C). After 21 days of osteogenic induction, mineralized nodules were formed and verified by Alizarin Red staining (Fig. 1D). The results of the flow cytometric analysis demonstrated that the DPSCs positively expressed CD73, CD90, and CD105, and negatively expressed CD19, CD45, CD34, CD11b, and HLA-DR (Fig. 1E). These results indicated that the obtained cells were MSCs.
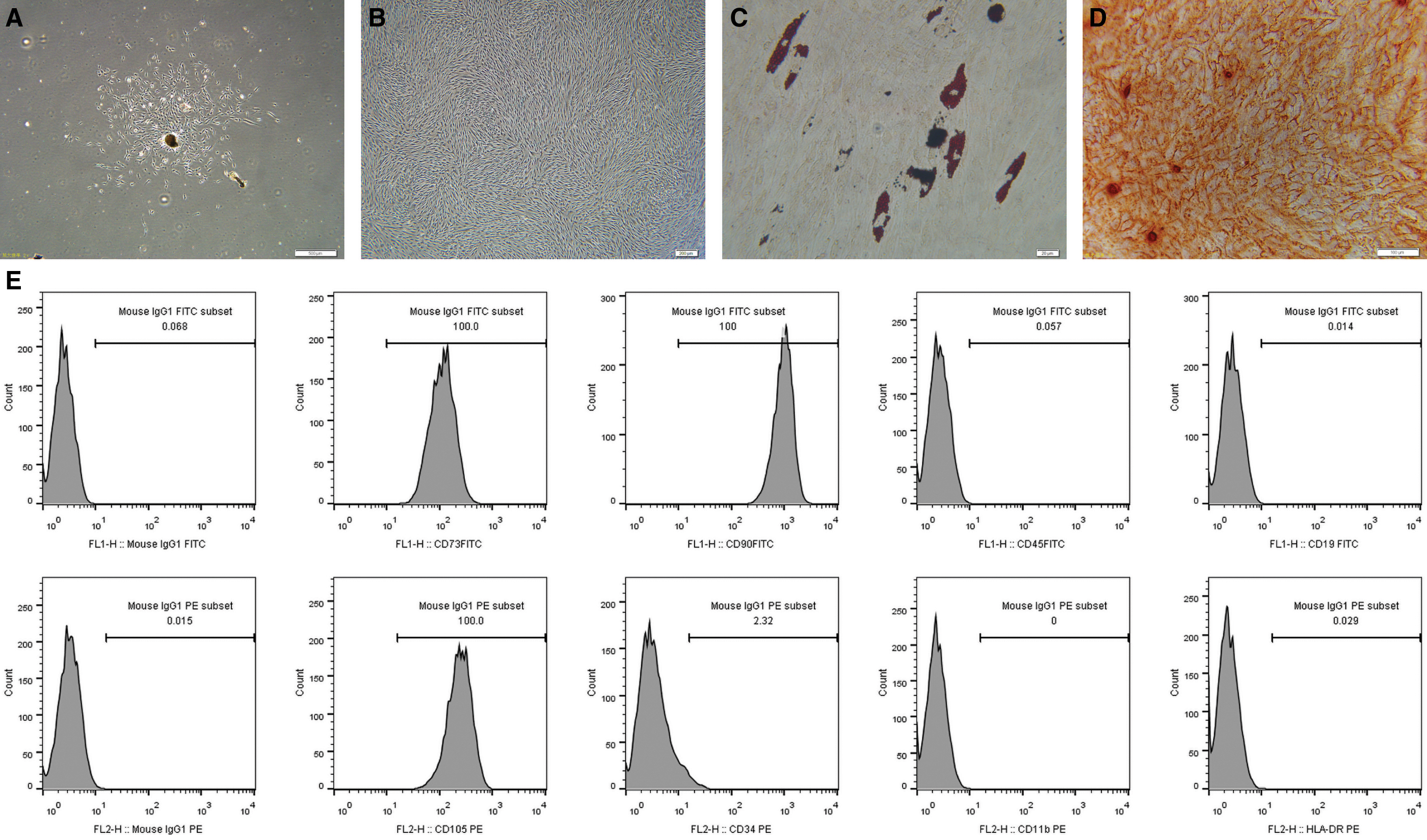
Isolation and characterization of DPSCs.
