
Abstract
Induced pluripotent stem cells (iPSCs) can serve as a biological resource for functional and conservation research for various species. This realization has led to the generation of iPSCs from many species, including those identified as endangered. However, the understanding of species variation in mammalian iPSCs remains largely unknown. To gain insight into species variation in iPSCs, we generated iPSCs from a new species Grevy's zebra (Equus grevyi; gz-iPSCs), which has been listed as endangered in the IUCN (International Union for Conservation of Nature) Red List. We isolated primary fibroblast cells from an individual and successfully reprogrammed them into iPSCs. The generated gz-iPSCs continued to grow under primed-type culture condition and showed pluripotency and differentiation potential. To describe the molecular characteristics of gz-iPSCs, we performed RNA sequencing analysis. The gz-iPSC transcriptome showed robust expression of pluripotency-associated genes reported in human and mouse, suggesting evolutionary conservation among the species. This study provides insight into the iPSCs from a rare species and helps the understanding of the gene expression basis underlying mammalian pluripotent stem cells.
Introduction
Mammalian induced pluripotent stem cells (iPSCs), which show unlimited self-renewal and the ability to differentiate into cells from any of the three germ layers, can be potential sources of differentiated tissue cells for fundamental research and conservation of diverse species, especially endangered species. In general, biological materials of nonmodel mammals are constrained because of ethical and technical concerns, and the potential properties of pluripotent stem cells (PSCs) allow for an increase in resources for functional studies and assisted reproductive technologies [1] for these species. The development of iPSC technology [2,3] has broadened the opportunity to study PSCs across a range of mammalian species.
PSCs exhibit both similarities and differences in their characteristics across species, highlighting the importance of understanding PSCs from various species. Derivations of PSCs have been reported in a range of species, including cows [4], pigs [5], horses [6 –10], naked mole-rats [11], and other mammalian species [12 –18]. In most studies, PSCs have been shown to satisfy many of the criteria for pluripotency, although some of the characteristics of the cells may differ among species [12,14]. Biological processes and associated genes that play crucial roles in PSCs have been extensively studied in humans and mice [2,19 –21], but molecular characteristics underlying mammalian PSCs have been poorly explored in nonmodel species.
The transcriptome approach is a powerful tool for elucidating the molecular characteristics of species cells [22]. While we previously described the evolutionary pattern in the pluripotency gene regulatory network based on changes in protein-coding genes [23], examining changes in gene expression may enable further insights into the phenotypic differences and similarities between species [22]. Gene expression comparison has previously highlighted the common regulation of signaling pathways in primates and mice [24], and the transcriptome of a new species will enable a better understanding of the molecular characteristics of mammalian PSCs.
Studies of PSCs from domestic animals, including cows [4] and pigs [5], have been reported, while research on Perissodactyla PSCs has been limited to horses [6 –10] and the northern white rhinoceros [13,15]. Grevy's zebra (Equus grevyi) belongs to the family Equidae, a taxonomic group including horses, donkeys, and zebras. It is one of three extant zebra species and the largest living wild equid. Grevy's zebra has experienced a serious population decline of 54% over the last 30 years, leaving ∼2,600 individuals [25] with decreasing genetic diversity [26]. It has been listed in CITES appendix I and is classified as “Endangered” on the IUCN (International Union for Conservation of Nature) Red List [25]. Studying iPSCs from Grevy's zebra may aid conservation efforts and will also provide new insights into mammalian PSCs.
Here, we report the first generation of iPSCs from Grevy's zebra (gz-iPSCs). We generated gz-iPSCs that exhibited a morphology of flat colonies and could be maintained under primed-type culture conditions. gz-iPSCs expressed pluripotency markers and exhibited differentiation abilities. To understand the molecular characteristics of the generated gz-iPSCs, we performed RNA sequencing (RNA-seq). This study provides insights into the variations in mammalian PSCs and contributes to the future conservation management of this endangered species.
Materials and Methods
General methods
A full description of the Supplementary Methods can be found in the Supplementary Data.
Generation of gz-iPSCs
This study was conducted in strict accordance with the guidelines for the ethics of animal research by Kyoto University and the Wildlife Research Centre of Kyoto University (WRC-2021-0016A). The sampling and methods were approved by the Kyoto City Zoo and Wildlife Research Centre of Kyoto University. Skin tissue samples were obtained from a female Grevy's zebra that died of natural causes at the Kyoto City Zoo (Japan). Primary fibroblasts were isolated and cultured from the skin tissue as previously described [27].
