
Abstract
Diabetes is a complex multifactorial disorder associated with hyperglycemia, oxidative stress, and inflammation. The pathological microenvironment impairs mesenchymal stem cell (MSC) viability and dysregulates their proregenerative and immune-modulatory function causing maladaptive tissue damage. Targeting stem cells to protect them against impairment could thus delay the onset of complications and enhance the quality of life in diabetes mellitus patients. The aim of this study was to investigate the efficacy of N-acetylcysteine (NAC) and ascorbic-acid-2-phosphate (AAP) oral supplementation as preventative measure against MSC impairment. Healthy wild-type control (C57BL/6J) (male, n = 24) and obese diabetic (B6.C-Lepob/J) (ob/ob) (male, n = 24) mice received either placebo or antioxidant (NAC/AAP) supplementation for a period of 6 weeks. Metabolic parameters (weight and blood glucose) and the oxidative status (serum total serum antioxidant capacity, malondialdehyde) of animals were assessed. At the end of the 6-week supplementation period, bone marrow MSCs were isolated and their functionality (growth rate, viability, adipogenesis, and osteogenesis) assessed ex vivo. Real time quantitative polymerase chain reaction microarray analysis was also performed to assess the expression of 84 genes related to oxidative stress in MSCs. Despite no change in the metabolic profile, NAC/AAP supplementation improved the antioxidant status of diabetic animals and reduced lipid peroxidation, which is indicative of cellular damage. NAC/AAP also improved the population doubling time of MSCs (first 6-days postisolation) and significantly downregulated the expression of two genes (Nox1 and Rag2) associated with oxidative stress compared to placebo treatment. Taken together, this study has shown reduced oxidative stress and improvements in MSC function following in vivo antioxidant supplementation in healthy control and type 2 diabetic mice.
Introduction
Type 2
Given the persistent systemic inflammation and oxidative stress evident in T2DM, pharmacological and/or natural agents that counteract these factors may have protective effects that could delay disease progression. Various preclinical studies have demonstrated the protective effect of antioxidants, such as N-acetylcysteine (NAC), against diabetes-induced hepatopathy, nephropathy, retinopathy, and myocardial injury in rodent models [8 –11]. This is supported by a meta-analysis of 28 studies illustrating the efficacy of NAC to alleviate inflammation and oxidative stress [12] and an open-labeled pilot study demonstrating the efficacy of NAC to reduce the risk burden in metabolic syndrome patients through its antioxidant and anti-inflammatory properties [13]. Taken together, these studies therefore support the potential clinical use of antioxidants as an adjuvant therapy in the treatment of metabolic disorders such as T2DM.
Bone marrow remodeling and dysfunction are often overlooked as a significant end-organ failure in T2DM. In a recent review of the literature, Vinci et al. [14] highlighted the role of hyperglycemia-induced bone marrow dysregulation in disease progression. The authors suggested that bone marrow-resident stem cells, which include both hematopoietic (HSC) and mesenchymal stem cells (MSC), are actively involved in the onset of complications [14]. Inflammation and oxidative stress-induced cellular senescence hamper the proregenerative and immune-modulatory function of these cells and skew their phenotype to cause further maladaptive tissue damage by amplifying inflammation [14 –17]. This is based on evidence from various preclinical models.
Katagi et al. [16] demonstrated that transplantation of dysfunctional bone marrow stem cells (HSCs) derived from mice with diabetic neuropathy into healthy animals produced neuronal dysfunction in the recipient mice [16]. Similarly, Xu et al. [18] showed that transplanting irradiation-induced senescent stem cells into young mice caused physical dysfunction and reduced lifespan. The authors furthermore provided proof-of-concept evidence that therapeutic intervention could alleviate dysfunction and reduce the mortality rate of recipient mice, supporting the notion that protecting MSCs against the pathological microenvironment could potentially delay disease progression and enhance the quality of life in T2DM patients [17].
Several in vitro studies have demonstrated that the antioxidants, NAC and ascorbic-acid-2-phosphate (AAP), can protect MSCs against hydrogen peroxide-induced oxidative stress [19 –21] and partially rejuvenate dysfunctional diabetic MSCs [15,22,23]. Consistent with this in vitro data, Yan et al. [22] showed that preconditioning of diabetic MSCs with either NAC or a Nox4 siRNA before transplantation reduced oxidative stress and improved the ability of diabetic MSCs to promote postischemic neovascularization in a mouse model of hindlimb ischemia. It is, however, unclear if antioxidant supplementation in vivo will be able to prevent the functional decline of bone marrow MSCs in T2DM. The aim of this study was to investigate the efficacy of in vivo NAC/AAP oral supplementation as preventative measure against MSC impairment in a mouse model of obesity and type 2 diabetes.