HGF overexpression enhanced the immunomodulatory abilities of DPSCs
To evaluate the transduction efficiency, the mRNA and protein expression levels of HGF in DPSCs, Null-DPSCs, and HGF-DPSCs were tested by Q-PCR analysis or ELISA analysis. The results showed that the HGF gene expression was significantly higher in HGF-DPSCs than in DPSCs (P < 0.05) and Null-DPSCs (P < 0.05), and there is no difference between the DPSC group and Null-DPSC group (P > 0.05, Fig. 2A). The concentration of HGF was 1,475 ± 326.1 pg/mL in DPSC culture supernatants, 1,420 ± 18.78 pg/mL in Null-DPSC culture supernatants, and 18,611 ± 187.9 pg/mL in HGF-DPSC culture supernatants (Fig. 2B), indicating that HGF was successfully overexpressed by Ad-HGF transfection in HGF-DPSCs.
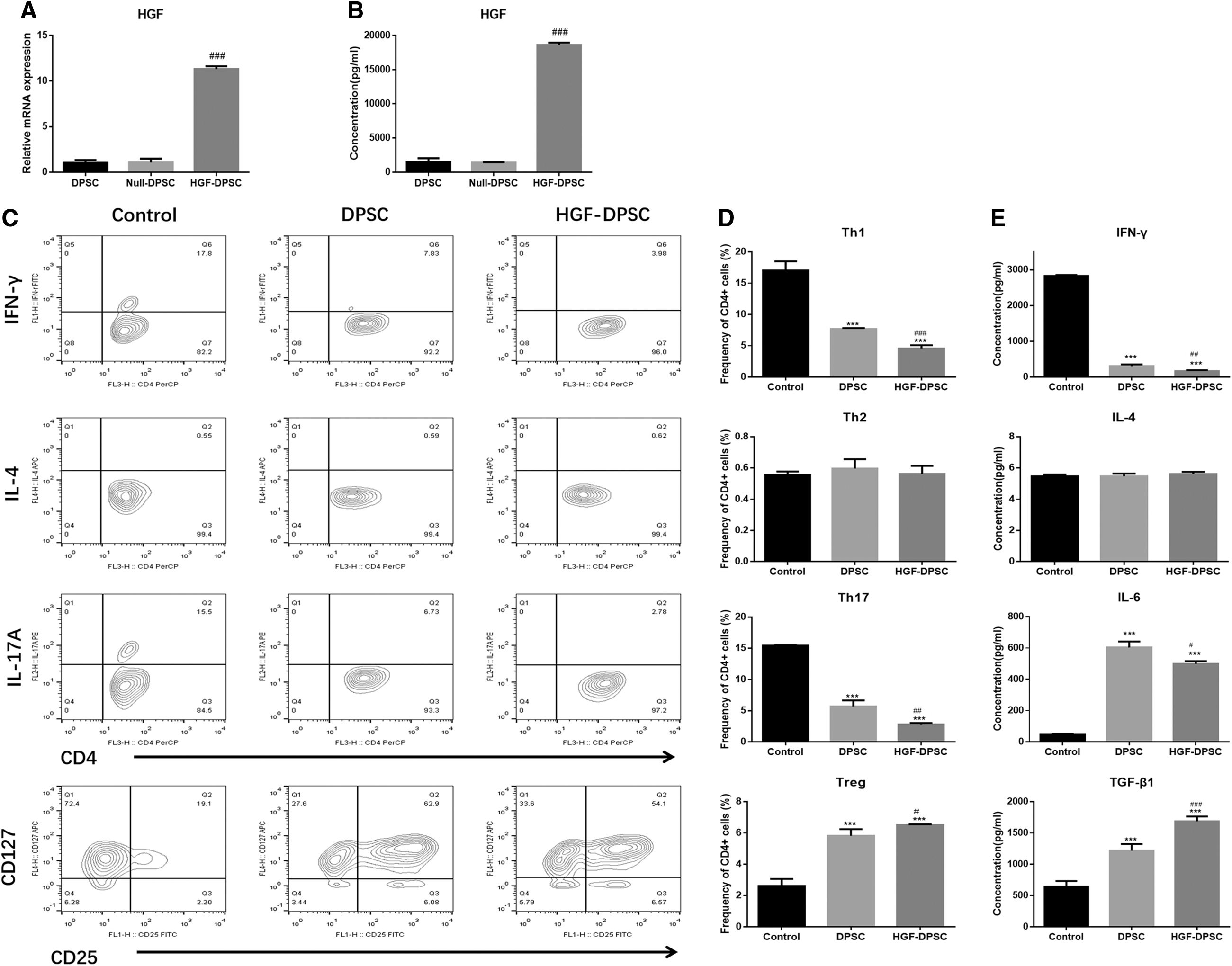
Characterization of HGF-DPSCs. DPSCs were transfected by Ad-Null or Ad-HGF, 48 h later,
To investigate the effect of HGF overexpression on the immunomodulatory abilities of DPSCs, DPSCs or HGF-DPSCs were cocultured with PBMCs under appropriate stimulation. The results of CD4+ T cell differentiation assay showed that the percentages of Th1 and Th17 cells were significantly lower, and the percentage of Treg cells were significantly higher in the DPSC group and HGF-DPSC group compared with the Control group (P < 0.05) (Fig. 2C, D). Moreover, in the HGF-DPSC group, the percentages of Th1 (4.57 ± 0.31, P < 0.05) and Th17 (2.83 ± 0.12, P < 0.05) cells were significantly decreased, and the percentage of Treg (6.51 ± 0.03, P < 0.05) cells were significantly increased compared with the DPSC group. However, no difference was found in the percentages of Th2 cells among these three groups (Fig. 2C, D). These results suggested that HGF overexpression could enhance the immunomodulatory abilities of DPSCs by suppressing Th1, Th17 cells' activities, and promoting Treg cells' activity. The ELISA results confirmed that compared with DPSC group, the concentrations of Th1 cell dedifferentiation-related cytokine IFN-γ, Th17 cell dedifferentiation-related cytokine IL-6 were downregulated, and the concentration of Treg cell dedifferentiation-related cytokine TGF-β was upregulated in the HGF-DPSC group. In addition, there was no difference in the concentration of Th2 cell dedifferentiation-related cytokine IL-4 among these three groups (Fig. 2E).
