
Abstract
Inflammation, the main factor in the progression of osteoarthritis (OA), impairs the chondrogenesis of bone mesenchymal stem cells (BMSCs), which is an appealing process to target to regenerate impaired articular cartilage. This article aimed to investigate whether SP600125, a competitive ATP-specific inhibitor of the JNK pathway, could promote the chondrogenesis of BMSCs by enhancing their anti-inflammatory capacity. Chondrogenic differentiation was assessed by Alcian blue staining, immunofluorescence staining, and Western blot. The inflammation level was associated with the expression of matrix metalloproteinases (Mmp), evaluated by Western blot. Intra-articular injection of BMSCs pretreated with or without SP600125 was carried out on C57BL/6 mice after inducing OA by surgical destabilization of the medial meniscus. Safranin O-fast green (SO) and hematoxylin-eosin staining were employed to evaluate the cartilage destruction and immunohistochemical analysis was adopted to detect the expression of Col2 and Mmp-13 proteins in the mouse knee joint. We showed that SP600125 could inhibit inflammation induced by tumor necrosis factor-α (TNF-α) and promote the chondrogenesis of BMSCs. In the presence of TNF-α, the expression of aggrecan (Agc) and collagen type II alpha 1 (Col2) was significantly decreased compared with that in the control group and increased with the addition of SP600125. Moreover, the expression of Mmp-1, Mmp-3, and Mmp-13 was increased in BMSCs treated only with TNF-α and downregulated in SP600125-treated BMSCs. In vivo study showed that SP600125 could enhance protective effects of BMSCs on OA mice. Our results indicated that SP600125 rescued the chondrogenesis of BMSCs by inhibiting inflammation induced by TNF-α, which provides a theoretical basis for solving the problem of cartilage repair under inflammatory conditions.
Introduction
Osteoarthritis (OA), the most common degenerative disease of the joint worldwide, is characterized by alterations in the whole joint, including the articular cartilage, subchondral bone, ligaments, capsule, and synovial membrane, ultimately resulting in joint failure [1,2]. Various risk factors, such as age, obesity, injury, genetics, wear, and inflammation, are involved in the development and progression of OA [3,4]. Patients with OA suffer from chronic pain and mobility inconvenience for a long time and eventually are saved from total joint replacement when the disease develops to the point of surgery [4]. Statistics in 2013 showed that the combined cost of lost wages and medical care due to OA exceeded $300 billion [5].
Accumulating evidence has demonstrated that inflammation, especially low-grade inflammation, acts as a main contributor to OA even in the early stage of the disease [3,6], which reprograms chondrocytes to hypertrophic differentiation and proinflammatory and procatabolic responses, thus reducing their viability [7]. Tumor necrosis factor-α (TNF-α) is considered to be a major proinflammatory cytokine secreted by chondrocytes, osteoblasts, mononuclear cells, and synovial tissues, the levels of which are significantly enhanced in the synovial fluid, synovial membrane, subchondral bone, and cartilage of patients with OA [6]. Overexpression of TNF-α can increase the gene expression of matrix-degrading enzymes such as matrix metalloproteinases (Mmp), which in turn promote the degradation of the cartilage extracellular matrix (ECM) and further facilitate the production of proinflammatory cytokines and synovial inflammation, leading to cartilage destruction [8,9].
The JNK pathway participates in various cell biology reactions, including cellular proliferation, differentiation, development, inflammation, and apoptosis [10]. In mice with OA, the JNK pathway can be activated by TNF-α and further triggers the degradation of aggrecan (Agc), an essential component of the ECM [11].
Bone mesenchymal stem cells (BMSCs) are prospective candidates for articular cartilage regeneration, which can be readily harvested and possess self-renewal, multidifferential, and anti-inflammatory capacities [12,13]. However, regardless of the ability to differentiate into osteocytes, chondrocytes, and adipocytes under specific conditions, the chondrogenesis of BMSCs is blocked by inflammation, constraining their utilization in the cartilage regeneration of joints [14].
In a previous study, we reported that SP600125, a competitive ATP-specific inhibitor of the JNK pathway, could augment the antiapoptotic capacity and migration of BMSCs treated with TNF-α [15], demonstrating its potential role in improving its application under inflammatory conditions. However, it has not been clarified whether SP600125 can promote the chondrogenic differentiation of BMSCs. In this study, we further investigated the effects of SP600125 on TNF-α-impaired chondrogenesis of BMSCs, and explored the protective influences of SP600125-pretreated BMSCs on OA in destabilization of the medial meniscus (DMM) model mice, hoping to provide a potential targeted method for cartilage repair under inflammatory conditions.