Materials and Methods
Animal housing and husbandry
This research study was conducted in accordance with the South African National Standard: The care and use of animals for scientific purposes (SANS10386:2008) and complied with the South African Animal Protection Act (Act no. 71, 1962). Ethical approval (ACU-2019-3857) was granted by Stellenbosch University's Research Ethics Committee: Animal Care and Use (REC:ACU). Healthy wild-type control (C57BL/6J) (male, n = 24) and obese diabetic (B6.C-Lepob/J) (ob/ob) (male, n = 24) mice were housed in groups (2–3 per cage) at a constant temperature of 21°C with a 12-h light/12-h dark cycle and ad libitum access to food (Rodent breeder, Labchef, SA) and drinking water. At age 4–5 weeks, mice were trained to consume a placebo jelly cube (0.5 g, 10% of recommended daily food) before the start of supplementation to ensure consistency in the uptake of oral supplement.
Oral antioxidant supplementation
The jelly cubes were prepared using a mixture of 66% strawberry-flavored gelatin (with a 50% reduced sugar content) [Tower, Pioneer Foods (Pty) Ltd., SA] and 33% unflavored gelatin (Sheridans, SA) dissolved in dH2O. The active antioxidants [7.5 mM NAC and 0.6 mM ascorbic acid-2 phosphate (AAP) (Sigma-Aldrich, Missouri)] or vehicle controls [phosphate buffered saline (PBS) and ddH2O] were added to the jelly mixture at 4°C. The jelly cubes were stored at 4°C and prepared fresh on a weekly basis. The animals received one cube of jelly (containing either placebo or active antioxidants) 5 days per week for a period of 6 weeks. At the end of the supplementation period, animals were thus 11–12 weeks old. Refer to Table 1 for the human/animal equivalent dose calculations. Note: the dose used in this study was based on an in vitro dose–response experiment previously performed and published by our group [15]. The in vivo treatment groups were as follows: C:P—wild-type control mice receiving placebo (n = 10). C:S—wild-type control mice receiving antioxidants (n = 10). DM:P—obese diabetic mice receiving placebo (n = 7). DM:S—obese diabetic mice receiving antioxidants (n = 7). Animal well-being (appearance and behavior) was monitored on a daily basis throughout the study.
Antioxidant Dose
The AED (mg/kg) was based on the body weight of the animals at baseline before the onset of supplementation. The HED was calculated using the following formula: HED (mg/kg) = (AED) × (Mouse Km/Human Km). Km human = 37; Km mouse = 3. Km ratio = 3/37 = 0.081 [35].
AAP, ascorbic-acid-2-phosphate; AED, animal equivalent dose; HED, human equivalent dose; NAC, N-acetylcysteine.
Metabolic measures
The body weight (g) and fasting blood glucose levels (mmol/L) were assessed at baseline and on a weekly basis thereafter for the duration of the study. Fasting blood glucose levels (tail vein prick) were determined using the contour plus glucometer system (Bayer, Leverkusen, Germany). The detectable glucose range for the glucometer is 0.6–33.3 mmol/L (10.8–600 mg/dL).
Serum collection and analysis
At the end of the 6-week supplementation period, the animals were placed under general anesthesia using isoflurane (Safeline pharmaceuticals, SA) gas through vaporizer (Penlon Ltd., UK) for 3–5 min. Immediately, following cervical dislocation, blood was drawn through cardiac puncture using a 25-gauge needle (Avacare™, Lasec, SA) and serum separating tubes (BD Microtainer® SST™). Collected blood samples were left at room temperature for 15 min to coagulate. After coagulation, the tubes were placed on ice and centrifuged at 4,000 RPM for 5 min (Prism™; Labnet, Inc.). Serum was aliquoted (∼200–400 μL per animal) and stored at −80°C until subsequent analysis. Serum samples were analyzed to determine the total antioxidant capacity (E-BC-K136-100 EL; Elabscience) and lipid peroxidation status [Malondialdehyde (MDA), E-BC-K025 EL; Elabscience] according to the manufacturer's instructions.
Isolation of bone marrow-derived MSCs
Bone marrow-derived MSCs were isolated from the femurs of animals as previously described [24]. Following, cervical dislocation, the femurs were dissected out and the proximal and distal ends of each femur cut open in a sterile biosafety cabinet. Bone marrow aspirates were flushed into six-well cell culture plates (femur 1 = well 1 and femur 2 = well 2) using a 25-gauge needle and syringe containing isolation media. Isolation media consisted of Dulbecco's modified Eagle's medium (DMEM) with ultraglutamine (4.5 g/L high glucose; BioWittaker, Lonza, Basel, Switzerland), 1% penicillin/streptomycin (BioWittaker), and 20% fetal bovine serum (FBS) (Biochrom, Berlin, Germany). The bone marrow aspirates were maintained at 37°C in a humidified incubator with 5% CO2 to allow for plastic adherence of MSCs. After 96 h, nonadherent cells were washed off using PBS solution and the media replaced with standard growth media (SGM). SGM consisted of DMEM with ultraglutamine (4.5 g/L high glucose; BioWittaker), 1% penicillin/streptomycin (BioWittaker), and 10% FBS (Biochrom). SGM was changed every 3 to 4 days.
For subculturing, adherent cells (passage 0, 70% confluent) were detached from the wells through enzymatic digestion using prewarmed Accutase (StemProAccutase, gibco). SGM was added to the detached cells to inactivate the enzymatic activity of Accutase and the cell suspension centrifuged at 1,000 RPM for 5 min (Eppendorf centrifuge 5804, Germany).