DPSCs and HGF-DPSCs reduced the severity of the pathological changes in psoriatic mice
To investigate the effect of DPSCs and HGF-DPSCs on psoriasis treatment, a psoriatic mouse model was established and treated with DPSCs or HGF-DPSCs by intravenously injection (Fig. 3A). Phenotypically, psoriasis-like erythema, scaling and thickening were observed on the back skin of the Control mice induced by IMQ application. Compared with the Control group, the psoriatic symptoms (especially scaling) were reduced in both DPSC group and HGF-DPSC group on day 6 (Fig. 3B). H&E staining results showed that compared with the Blank group, IMQ application induced psoriasis-like pathological changes in the back skin of the control mice, such as inflammatory infiltration, thickened epidermally, parakeratosis, and hyperkeratosis. Moreover, both DPSC and HGF-DPSC treatment ameliorated these psoriatic-like pathological changes. Compared with the DPSC group, the inflammatory infiltration and thickened epidermis were more alleviated in the HGF-DPSC group (Fig. 3C). As shown in the calculation results, the epidermal thickness was lower in the DPSC group and HGF-DPSC group compared with the Control group and more reduced in the HGF-DPSC group (Fig. 3D). To further investigate the pathological changes, the expression of inflammatory infiltration-related cytokines CD8 and CD83, and keratinocyte differentiation-related cytokines CK6 and CK17 in the psoriatic skin lesions were tested. As shown in the immunohistochemical results, the expression of CD8, CD83, CK6, and CK17 were upregulated in the Control group compared with the Blank group. While compared with the Control group, the expression of CD8, CD83, CK6, CK17 were downregulated in the DPSC group and HGF-DPSC group (Fig. 4).