Materials and Methods
Cell culture
The BMSCs of Sprague-Dawley rats, which were positive for CD44, CD29, and CD90 and negative for CD45, CD34, and CD11, were purchased from Cyagen Corporation (Guangzhou, China). The BMSCs of C57BL/6 mice, which were positive for CD29, CD44, and Sca-1 and negative for CD117, were purchased from Cyagen Corporation. BMSCs were maintained in DMEM/F12 (HyClone, UT) containing 10% FBS (Gemini, CA) at 37°C with 5% CO2. When the cells reached 80% confluence, the BMSCs were digested, centrifuged, resuspended, and used between passages 3 and 8.
Chondrogenic differentiation
BMSCs digested with 0.25% trypsin-EDTA solution were seeded in six-well plates for chondrogenic differentiation culture as described previously [16]. In short, the BMSCs were trypsinized, washed, and then resuspended at 2 × 107 cells/mL in chondrogenic differentiation medium consisting of DMEM low glucose supplemented with 2.5% FBS, 1 × Insulin-Transferrin-Selenium Solution (Gibco Life Technologies, CA), 1 mM sodium pyruvate (Solarbio, Beijing, China), 37.5 μg/mL ascorbic acid (Solarbio), and 10 ng/mL transforming growth factor-beta 1 (Peprotech, NJ). Droplets of the cell suspension were carefully placed in the center of a well of a 24-well plate. Cells were allowed to adhere at 37°C for 2 h, followed by the addition of 500 μL chondrogenic induction medium and treatment with or without SP600125 before incubation with 10 ng/ml TNF-α (Peprotech). The medium was renewed every 3 days and the pellets were harvested on days 7, 14, and 21.
Cell viability studies
The viability of the BMSCs was evaluated by MTT assay. The BMSCs were digested, diluted to 8 × 104 cells/mL, and then incubated in a 96-well plate for 24 h. When the confluence reached ∼80%–90%, the BMSCs were incubated with or without SP600125 (Selleck Chemicals, TX) for 1 h before treatment with TNF-α for the next 1, 7, or 14 days. Each treatment group was set up with four replicate wells. Subsequently, each well was supplied with 20 μL MTT solution (Sigma, MO) for 4 h. After that, the medium was discarded, and 150 μL DMSO (Sigma) was added to dissolve the MTT crystalline solid. Then, the absorption was measured at a wavelength of 570 nm.
Quantitative real-time polymerase chain reaction
The mRNA expression of cartilage differentiation-related genes, including Agc and SRY-box 9 (Sox9), was examined by quantitative real-time polymerase chain reaction (qRT-PCR). Total RNA was obtained by employing TRIzol reagent (Takara Biotechnology, Dalian, China) and reverse transcribed to cDNA using the PrimeScript™ RT reagent Kit (Takara Biotechnology) according to the manufacturer's protocols. The investigation of mRNA expression by PCR was conducted using a SYBR® Premix Ex Taq™ II kit (Takara Biotechnology) under the manufacturer's protocols. The investigation of mRNA expression in PCR was conducted using SYBR Premix Ex Taq II kit (Takara Biotechnology) with a fluorescence quantitative PCR instrument (ROCHE). GAPDH was used as the internal control for normalization. The relative mRNA expression of Agc and Sox9 was calculated using the 2–ΔΔCt method with reference to GAPDH. All the primers involved are listed in Table 1.
Primers used for Quantitative Real-Time Polymerase Chain Reaction
Alcian blue staining
The BMSCs were differentiated into cartilage cells for 14 days and then washed thrice with PBS. Subsequently, the BMSCs were stained with Alcian blue (Leagene, Beijing, China) and washed with PBS again. After that, the morphology of induced cartilage cells was observed and captured under an inverted microscope.
Three-dimensional culture assay
The BMSCs were transferred into 15 mL centrifuge tubes for centrifugation at 150g for 5 min. The precipitate was resuspended using chondrogenic differentiation medium at a density of 5 × 105 cell/mL. After that, the droplets were carefully transferred into 15 mL centrifuge tubes for centrifugation at 150g for 5 min. Next, the BMSCs were incubated in a constant temperature incubator at 37°C with 5% CO2 for 21 days. During differentiation, the culture solution was replaced with rat BMSC differential medium every two days, and then the bottom of the centrifuge was flicked to suspend the cartilage aggregates. After incubation for 21 days, the morphology and size of cartilage pellets in each group were observed by photography and then fixed in 4% paraformaldehyde for 3 h and stored at −80°C.