Cellular reprogramming was conducted using retrovirus vectors, as previously described [2,3]. Briefly, retroviral vectors were prepared using human POU5F1, SOX2, KLF4, and c-MYC [2]. Two independent reprogramming experiments were performed, and the reprogramming conditions tested in this study are summarized in Supplementary Table S1. The established gz-iPSCs were tested to determine whether they met the defining criteria for mammalian PSCs (Supplementary Data).
Cell culture
gz-iPSCs were cultured in mTeSR-1 (STEMCELL Technologies, Vancouver, Canada) [28] with 10 μM Rho-associated coiled-coil forming kinase (ROCK) inhibitor Y-27632 (Fujifilm Wako Pure Chemical Corporation, Osaka, Japan) on dishes coated with Matrigel (Corning, Corning, NY). The cells were cultured in a humidified incubator at 37°C with 5% (v/v) CO2. Cellular samples were tested for mycoplasma infection using the MycoAlert Mycoplasma Detection Kit (Lonza, Basel, Switzerland) according to the manufacturer's protocol.
RNA-seq analysis
Total RNA from gz-iPSCs (n PSC = 3; clone A; passages 16, 25, and 31) and fibroblasts (n fib = 3; passage 3, 6, 7) was used for RNA-seq. Isolated RNA was quantified using a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific) and a Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA). Total RNA (40 ng) was used for library preparation using Oxford Nanopore Technologies (ONT, Oxford, United Kingdom) long-read complementary DNA (cDNA) sequencing. cDNA was generated using the polymerase chain reaction (PCR)-cDNA Barcoding kit (SQK-PCB109; ONT), according to the manufacturer's protocol. For sequencing, libraries were applied to a single Nanopore Flow Cell (v 9.4.1) with MinION sequencer and run for 72 h.
Sequenced reads were base-called and demultiplexed using ONT EPI2ME software. Adapter sequences were trimmed from the reads using Porechop (v. 0.2.4) [29]. Low-quality reads were filtered using NanoFilt in NanoPack (v. 2.7.1) [30]. High-quality genome assembly and annotation are lacking in zebras, and therefore, horse (Equus caballus, EquCab3.0) was used as a reference for assembly. The filtered reads were mapped to the horse transcriptome using Minimap2 (v. 2.17) [31]. The mapped reads were counted using Salmon software (v. 1.3.0) [32]. Transcript reads were converted to gene-level abundance using tximport (v. 3.13) [33] and annotated with human orthologs using the BioMart tool in Ensembl (
Differentially expressed genes (DEGs) were identified using DESeq2 (v. 1.28.1) [35] with a false discovery rate (FDR)-adjusted P value of 0.1 [36], and | log2FoldChange | > 1 as default.
A volcano plot was constructed using EnhancedVolcano (v. 1.6.0), in which log2FoldChange values were shrunk using the apeglm method in DESeq2 [37], and an FDR lower than 10−20 was compressed for visualization. Protein analysis through evolutionary relationships (PANTHER) provided by the Gene Ontology (GO) Consortium (
Results
Generation of Grevy's zebra iPSCs
As the source for gz-iPSCs, we used primary fibroblasts from the skin tissue of an adult female Grevy's zebra (Fig. 1A, B). To reprogram somatic cells, we transduced gz-fibroblasts at passage 3 with retroviral vectors (Fig. 1C and Supplementary Fig. S1). We observed the formation of PSC-like colonies on day 11 and picked colonies on day 17 (Supplementary Fig. S2 and Supplementary Table S1).