The treatment effects of DPSCs and HGF-DPSCs on IMQ-induced psoriasis in mice.
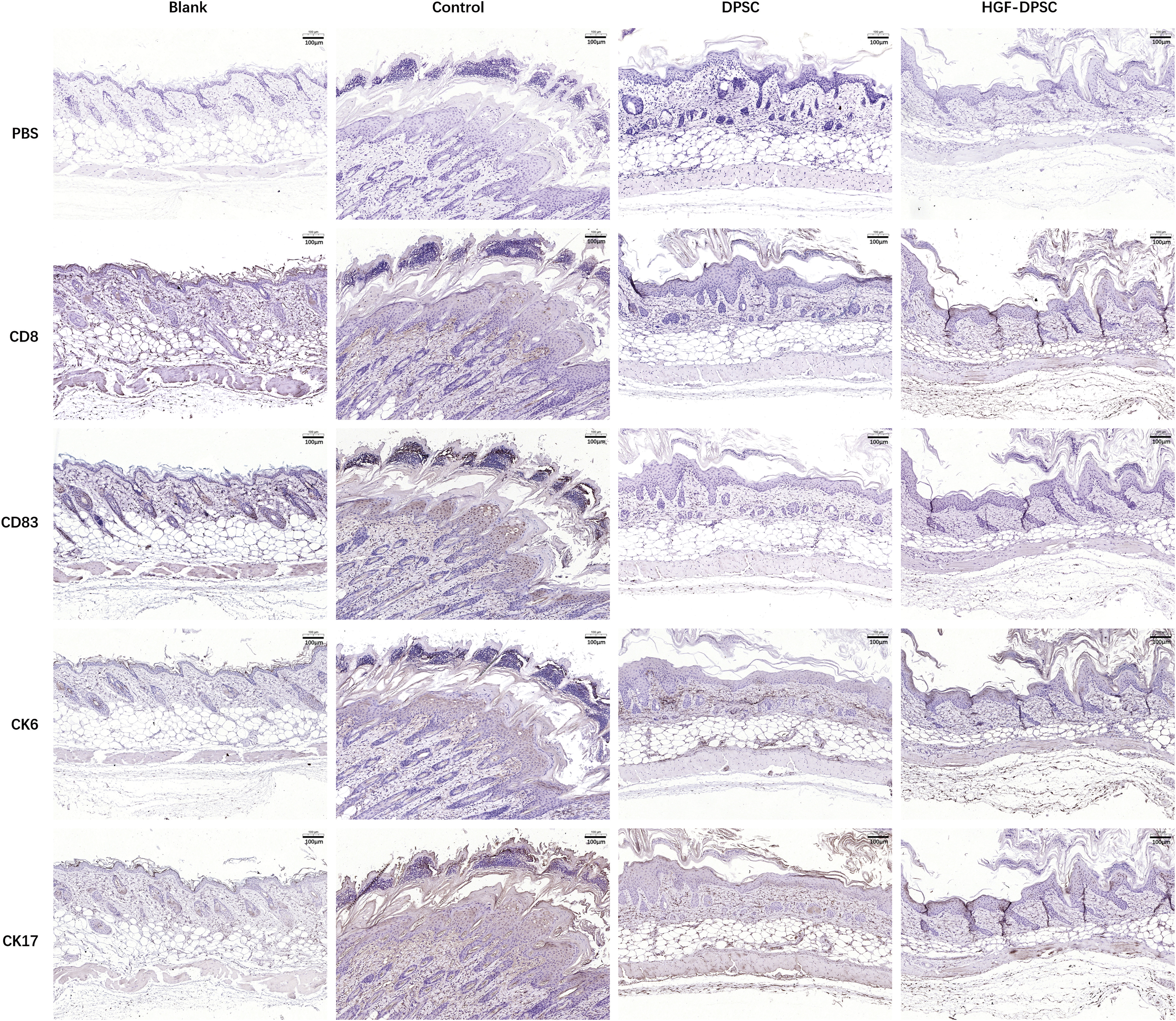
Immunohistochemical examination of the psoriatic back skin lesions. The skin sections were analyzed by immunohistochemical staining with anti-mouse CD8, CD83, CK6, and CK17. CK, cytokeratin. Color images are available online.