Flow cytometry
At the end of passage 0, MSCs (80% confluent) from a subgroup of animals [n = 3, obese diabetic (B6.C-Lepob/J)] were harvested through trypsinization, pooled together, and resuspended in PBS containing 1% bovine serum albumin (Sigma-Aldrich, Germany). The cell suspension at a concentration of 1 × 106 cells per 100 μL was stained with the stem cell marker Ly-6A/E (Sca-1) (rat anti-mouse PE-conjugated, #BD561076, BD Pharmingen). Flow cytometry was performed on a FACS Canto II instrument using FACSDiva software. A total of 15,000 events were recorded before data analysis. An unstained control was used as a negative control for gating purposes. Data analysis was performed using Flow Jo software (BD Biosciences).
Crystal violet staining
Following subculture, MSCs (passage 1) were seeded into three 96-well plates at a seeding density of 2 × 104 cells/well (666 cells/mm2). Cell viability was assessed over a period of 144 h (6 days) postseeding using crystal violet (hexamethyl pararosaniline chloride) staining. At 6 days postseeding, the media were removed from each well, the cells washed with PBS, and fixed with 150 μL ice-cold methanol [Sigma Aldrich, Merck chemicals (Pty) Ltd., RSAUS] for 30 s. MSCs were stained with 0.01% crystal violet solution (Sigma-Aldrich, Germany) for 5 min and washed twice with PBS. Tile scan images were taken of each well using a light microscope (Zeis Axio Observer 7 inverted microscope Axiocam 305 color, Germany) at 10 × magnification. Images were analyzed using Image J software (Version 1.52; NIH, Maryland;
Osteogenic and adipogenic differentiation
MSCs (passage 1) were seeded into 96-well plates at a seeding density of 2 × 104 cells/well and allowed to reach 100% confluence in SGM before induction of differentiation. SGM was replaced with either adipogenic induction media (AM) or osteogenic induction media (OM) for a period of 10 days. AM consisted of SGM supplemented with 10 μM insulin (human, I9278-5 mL, Sigma-Aldrich, Missouri), 0.5 mM 3-isobutyl-1-methylxanthine (IBMX; Sigma-Aldrich, Missouri), 1 μM dexamethasone (Sigma-Aldrich, Missouri), and 56 μM indomethacin (Sigma-Aldrich, Missouri). For adipogenic differentiation, AM was changed thrice a week for the duration of the induction period. OM consisted of SGM supplemented with 50 μM ascorbic acid (Sigma-Aldrich, Missouri), 10 nM dexamethasone (Sigma-Aldrich, Missouri), and 10 nM β-glycerophosphate (Sigma-Aldrich, Missouri). For osteogenic differentiation, OM was changed twice a week for the duration of the induction period. Following the 10-day induction period, the extent of lipid accumulation (adipogenesis) and mineralization (osteogenesis) was determined using Oil Red O (0.7% v/v) (Sigma-Aldrich, Missouri) or Alizarin Red S (100 mM; Sigma-Aldrich, Missouri) staining as previously described [24].
For both adipogenesis and osteogenesis, tile scan images were taken at 10 × magnification using a light microscope (Zeiss Axio Observer 7 inverted microscope Axiocam 305 color, Germany). The extent of differentiation (positive stained area) was quantified using Image J software (Version 1.52; NIH;
RNA isolation, cDNA synthesis and polymerase chain reaction micro-array analysis
Total RNA was isolated from MSCs (DM:P and DM:S) using the RNeasy kit (Qiagen, Berlin, Germany) with on-column DNA digestion. An in vitro treatment (NAC/AAP) group was used as positive control: a subgroup of MSCs derived from obese diabetic mice (n = 3, B6.C-Lepob/J) did not receive oral supplementation, instead following isolation, the cells were expanded in the presence of NAC/AAP in culture. Total RNA (0.5 μg) was used as a template for cDNA synthesis using the RT2 First Strand Kit (#330401; Qiagen). cDNA samples were analyzed with an RT2 Profiler polymerase chain reaction (PCR) mouse oxidative stress microarray (PAMM-065ZC-12, 96-well plate format; Qiagen) and RT2 SYBR Green Master mix (#330522, Rox PCR Master mix; Qiagen) using the QuantStudio ™ 5 PCR system (ThermoFisher) and QuantStudio Design & Analysis software (v1.5.1). Relative gene expression was calculated using Gapdh (Glyceraldehyde-3-phosphate dehydrogenase; NM_008084) as housekeeping gene. The PCR array also included a mouse genomic DNA contamination (SA_00106), reverse transcriptase (SA_00104), and positive PCR controls (SA_00103).