Western blot analysis
The BMSCs were pretreated with SP600125 (10 μM) for 1 h, supplied with TNF-α for 24 h, and induced by chondrogenic differentiation medium. The collection of total protein was conducted by centrifugation at 150g for 10 min after washing twice with ice-cold PBS and lysing with RIPA lysis buffer. Each protein sample calculated by BCA protein assay was separated by SDS-PAGE and transferred onto a PVDF membrane. Next, the membranes were cultured with 5% skim milk in 100 mL TBST solution. After that, membranes were removed from the blocking reagent, washed with TBST for 5 min, and marked by different rabbit polyclonal antibodies, including Agc (Proteintech Group, Hubei, China), Col2 (Bioss, Beijing, China), Mmp-1 (Wangleibio, Liaoning, China), Mmp-3 (Proteintech Group), Mmp-13 (Proteintech Group), and GAPDH (Goodhere, Zhejiang, China), at 4°C overnight. Subsequently, a goat anti-rabbit secondary antibody was added to the TBST solution at a ratio of 1:5,000, which was incubated with the membranes for 2 h. An ECL imaging system was utilized to analyze the densities of the proteins according to the manufacturer's protocol.
Immunofluorescence staining
Immunofluorescence staining was utilized to detect Col2 secreted by chondrocytes after the differentiation of the BMSCs. Briefly, following the chondrogenic differentiation of the BMSCs, the cells were immobilized with precooled paraformaldehyde at room temperature for 20 min, permeabilized with Triton X-100, and blocked with goat serum for 1 h. After each step mentioned above, the cells were washed with PBS solution twice. Col2 was incubated with a rabbit polyclonal antibody against rat Col2 (1:100) overnight at 4°C and then cultured with an FITC-conjugated goat anti-rabbit IgG (1:100) for 2 h. Following staining with 4′,6-diamidino-2-phenylindole (DAPI) for 15 min in the dark place, Col2 was observed by inverted fluorescence microscopy.
Animal models
Twelve-week-old male C57BL/6 mice purchased from SJA Laboratory Animal Co. Ltd. (Hunan, China) were divided into three groups randomly: sham group, BMSC group, and BMSC-SP600125 group. The OA model was induced by surgical DMM of the right knee. In detail, the DMM surgery was performed by severing the medial meniscotibial ligament and the sham operation was simply performed by exposing the medial meniscotibial ligament. After DMM surgery for 4 weeks, the mice were given multiple intra-articular injections of 10 μL PBS with 2.5 × 107 cells/mL BMSCs for 6 weeks (once a week) with a micro syringe. Before intra-articular injection, BMSCs were incubated with or without SP600125 for 1 h. All animal experiments were approved by the University of South China Institutional Animal Use and Ethics Committee (Hengyang, China; IRB approval No. USC20200816XS).
Specimen preparation and histological analysis
Mice were sacrificed at 6 weeks following intra-articular injection of BMSCs. Knee joints were separated and fixed with 4% paraformaldehyde. Samples were decalcified in 20% formic acid, and then embedded with paraffin. Five-micrometer sections were collected to obtain serial sagittal sections, including the entire joint. Safranin O-fast green (SO) and hematoxylin-eosin (HE) staining were used for histological analysis. Osteoarthritis Research Society International (OARSI) scoring system was adopted to evaluate the histological grading of cartilage degeneration [17].
Immunohistochemical analysis
Three percent H2O2 was used to block endogenous peroxidase activity. After antigen retrieval with 0.1% trypsin, sections were blocked with goat serum for 1 h and then incubated at 4°C overnight with different antibodies, including Col2 (1:800; Proteintech) and Mmp-13 (1:200; Proteintech). Appropriate secondary antibodies were used to incubate samples. After washing with TBS, horseradish peroxidase-conjugated streptavidin-biotin was added to sections. Immunoreactivity was visualized with 3,3′-diaminobenzidine followed by counterstaining with hematoxylin. ImageJ 1.44p software was used to count stained positive regions and the total number of chondrocytes. The percentage of positive stained cells was determined by stained positive regions divided by the total number of chondrocytes of different groups (compared to sham group).
Statistical analysis
In this study, SPSS 21.0 software was used for all statistical analysis, and the comparison of means was conducted by t-tests. Statistical data are presented as the mean ± standard deviation, and P < 0.05 was considered statistically significant.