HGF overexpression enhanced the treatment effect of DPSCs on psoriasis by reducing inflammatory responses
The ELISA results showed that the concentration of HGF in the mice blood serum was higher in the DPSC group and HGF-DPSC group compared with the Control group, and it was significantly higher in the HGF-DPSC group (1,031 ± 20.80 pg/mL) than in the DPSC group (628 ± 39.39 pg/mL, P < 0.05, Fig. 3C), indicating that HGF was overexpressed by HGF-DPSC transplantation in the treated mice (Fig. 5A). Spleens are important immune organs, and at the end of the animal experiments, the mice spleens were enlarged in the Control group, whereas the spleen masses were decreased in both DPSC group and HGF-DPSC group compared with the Control group. Moreover, the spleen masses were significantly lower in the HGF-DPSC group (10.14 ± 0.18) than in the DPSC group (11.68 ± 0.42, P < 0.05, Fig. 5B, C). It was demonstrated that the serum levels of IFN-γ, TNF-α, and IL-17A are significantly increased in psoriatic patients [2]. In this study, as shown by the MILLIPLEX analysis, the serum expression levels of IFN-γ, TNF-α, and IL-17A were lower in both DPSC group and HGF-DPSC group compared with the Control group, with a significantly lower rate in HGF-DPSC group than in the DPSC group (P < 0.05, Fig. 5D).