Statistical analysis
Statistical analysis was performed using GraphPad Prism (v9.1.1). Analysis of variance (one way and three way) with Tukey post hoc test was used to determine the effect of group (C vs. DM), treatment (S vs. P), and group × treatment. Population doubling time (dT) of MSCs was calculated using the following formula: dT = (T2-T1)/[(log cell number T2 – log cell number T1)/log2]. For the PCR data analysis, the web-based PCR Profiler Array data analysis package was used for ΔΔCt based for change calculations (Qiagen,
Results
Metabolic and antioxidant status of animals
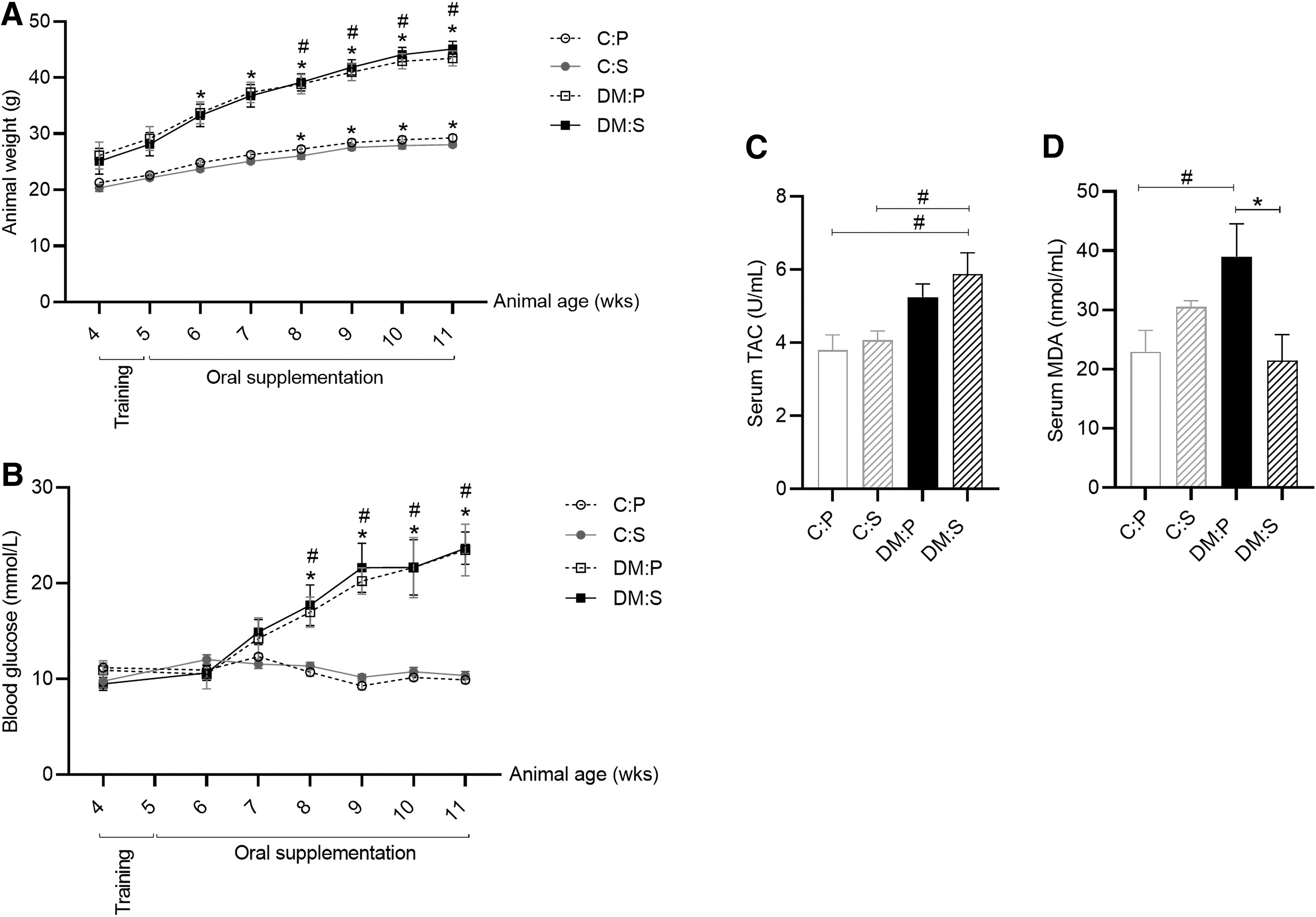
Body weight was assessed on a weekly basis from baseline (age 4 weeks) to the end of supplementation period (age 11 weeks). A normal growth curve was evident in the wild-type control mice (C) with body weight increasing slightly over time from 20.8 ± 1.7 g (mean ± standard deviation, SD) at baseline to 28.7 ± 1.6 g after 6 weeks of supplementation (P < 0.01) (Fig. 1A). Excessive weight gain was evident in the obese diabetic mice (DM), with body weight increasing from 25.6 ± 6 g at baseline to 44.3 ± 3.5 g after 6 weeks of supplementation (P < 0.001) (Fig. 1A). Although a significant difference in body weight was evident between C and DM animals (P < 0.001), there was no difference between antioxidant supplementation and placebo treatment in either of the groups (Fig. 1A).
Metabolic and antioxidant status of animals.