Results
Cell activity evaluation and preliminary analysis of chondrogenic differentiation
MTT was adopted to investigate the viability of cells treated with TNF-α in the presence and absence of SP600125 during the process of chondrogenic differentiation. As shown in Fig. 1A, the cell viability of the different groups was not significantly different at various time points, implying that TNF-α and SP600125 did not impact the viability of the BMSCs.
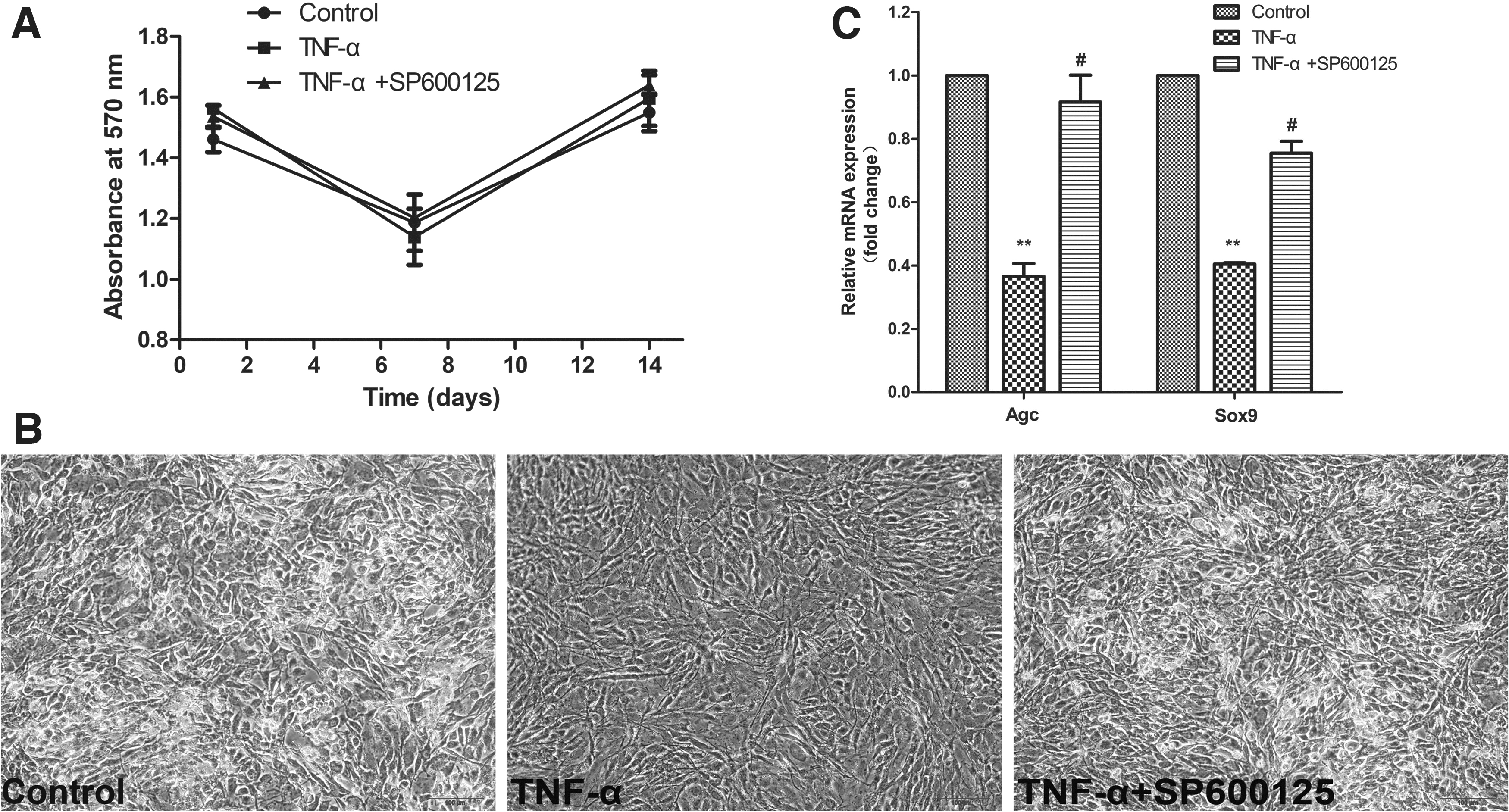
Cell activity evaluation and preliminary analysis of chondrogenic differentiation.