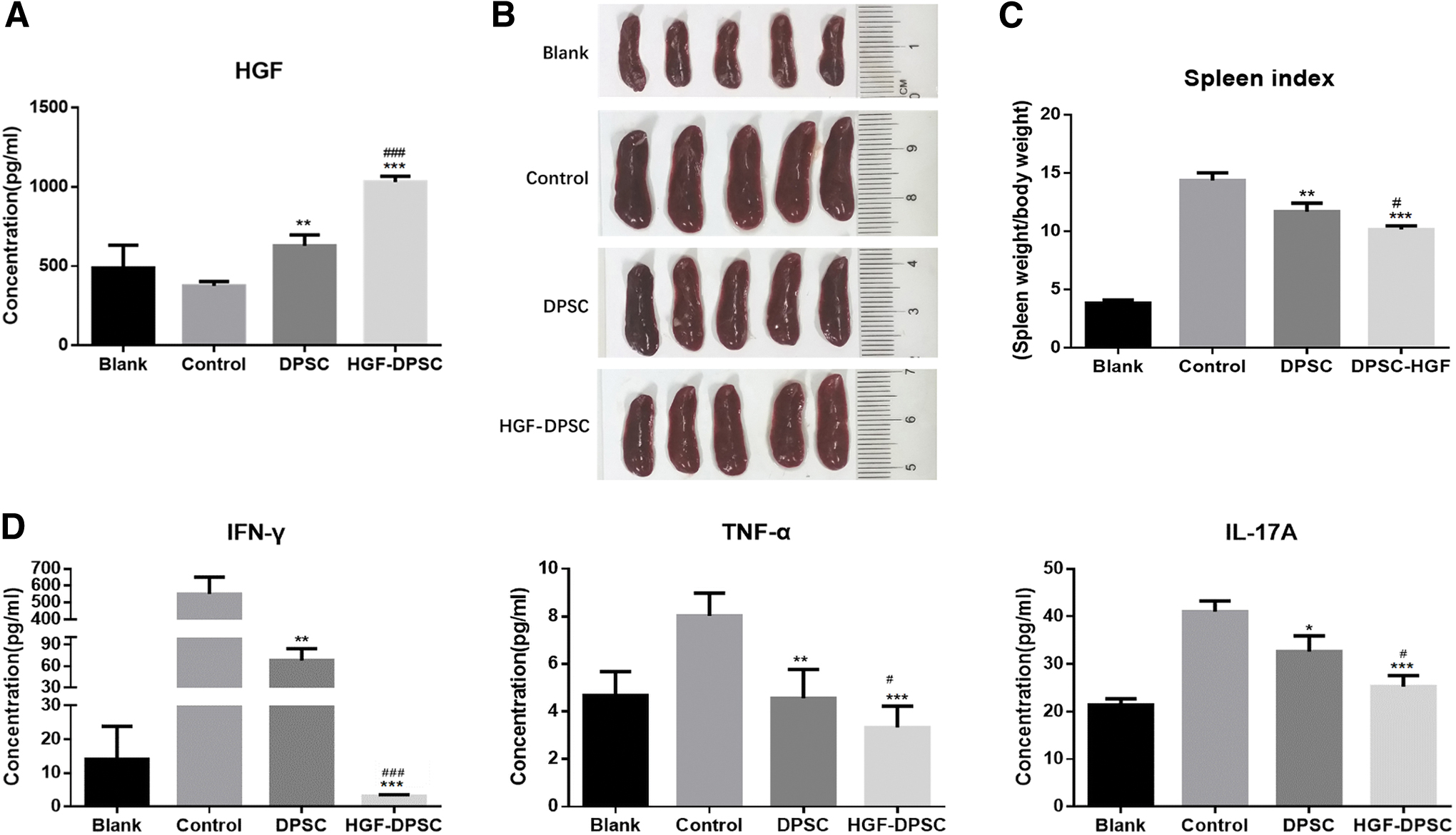
Effects of DPSCs and HGF-DPSCs on psoriatic inflammation in mice.
To further investigate the suppression effect of HGF overexpression on psoriatic inflammation, the expression levels of CD4+ T cell-related key transcription factors and cytokines in the mice back skin lesions were analyzed by Q-PCR. The results showed that compared with the Blank group, IMQ application upregulated the expression levels of T-bet (Th1 transcription factor), IFN-γ (Th1-related cytokine), RORγt (Th17 transcription factor), IL-17A, IL-17F, and IL-23 (Th17-related cytokines); downregulated the expression levels of GATA3 (Th2 transcription factor), IL-4 (Th2-related cytokine), Foxp3 (Treg transcription factor) and IL-10 (Treg-related cytokine) in the Control group. While compared with the Control group, DPSC and HGF-DPSC treatment significantly downregulated the expression levels of T-bet, IFN-γ, RORγt, IL-17A, IL-17F, and IL-23, and upregulated the expression levels of GATA3, Foxp3, and IL-10. Moreover, compared with the DPSC group, HGF overexpression enhanced the downregulation of T-bet, IFN-γ, RORγt, IL-17A, IL-17F, and IL-23, and upregulation of the Foxp3 and IL-10 in the HGF-DPSC group (Fig. 6).