Similarly, blood glucose levels remained constant in C mice throughout the study (10.8 ± 1.5 mmol/L; 194 ± 27 mg/dL) (mean ± SD), whereas a sharp increase was evident in the DM mice from 10.2 ± 2.3 mmol/L (183.6 ± 41.4 mg/dL) at baseline to 23.6 ± 5.7 mmol/L (424.8 ± 102.6 mg/dL) after 6 weeks of supplementation (P < 0.001) (Fig. 1B). A significant difference was thus evident in the blood glucose levels between C and DM animals (P < 0.01) (Fig. 1B), with hyperglycemia indicative of diabetes (>16.7 mmol/L; >300 mg/dL) evident in DM mice from week 8 (age) onward. Antioxidant supplementation did not affect glucose levels in either of the groups (Fig. 1B).
At the end of supplementation period, the DM animals tended to have an overall higher total serum antioxidant capacity (TAC) (DM:S 5.9 ± 1.5 U/mL and DM:P 5.2 ± 0.9 U/mL) than healthy control animals (C:P 3.8 ± 1.3 U/mL and C:S 4.1 ± 0.8 U/mL) (Fig. 1C). A significant difference was evident between the TAC in DM:S and control (C:P and C:S) animals (P < 0.05) (Fig. 1C). Consistent with the TAC data, antioxidants reduced serum MDA levels in DM animals (DM:P 39.0 ± 14.7 nmol/mL and DM:S 21.5 ± 11.6 nmol/mL) (P < 0.01) to levels comparable to that of healthy C animals (C:P 22.9 ± 11.4 nmol/mL) (Fig. 1D).
MSC functional characteristics