Cartilage-specific proteoglycan in chondrocytes was dyed by Alcian blue staining and further photographed with fluorescence microscope to determine the phenotype of BMSCs or chondrocytes. It was observed that cells cultured in the plate with TNF-α treatment tended to be long and stringy, which were rarely stained by Alcian blue staining compared to the control group. However, the result was conversed by the application of SP600125 (Fig. 1B).
The chondrogenic differentiation potential of the BMSCs was evaluated after the cells had been incubated for 14 days under chondrogenic induction conditions. The qRT-PCR results demonstrated that the mRNA expression of genes related to chondrogenic differentiation, including Agc and Sox9, was downregulated by TNF-α treatment compared with the control group and was increased by pretreatment with SP600125 (Fig. 1C).
Expression of proteins associated with chondrogenic differentiation
Immunofluorescence staining was further performed to investigate the expression of the chondrogenic marker Col2 during BMSC differentiation, which is consistent with the Western blot results (Fig. 2A, B). Western blot analysis indicated an obvious decrease in Agc and Col2 protein levels in the BMSCs treated with TNF-α, which was significantly augmented by the addition of SP600125 (Fig. 2C, D).
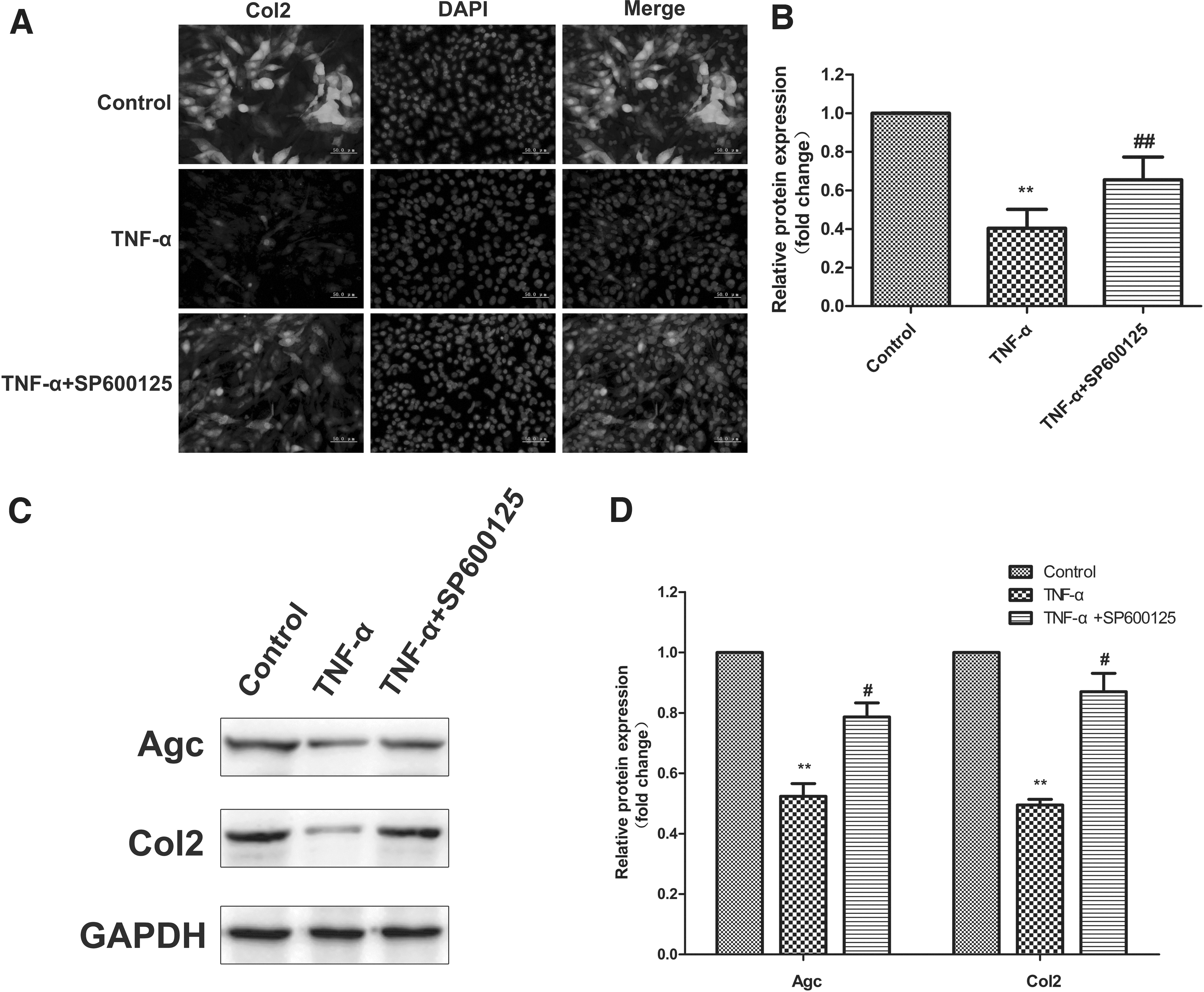
Expression of proteins associated with chondrogenic differentiation.