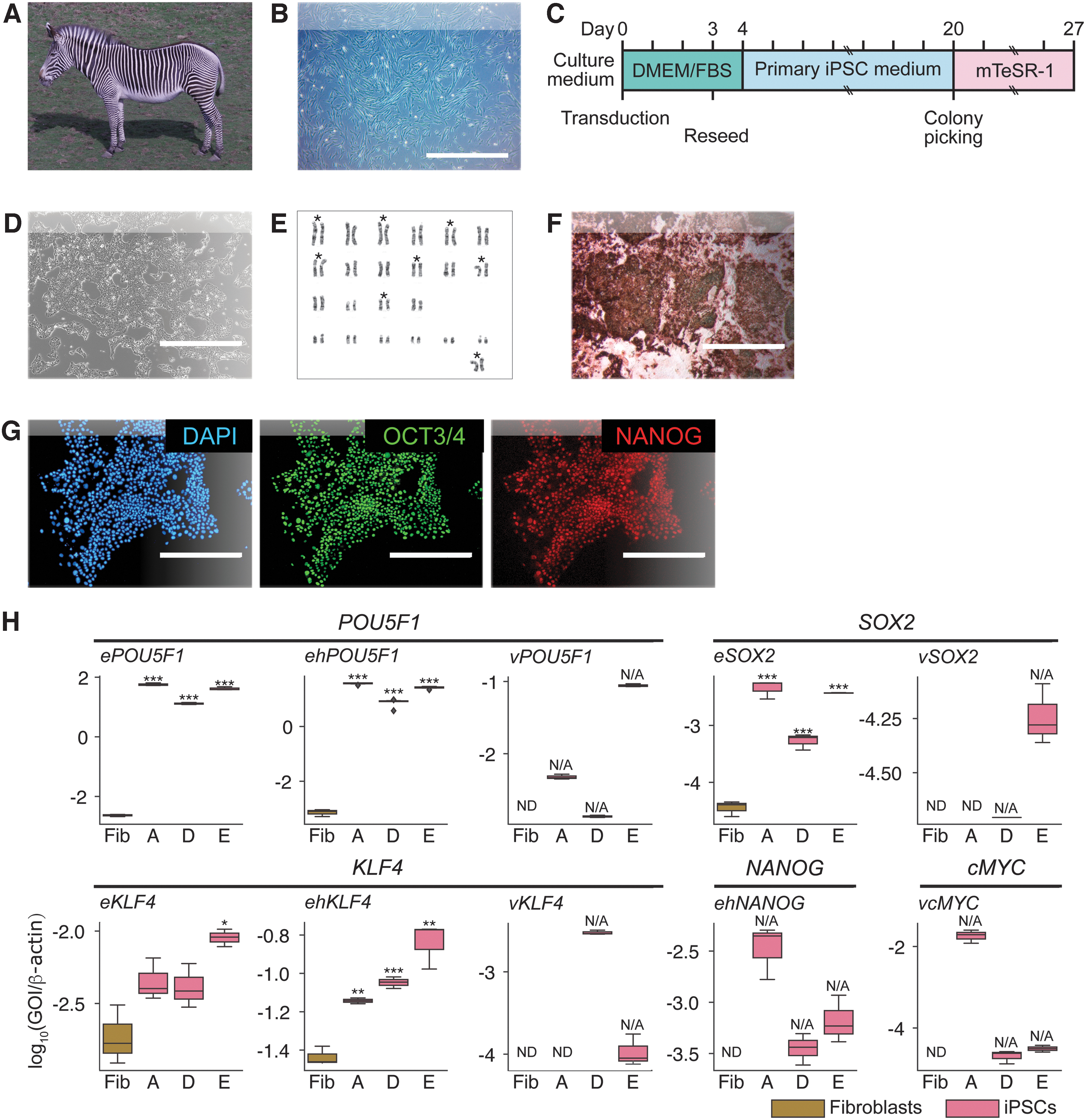
Generation of Grevy's zebra iPSCs from primary fibroblasts.
To optimize culture conditions, we explored culture media and the addition of small molecules and found that gz-iPSCs grew most stably with feeder-free and primed-type conditions of mTeSR-1 [28] with ROCK inhibitor [41] (Supplementary Fig. S3). To select the primary clones for continuous culture and subsequent analyses, we carried out preliminary pluripotency assessment experiments. We identified three primary clones (named A, D, and E) based on alkaline phosphatase (AP) activity, gene expression of pluripotency markers, and the silencing of viral markers (Supplementary Fig. S4).
To determine whether the chosen colonies exhibited the defining features of mammalian PSCs, we investigated the cellular characteristics of the putative gz-iPSCs. The morphology of the gz-iPSCs resembled human primed-type PSC colonies, which form a monolayer of cells with clear colony edges [2] (Fig. 1D). During passages 24–28, gz-iPSCs showed doubling times of 22.6 ± 2.4 h, similar to that of human embryonic stem cells (ESCs) [42].
The karyotype showed that 68% of clone A cells had a normal number of chromosomes at passage 13, however, 32% of them had one extra chromosome (Fig. 1E). In addition, the chromosomes with a normal number shows abnormalities in their length and banding patterns, and we could arrange a limited number of chromosomes with confidence for their pairing and position, in comparison with the reference karyotype of the Grevy's zebra [43].
The MycoAlert test showed that all three clones were negative for mycoplasma (Supplementary Fig. S5). To date, these gz-iPSC lines have been maintained for more than 30 passages.