Representative images of MSCs (passage 0) derived from each group are presented in Fig. 2A. Flow cytometry analysis demonstrated that at the end of passage 0, >85% of isolated cell populations expressed the stem cell marker Sca-1 (Fig. 2B). Exponential growth (log phase) was evident from day 4 to 8 postisolation in MSCs derived from healthy C animals, whereas growth slowed after day 6 in the MSCs derived from obese DM animals (Fig. 2C). The population doubling (dT) time during the log phase (day 4–6) indicated that in vivo antioxidant supplementation was able to improve cellular growth and reduce the dT in MSCs derived from both groups (C:P 14 days; C:S 12 days; DM:P 15 days; and DM:S 5 days) (P < 0.05) (Fig. 2D). This effect was, however, not sustained and only evident during the first 6 days postisolation. In both C and DM animals, in vivo antioxidant supplementation slightly increased (1.5-fold) the yield of MSCs at the end of passage 0 (before subculture); due to variability, this observation was, however, not significant (data not shown). Following subculture (passage 1), the survival rate of MSCs derived from C mice was significantly improved by the in vivo antioxidant supplementation (C:P vs. C:S) (P < 0.05) (Fig. 3A). Refer to Fig. 3A for representative Crystal Violet images taken at 6 days post-subculture.
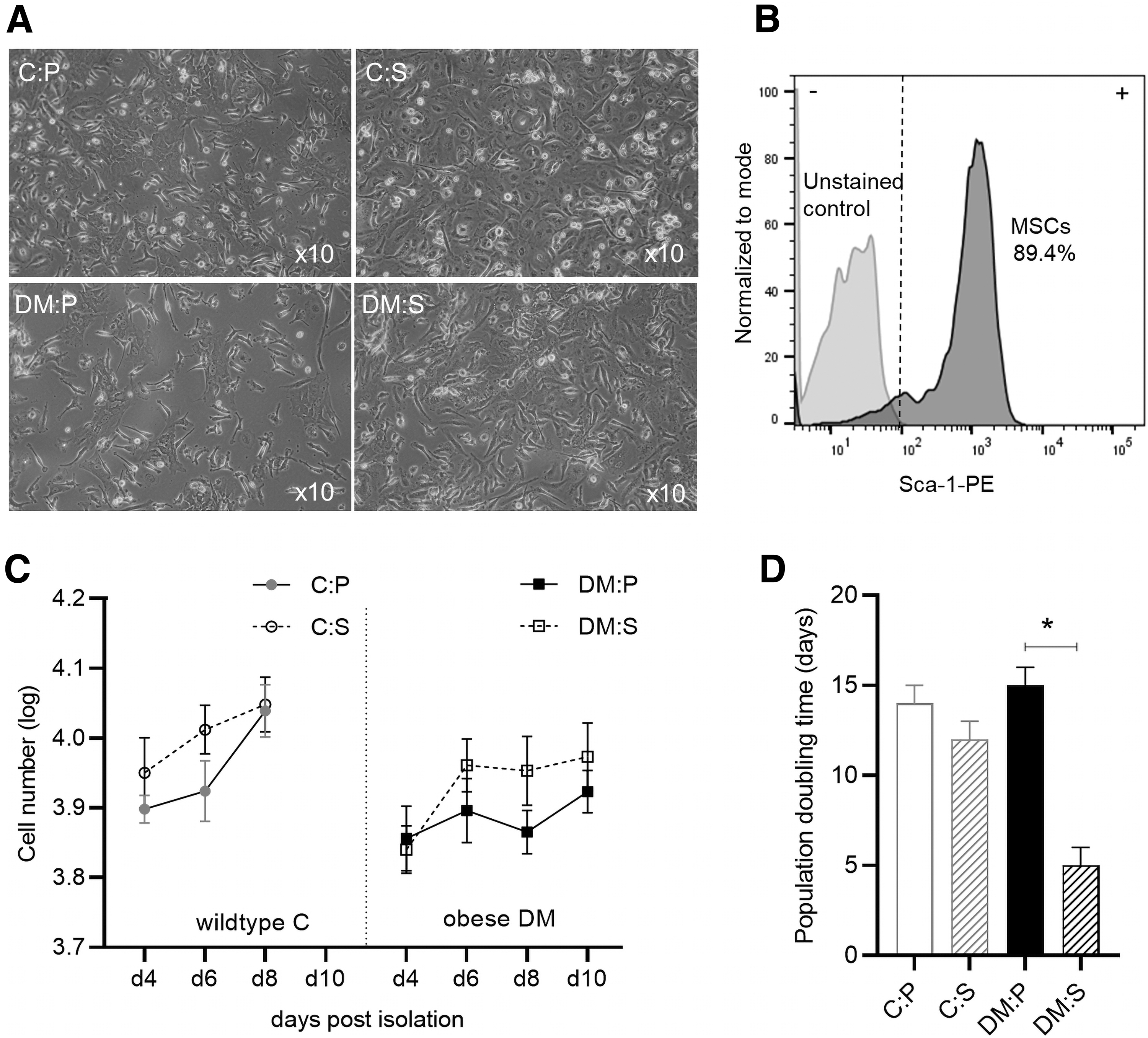
MSC growth postisolation (passage 0).
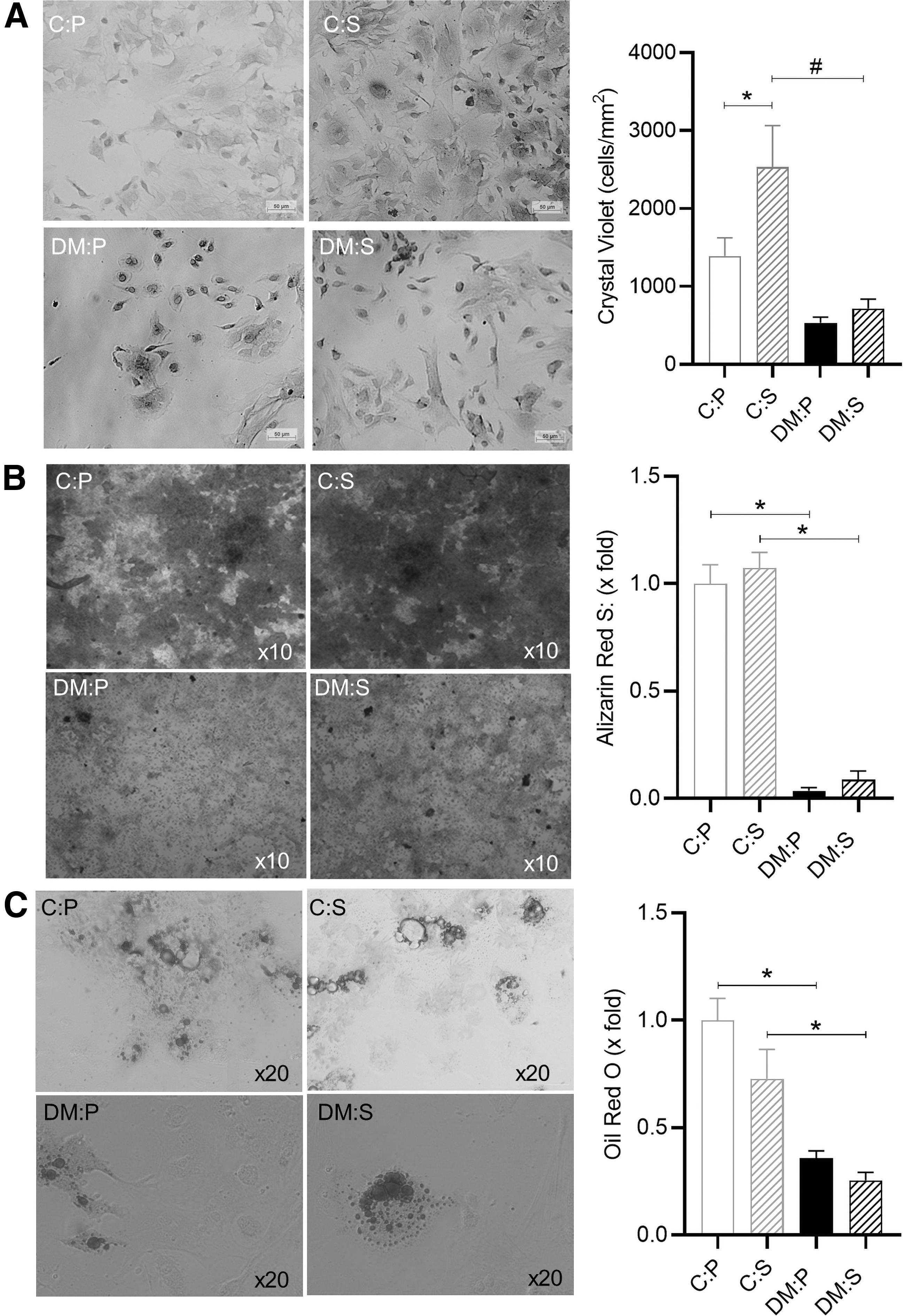
MSC functionality following subculture (passage 1).