Chondrogenic differentiation of cells in the three-dimensional pellet culture
To assess the cartilage formation of BMSCs, a three-dimensional (3D) pellet culture was utilized. In comparison to the control group, the cartilage balls treated with TNF-α were smaller and rough and showed irregular shapes. SP600125 treatment significantly improved pellet formation, making the cartilage ball larger, more regular, and symmetrical (Fig. 3A). The cartilage ball was further frozen and sectioned and dyed with Alcian blue staining to observe the chondrogenic differentiation of BMSCs. The results showed that the Alcian blue staining of cells in the 3D pellet culture was more obvious than that in the plate cultivation, suggesting that 3D pellet culture was more beneficial to the chondrogenesis of BMSCs (Fig. 3B, C).

Chondrogenic differentiation of cells in 3D pellet culture and expression of inflammation-related proteins.
Expression of inflammation-related proteins
Under inflammatory conditions, chondrocytes can produce a series of Mmp to promote the degradation of ECM. Western blot was conducted to analyze the quantitative expression of Mmp-1, Mmp-3, and Mmp-13 in chondrogenic differentiation-induced BMSCs treated with TNF-α in the absence or presence of SP600125. TNF-α treatment significantly enhanced the levels of Mmp-1, Mmp-3, and Mmp-13, which were blocked by the presence of SP600125 (Fig. 3D, E), demonstrating that SP600125 exerted an effect on Mmp-associated processes to rescue the chondrogenic differentiation of BMSCs impaired by TNF-α.
Detection of articular cartilage in OA mice
To evaluate the histological changes of OA in DMM operated mice, we observed the cartilage destruction of the mouse knee joint by SO and HE staining. No pathological change of OA was observed in sham group mice (Fig. 4A). In contrast, DMM-induced mice with treatment of BMSCs presented localized injury characterized by vast proteoglycan loss, cartilage erosion, cartilage fibrillation, and cartilage superficial destruction (Fig. 4A). However, the pretreatment of BMSCs with SP600125 could effectively alleviate the cartilage damage of the mouse knee joint induced by DMM operation (Fig. 4A). In addition, OARSI scoring system was applied to evaluate the histological conditions, and we found that DMM-induced mice injected with BMSCs had remarkably higher scores in comparison to sham group, with obviously downregulated OARSI scores after injecting SP600125-pretreated BMSCs (Fig. 4B). These results suggested that SP600125 could enhance the protective effects of BMSCs on OA in DMM mice.