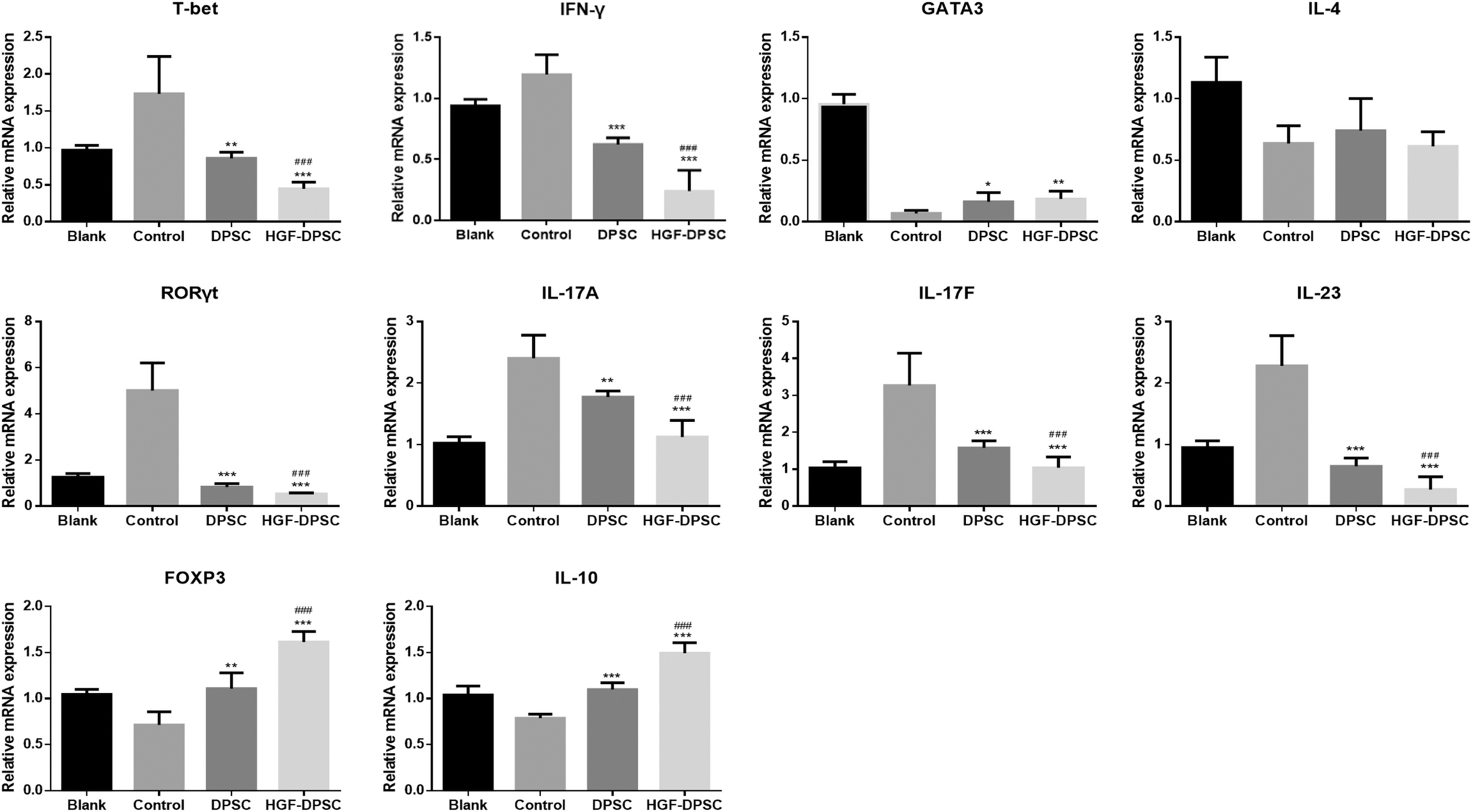
The expression levels of CD4+ T helper cell-related transcription factors and cytokines in the skin lesions of all mice. The expression levels of T-bet, IFN-γ, GATA3, IL-4, RORγt, IL-17A, IL-17F, IL-23, FOXP3, and IL-10 in the psoriatic skin lesions of all mice were tested by Q-PCR and calculated by the 2−ΔΔCT method. * means P < 0.05, compared with control group; ** means P < 0.01, compared with control group; *** means P < 0.001, compared with Control group; ### means P < 0.001, compared with DPSC group. FOXP3, Forkhead box protein 3; GATA3, GATA-binding protein 3; RORγ, retinoic acid-related orphan receptor-γt.