The osteogenic and adipogenic differentiation capacity of MSCs were also determined after subculture (passage 1). Extensive mineralization (>60% of surface area) was evident in the MSCs derived from healthy C animals, whereas osteogenic differentiation was significantly impaired in the MSCs derived from DM animals (<10% surface area) (P < 0.001) (Fig. 3B). Although in vivo antioxidant supplementation did show some signs of improved osteogenesis in the DM:S group (6% ± 3% surface area) compared to placebo treatment (DM:P 2% ± 1% surface area), it was not significant (Fig. 3B). Similarly, lipid accumulation during adipogenesis was impaired in the MSCs derived from DM mice compared to healthy C mice (P < 0.01) (Fig. 3C).
PCR array data
The relative expression level of a total of 84 genes related to oxidative stress is presented in Fig. 4A. The use of an in vitro treatment group as positive control (+control) demonstrated that NAC/AAP was able to significantly reduce (>2-fold; P < 0.05) the expression of 33 genes related to oxidative stress in MSCs derived from DM animals (Table 2; Fig. 4A, B). In vivo antioxidant supplementation showed a similar trend and slightly reduced the expression of nine genes (Apoe, Gclm, Idh1, Nqo1, Prnp, Ucp3, Prdx1, Nox1, and Rag2) of interest in the DM:S group (Table 2; Fig. 4B). In comparison with placebo treatment (DM:P), two genes of interest (Nox1 and Rag2) were significantly (>2-fold; P < 0.05) downregulated in DM:S (Fig. 4B, C).

Gene expression: qPCR array.
Relative Gene Expression (Compared to Placebo Treatment)
The data are presented as relative gene expression compared to the placebo treatment (DM:P) group. It is calculated using the ΔΔCt method, with data normalized [AVG Delta(Ct) (Ct(GOI)−Ave Ct(HKG)] using the housekeeping gene, Gapdh.
GOI, gene of interest; HKG, housekeeping gene; Rag2, recombination activating gene 2.
Discussion
Diabetes is a global epidemic. The complex and multifactorial nature of the disease cause aberrant changes in the microenvironment, which not only cause tissue damage but also desensitize and dysregulate the function of cellular mediators of repair [1,3,5,14]. The functional decline of stem/progenitor cells is known to be a contributing factor in disease progression and the development of comorbidities [14,17]. Stem cell defects in T2DM include DNA damage, mitochondrial fragmentation, senescence with an altered secretome, impaired viability, and reduced proliferation, migration, and multilineage differentiation capacity [5,15,19 –21,23].
Evidence from in vitro studies indicate that antioxidants such as NAC/AAP can improve the ex vivo viability of bone marrow-derived MSCs, can partially restore the paracrine function of impaired diabetic MSCs, and has protective effects against acute oxidative stress insults [15,19 –21,23]. There is, however, a paucity of studies assessing the beneficial effect of these therapeutic factors in preventing the functional decline of stem cells in vivo. In this study, we demonstrated slightly improved MSC growth and viability postisolation and a significant reduction in the expression of two genes (Nox1 and Rag2) related to oxidative stress following 6 weeks of in vivo NAC/AAP supplementation. These effects were observed in standard cell culture conditions, without continued NAC/AAP treatment, suggesting that the diabetic MSCs were able to partially retain the beneficial effects from in vivo supplementation. These observations were evident despite no change in the metabolic profile of animals.
In this study, the low-dose oral supplementation for a relatively short period of time (6 weeks) was able to improve the total antioxidant capacity and reduce lipid peroxidation in diabetic animals. This is consistent with other rodent studies demonstrating improved antioxidant capacity and reduced oxidative damage in specifically liver tissue following short-term low-dose antioxidant treatment. Genç & Peker (2021) showed that daily intraperitoneal injection of 60 mg/kg NAC for a period of 3 weeks prevented hepatopathy in streptozotocin-induced type 1 diabetic rats by improving glutathione (GSH) levels and reducing lipid peroxidation within tissue [9]. In type 2 diabetic models, potential changes in the metabolic profile should, however, also be considered during the treatment period. The metabolic parameters assessed in this study clearly indicated that the DM animals had excessive weight gain and hyperglycemia compared to the healthy control animals. The oral supplementation did not alter, improve, or prevent metabolic decline in DM animals and can therefore not replace standard primary antidiabetic treatment. This observation is consistent with a previous study done by Szkudlinska et al. [26], indicating that short-term NAC supplementation in well-controlled (glycosylated haemoglobin, HbA1c <7%) middle-aged diabetic patients did not improve glucose tolerance and β-cell function [26]. Further studies are, however, required to determine if NAC/AAP affects tissue-/organ-specific insulin resistance and hyperinsulinemia.