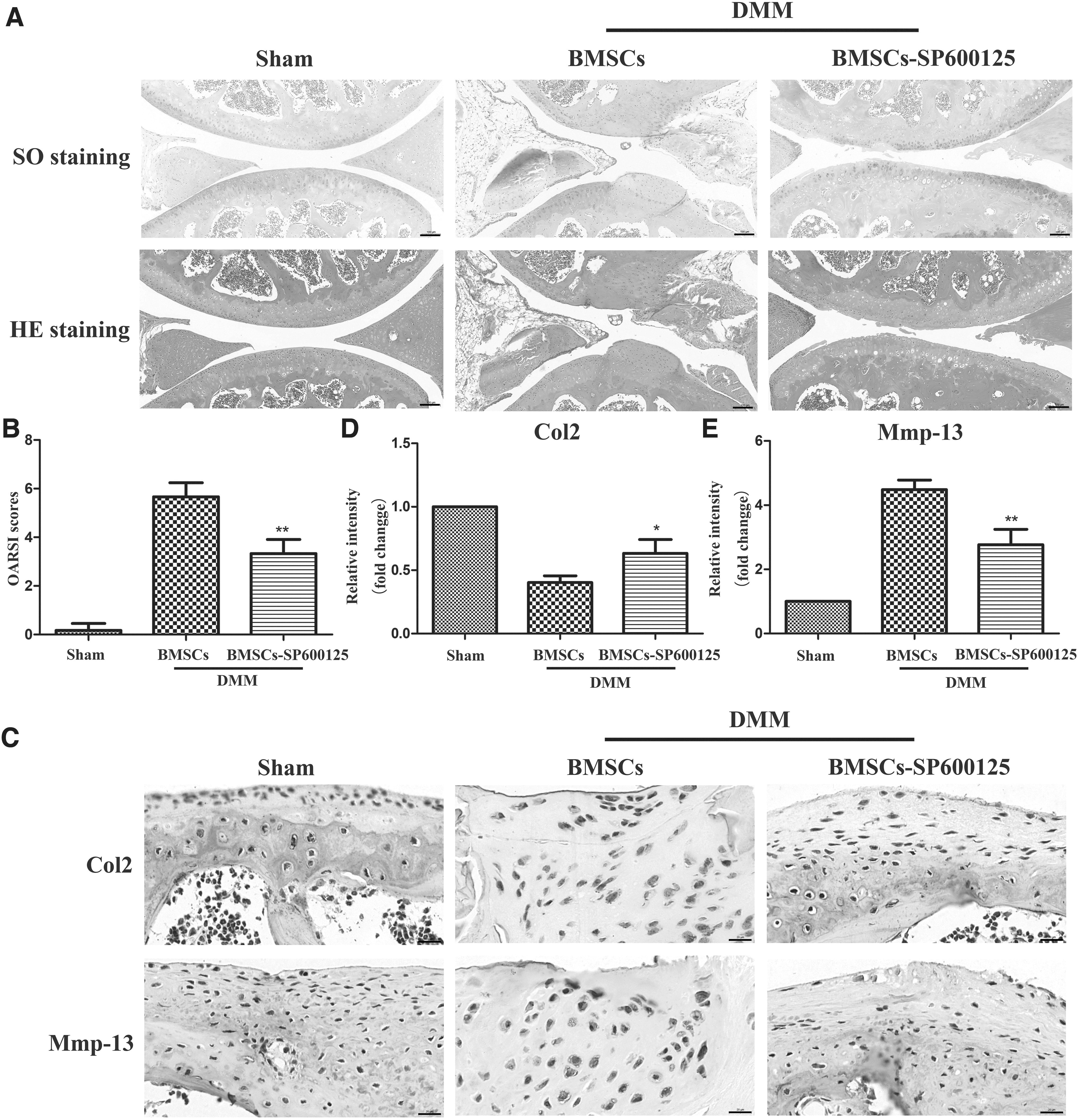
Detection of articular cartilage in OA mice.
To further explore the functional proteins and inflammation of mouse knee cartilage in different groups, immunohistochemical technology was used to detect the expression of Col2 and Mmp-13 proteins. As shown in Fig. 4C, D, and E, DMM-induced mice with intra-articular injection of BMSCs presented reduced expression of Col2 as well as increased level of Mmp-13 compared to mice in sham group, which was reversed by the pretreatment of BMSCs with SP600125. The results showed that SP600125 could enhance the protection of BMSCs on OA by inhibiting matrix degradation and inflammation.
Discussion
OA, whose main pathological basis is cartilage degeneration, can seriously affect the quality of life of patients. The use of chondrogenic differentiation of BMSCs for the treatment of OA has a good theoretical basis and prospect for application. The main manifestation is that inflammatory factors trigger synovitis and accelerate the destruction of articular cartilage, thus causing joint pain [18,19]. Inflammatory factors not only promote cartilage destruction but also inhibit the repair of articular cartilage by BMSCs. In a mouse model of OA, TNF-α was found to activate the JNK-2 gene and thus degrade proteoglycans in cartilage [11]. Therefore, this study modified rat BMSCs by inhibiting the JNK signaling pathway and further explored whether the chondrogenic differentiation ability of modified BMSCs could be improved by treatment with TNF-α, thus providing a theoretical basis for the treatment of OA.
As a competitive ATP-specific inhibitor of JNK discovered in 2001, SP600125 can block the catalytic region of all subtypes of JNK and reversibly inhibit JNK activity [20,21]. SP600125 and TNF-α have certain cytotoxicities under certain concentrations and treatment modes [15,22]. Therefore, we first confirmed that TNF-α and SP600125 used in this study showed no cytotoxicity to BMSCs in terms of time, concentration, and treatment used (Fig. 1A).