Discussion
Psoriasis is a Th1 and Th17 cell overacting-related chronic inflammation disease. In this study, we first demonstrated that DPSCs could alleviate the severity of psoriasis, and HGF overexpressing could enhance their treatment effect by reducing psoriatic inflammatory conditions. It was reported that MSCs could reduce the psoriatic inflammation by rebalancing the Th1 and Th17 responses in psoriatic mice [17,18,20]. In this study, except for suppressing Th1 and Th17 cells-related transcription factors and cytokines, DPSC treatment promoted the Treg transcription factor Foxp3 and Treg-related cytokine IL-10 in the psoriatic skin lesions (Fig. 6), indicating that DPSCs reduced the severity of psoriasis not only by downregulating Th1 and Th17 cells' activities but also by upregulating Treg cells' activities. Treg cells are a kind of immunoregulatory cells, which are capable of suppressing autoimmunity [35]. It has been demonstrated that a same precursor cell was shared by Th17 cells and Treg cells [36]. Additionally, its differentiation was regulated by IL-6 and TGF-β in a concentration-dependent manner; low concentrations of TGF-β synergize with IL-6 to promote the activation of RORγt and Th17 differentiation, whereas high concentrations of TGF-β induce Foxp3 expression and Treg differentiation [37]. In this study both the in vitro and in vivo experiments showed that high concentrations of TGF-β was induced by DPSC treatment (Fig. 2E and Supplementary Figure S1). Therefore, other than suppressing Th1 and Th17 responses, the underlying mechanism of the treatment effect of DPSCs on psoriasis might include promoting TGF-β secretion and Treg cells' differentiation.
HGF is primarily secreted by mesenchymal cells and is useful for attenuating inflammatory response [38,39]. It was reported that MSCs' conditioned medium reduced proinflammatory cytokine expression in a model of EAE, which could be blocked by HGF neutralization [40]. This indicated that HGF overexpression may enhance the immunoregulation abilities of MSC. However, an in vitro experiment showed that little additive impact was exhibited by HGF overexpression on the suppression of T lymphocyte proliferation [40]. In this study, we evaluated the effect of HGF overexpression on PBMCs' differentiation and proliferation. The results showed that HGF overexpression enhanced the suppression effect of DPSCs on the activities of Th1 and Th17 cells, and enhanced the promotion effect of DPSCs on the activities Treg cells (Fig. 2). While HGF overexpression did not change the suppression effect of DPSCs on the proliferation of PBMCs (Supplementary Fig. S2). These results indicated that HGF overexpression enhanced the immunoregulation abilities of DPSCs mainly by promoting their rebalancing effect on PBMCs' differentiation not proliferation. The HGF overexpression significantly downregulated the expression levels of T-bet, IFN-γ, RORγt, IL-17A, IL-17F, IL-23, and upregulated the expression levels of Foxp3, IL-10 in mice psoriatic skin lesions (Fig. 6), suggesting that HGF overexpression reduced psoriatic inflammation by suppressing Th1 and Th17 activities and promoting Treg activities.
TGF-β is a kind of cytokine that plays multiple roles in wound healing, fibrogenesis, and immunoregulation [41]. It was reported that HGF can act synergistically or antagonistically with TGF-β [42]. During fibrosis, HGF overexpression frequently downregulates TGF-β to alleviate fibrotic degree by reducing the excessive deposition of collagen and extracellular matrix [43 –45]. However, in organ transplantation conditions, HGF treatment usually elevates TGF-β expression to prolong the transplants' survival by decreasing allograft rejection [46 –48]. In this study, the expression of TGF-β was upregulated by HGF overexpression in the psoriatic mice model (Supplementary Fig. S1), suggesting that in autoimmune conditions, HGF may act synergistically with TGF-β to reduce the severity of inflammation.
This study still has several limitations. First, the psoriasis mouse model adopted in this study was induced by IMQ. It is a prevalent model. However, the skin inflammation decreases during the seventh or eighth day when IMQ is applied to the mice. Therefore, a longtime treatment effect cannot be evaluated in this study. Second, further investigations are needed to confirm the role of TGF-β in the psoriasis severity, as well as the correlations between HGF overexpression and TGF-β under inflammation conditions.
Conclusions
Our study demonstrated that the administration of DPSCs or HGF-DPSCs ameliorated the severity of psoriasis, and HGF overexpression enhanced the treatment effect of DPSCs on psoriasis mainly by reducing inflammatory responses.
Footnotes
Acknowledgment
The buffy coats of healthy donors were obtained through the Beijing Red Cross Blood Center from Yanjun Jia. The MILLIPLEX analysis was performed in Neobioscience Biotech. The authors thank all of them for helping.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.