To evaluate the expression of pluripotency markers, we conducted molecular analysis at the protein and genetic levels. Protein and gene expression analysis showed that the generated gz-iPSCs expressed pluripotency markers (Fig. 1F–H and Supplementary Figs. S4A–E and S6). To determine whether the expressed genes were endogenous or exogenous, we designed equine-specific primers; ePOU5F1, eSOX2, and eKLF4 (primers used in this study are listed in Supplementary Table S2). We also designed multispecies-specific primers to react with both equine and human genes, named ehPOU5F1, ehKLF4, and ehNANOG, to confirm the expression of these markers.
We observed higher expression levels of every analyzed pluripotency marker in iPSC samples compared with fibroblasts (Supplementary Table S3), using both equine-specific and multispecies-specific primers at passage 25. The expression of virally transduced genes was not completely silenced but observed at low levels. In addition, the expression levels of endogenous genes were much higher than those of the exogenous genes (Fig. 1H). Importantly, both protein and gene expression analyses indicated expression of NANOG, which was not introduced with the reprogramming vectors or not observed in the original gz-fibroblasts. These results indicate that endogenous pluripotency markers were induced by reprogramming, and that the generated gz-iPSCs were able to maintain their status by producing their own pluripotency molecules.
To examine the differentiation potential of gz-iPSCs, we conducted embryoid body (EB) formation and observed that gz-iPSCs grew in ball-like aggregates (Fig. 2A), indicating the formation of EBs. Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) analysis revealed increased expression of lineage markers for the three germ layers, including ectoderm (NES, TUBB3, and PAX6), mesoderm (SMA and BMP4), and endoderm (AFP, GATA4, SOX17, and CXCR4) [44] (Fig. 2B). In summary, these results show the differentiation capability of gz-iPSCs and indicate that gz-fibroblasts were successfully reprogrammed into iPSCs.
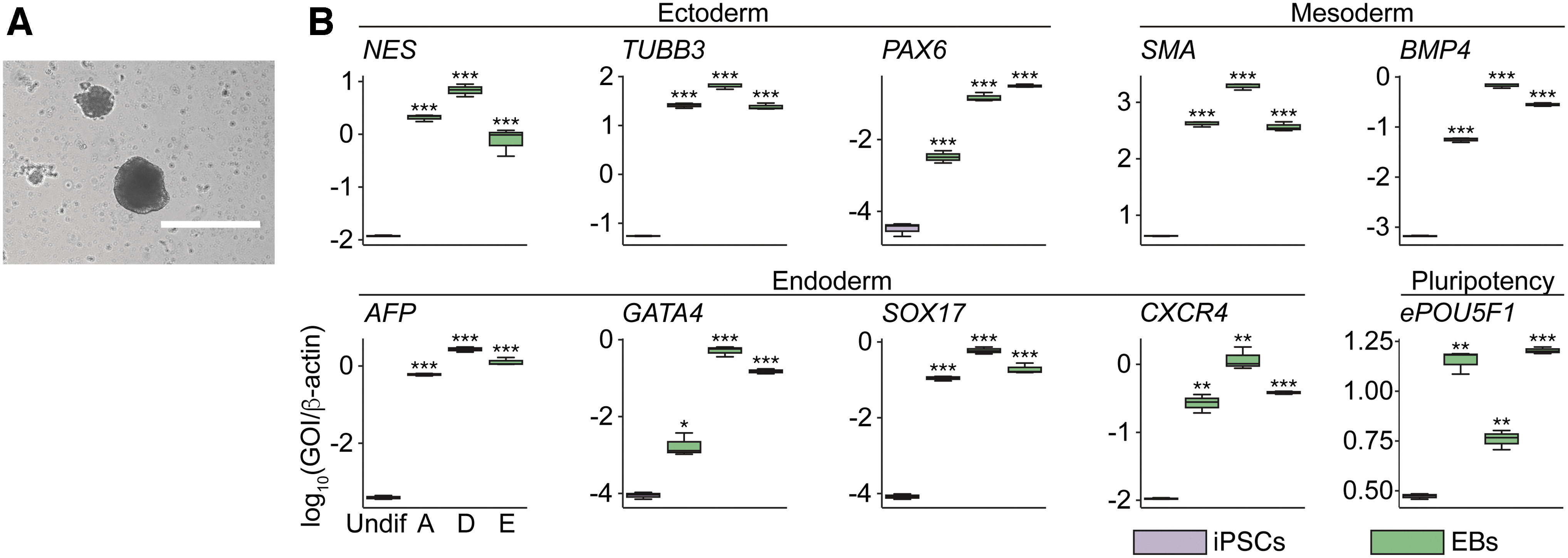
In vitro EB formation for the differentiation capability of gz-iPSCs.