Alcian blue staining solution can stain cartilage-specific proteoglycans [23], which is a simple and feasible method to judge chondrogenic differentiation. We found that SP600125 could enhance the chondrogenic differentiation of BMSCs treated with TNF-α by analyzing the changes in cell morphology and the staining effect of Alcian blue in each group (Fig. 1B). Agc and Sox9 are the signature genes of chondrogenic differentiation in BMSCs [24]. Our qRT-PCR results for these two genes indicated that SP600125 enhanced the chondrogenic differentiation of BMSCs inhibited by TNF-α at the gene level (Fig. 1C). Col2 is an important component of the cartilage matrix and a functional protein of cartilage tissue. To intuitively observe the expression of Col2 in the cells after chondrogenic differentiation, immunofluorescence imaging technology was used to analyze the cells. The results showed that SP600125 reduced the inhibitory effect of TNF-α on the Col2 protein (Fig. 2A, B).
Considered the hyaline cartilage, articular cartilage is mainly composed of chondrocytes and ECM, which mainly includes collagen and proteoglycan. Col2 is a characteristic marker of articular cartilage collagen that accounts for ∼90%–95% of the total cartilage collagen. Because proteoglycan has a strong hydrophilicity, it can combine with a large number of water molecules to form a sticky gelatinous substance [25]. To analyze the matrix formation ability of differentiated cartilage, we quantitatively assessed the expression of the Agc and Col2 proteins in each group by Western blot. The results suggested that SP600125 could effectively enhance the matrix formation ability of differentiated cartilage inhibited by TNF-α (Fig. 2C, D).
Currently, there are many uncontrollable factors in animal arthritis models, including the type and concentration of inflammatory cytokines, mechanical stimulation, oxygen partial pressure, acid-base environment, and so on. Therefore, we applied the 3D culture method, which is closer to the environment in vivo [26], to detect the chondrogenic differentiation ability of BMSCs in each group. The results suggested that SP600125 could effectively enhance the chondrogenic ability of BMSCs inhibited by TNF-α in 3Ds (Fig. 3A–C). Moreover, the results also indicated that the differentiation of cells induced by 3D culture was more complete than that induced by the plate-based culture (Figs. 1C and 3B).
Mmp, belonging to the homologous zinc-dependent endopeptidase family, play a crucial role in cartilage degradation and the progression of OA. Degradation of type II collagen fibrin mediated by Mmp is considered to be an irreversible part of the pathological process of OA [27]. OA-related inflammatory factors can upregulate chondrocyte matrix degradation enzymes such as Mmp-1, Mmp-3, and Mmp-13 [28,29]. Our study found that SP600125 can inhibit the high expression of Mmp-1, Mmp-3, and Mmp-13 induced by TNF-α, suggesting that SP600125 may promote chondrogenic differentiation of BMSCs by inhibiting the inflammatory response (Fig. 3D, E).
Finally, the DMM mice model, a frequently used OA model strongly associated with clinical process [30], was employed to explore the anti-OA effect of BMSCs pretreated with SP600125 in vivo. In this study, we first demonstrated that SP600125 effectively enhanced the anti-OA effect of BMSCs (Fig. 4), which was indicated by increased Col2, lower expression of Mmp-13, alleviated cartilage erosion, improved cartilage superficial destruction, and decreased OARSI scores following the pretreatment of BMSCs with SP600125 when compared to those treated with BMSCs only in DMM model. In addition, the results also showed that inflammation was the main factor to suppress the therapeutic effect of BMSCs on OA, and inhibiting the JNK signaling pathway in BMSCs activated by inflammation was a potential candidate for promoting the application of BMSCs on OA treatment.
Conclusion
In conclusion, our results indicated that SP600125 could enhance the chondrogenic differentiation of BMSCs treated with TNF-α by inhibiting inflammation, and improve the protective effects of BMSCs on OA in DMM mice. This study is a deeper exploration of cartilage repair under inflammatory conditions on the basis of our previous studies, which provides a solid theoretical basis for the treatment of OA.
Footnotes
Acknowledgment
We would like to thank Elsevier for English language editing (Serial number: LE-203730-140778746056).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study is supported by the National Undergraduate Innovation Program (Grant No. S202010555068), the Scientific research project of Hunan Education Department (Grant Nos. 20B506 and 18B272), the Research project of University of South China (Grant No. 190XQD026), and the Natural Science Foundation for Young Scientists of Hunan Province (Grant No. 2021JJ40478).