The potent antioxidant capacity of NAC and AAP is, however, well established [27,28], suggesting that the effects observed in this study were mainly due to the promotion of the antioxidant defense mechanisms. This was confirmed by a reduction in serum MDA levels with supplementation in DM animals. Free radicals (oxidative stress) cause peroxidation of lipids and MDA is one of the final products of peroxidation of polyunsaturated fatty acids in cells [29]. A reduction in serum MDA therefore indicated that the free radical scavenging capacity of NAC/AAP reduced oxidative stress and cellular damage in DM animals. This observation is supported by a previous 15-day randomized controlled trial, which showed improved postprandial oxidative stress in diabetic patients receiving a combination of NAC, vitamin C (ascorbic acid), and vitamin E [30]. Taken together, the in vivo data therefore confirm that the low-dose, short-term supplementation in this study was able to reduce oxidative stress.
Despite the promising in vivo data, the beneficial effect on MSCs was short lived postisolation without the continued presence of NAC/AAP. The improved cellular growth rate (reduced population dT) in MSCs derived from both control and DM animals that received supplementation was only evident for the first 6 days postisolation. Following subculture, increased viability, osteogenesis, and a tendency to reduce adipogenesis were only evident in the MSCs derived from healthy control mice, suggesting that without the continued support of NAC/AAP, diabetic MSCs will revert back to their impaired status. Despite the modest nature of functional improvements observed, it is consistent with the beneficial in vitro treatment effects described in the literature, [15,19 –21,23] and might be more profound when supplementation is given for a longer period at higher concentrations. Indeed, a recent long-term (12 month) high-fat high-sucrose diet-induced DM murine study demonstrated that early NAC intervention (6–12 months) reduced cellular senescent markers, improved mitochondrial function, reduced oxidative protein/DNA damage, and reduced inflammation within adipose tissue [31]. Although the authors did not assess MSC function, a reduction in senescent markers following 6 months of antioxidant intervention supports our suggestion that prolonged treatment might show more profound improvements in MSC function.
In vitro treatment of diabetic MSCs with NAC/AAP as a positive control confirmed that it completely downregulated the expression of 28 out of 84 genes of interest assessed, which are associated with oxidative stress. A similar trend was evident with in vivo supplementation; however, only NADPH oxidase (Nox1) and recombination activating gene 2 (Rag2) remained significantly downregulated in DM MSCs postisolation. NADPH oxidases (Nox) are enzymes dedicated to the production of reactive oxygen species (ROS) with Nox1 and Nox2 being the major source of oxidative stress in microvascular complications [32,33]. This study therefore suggests that downregulation of Nox1 expression with NAC/AAP supplementation is limiting the production of ROS formation, in addition to its free radical scavenging capacity, and warrants further investigation. Nox1-derived ROS formation has been implicated in endothelial dysfunction, retinopathy, cytokine-induced β-cell dysfunction, premature cellular senescence, and nephropathy in T2DM [32 –38]. Therapeutic intervention targeting NADPH oxidases (Nox inhibitors) to limit oxidative stress-induced damage underlying T2DM complications has been previously proposed [39]. In this study, we demonstrate the ability of NAC/AAP to downregulate specifically Nox1 expression in dysfunctional diabetic MSCs. The role of Rag2 expression and its downregulation with supplementation are, however, less clear.
The recombination activating genes (Rag1/2) are involved in immune function, specifically the maturation of lymphocytes, but its activity can also cause double-stranded breaks by cleaving DNA [40]. Under normal healthy conditions, the DNA damage response activates an ataxia-telangiectasia mutated kinase-dependent negative feedback loop, which downregulates Rag1/2 expression to prevent further damage [40]. Hladik et al. [41] did, however, demonstrate that prolonged ex vivo culture of MSCs impairs their ability to recognize DNA breaks, which causes genomic instability. Similarly, oxidative stress-induced DNA damage and genomic instability are prevalent in dysfunctional diabetic MSCs [5]. Despite showing reduced Rag2 gene expression, it was beyond the scope of this study to determine the involvement of Rag2 in either DNA damage or the immunomodulatory properties of MSCs.
Taken together, we have, however, shown reduced oxidative stress and modest improvements in MSC function following in vivo antioxidant supplementation in healthy control and type 2 diabetic mice. Further investigation is, however, required to examine the underlying mechanisms associated with Nox2 and Rag2 activity in tissues such as the pancreas, muscle, and liver, which are sensitive to oxidative damage. Nonetheless, taken together with evidence from literature, the overall reduction in oxidative stress achieved with NAC/AAP as adjuvant treatment could potentially delay the onset of diabetic comorbidities and warrants further investigation. Long-term patient-based studies are required to determine whether antioxidants as supportive treatment for diabetic patients can delay the onset of comorbidities.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a grant from the South African Sugar Association. M.M. was supported by a bursary from SA Akademie.