Identification of genes altered by the generation of gz-iPSCs
To investigate the comprehensive changes in gene expression, we performed RNA-seq and analyzed the DEGs between gz-iPSCs and fibroblasts. Our RNA samples showed RNA integrity number (RIN) values of 10, 9.9, and 10 (gz-iPSCs; passages 16, 25, and 31) and 8.4, 9.8, and 10 (gz-fibroblasts; passages 3, 6, and 7). RNA-seq yielded 462,723–1,418,369 reads, and 92.32%–95.72% of the reads were successfully mapped (Supplementary Table S4). DEG analysis revealed 1,144 upregulated and 1,495 downregulated DEGs with adjusted P values FDR <0.1 and | log2FoldChange | > 1 (Fig. 3A and Supplementary Table S5).

DEG analysis of Grevy's zebra fibroblasts and iPSC transcriptome.
As expected, the upregulated genes included well-known pluripotency genes, such as POU5F1, DNMT3B, SALL4, ZFP42 (also known as REX1), and LIN28 [45], which were highly expressed compared with the fibroblasts. In contrast, the downregulated genes included the fibroblast genes VIM, DDR2, TGFBR2, COL1A1, COL1A2, and FSP1 (also known as S100A4) [46].
To characterize the derived gz-iPSCs for representative biological functions, we performed GO enrichment analysis (Fig. 3B and Supplementary Tables S6 and S7). GO analysis revealed enrichment of terms related to pluripotency such as embryo development and telomere maintenance. GO terms associated with metabolism and viral defence were consistent with those of human iPSCs [47,48].
To gain insight into the molecular basis underlying the enriched biological processes, we compared the transcripts per million (TPM) of the DEGs (Supplementary Fig. S7). We found upregulation of further pluripotency signature genes, including EPCAM and DPPA3.
To further evaluate the expression changes in DEGs, we inspected log2FoldChanges >4 and found the highly upregulated DEGs included EPCAM, ESRP1, and CDH1 (also known as E-cadherin) (Supplementary Table S5). In addition, highly upregulated DEGs included markers of pluripotency state and germ line cells, including GATA3, VGLL1, TFAP2A, TFAP2C, and SOX15. Together, our RNA-seq analysis revealed that the gene expression profile of reprogrammed gz-iPSCs is different from that of gz-fibroblasts and shows molecular characteristics associated with mammalian PSCs, thus supporting the successful generation of gz-iPSCs.
Discussion
In this study, we report the generation of iPSCs from an endangered species, Grevy's zebra. Primary gz-fibroblasts were successfully reprogrammed into gz-iPSCs. The gz-iPSCs generated in this study exhibited PSC characteristics in terms of morphology, expression of pluripotency markers, and differentiation potential into three germ layers.
Using the RNA-seq results, we elucidated the molecular basis regulating the pluripotency characteristics of gz-iPSCs. Similarly to ESCs, iPSCs can differentiate into three germ layers, maintain high telomerase activity, and exhibit proliferative potential [49]. The observed GO enrichment terms indicates that the derived gz-iPSCs have acquired the general characteristics of PSCs, as also shown by EB differentiation experiments and cell doubling time similar to that of human iPSCs. We observed the expression of well-known pluripotency-associated genes, including the core pluripotency transcription factor POU5F1 [50,51], DNA methyltransferase DNMT3B [52], RNA-binding protein ESRP1 [53], and cell adhesion molecules EPCAM [54] and CDH1 [55,56], suggesting that these genes play important roles in gz-iPSCs as in human PSCs.
Whereas iPSCs have been generated from highly endangered species previously [13,15,18], the number of reports is still limited. When an anthrax outbreak occurred and killed 53 Grevy's zebras in Kenya, uncertainty about the possible adverse effects of vaccinating Grevy's zebras against anthrax impeded the immediate application of medical treatment [57]. Safe and effective disease control protocols are especially important for species with declining populations, and iPSCs from endangered species could enable the development of therapeutic applications [1].
One potential iPSC technology relevant for conservation management is genetic rescue [58], and the generation of iPSCs from endangered species provides resources for in vitro derivation of germ line materials. In this study, we applied retroviral vectors that enable high reprogramming efficiency but involve transgene integration into a host genome, and successfully derived three clones of gz-iPSCs. Transgene-free reprogramming methods have recently been reported to be effective across a range of taxonomic groups [59] although our integration-free method did not lead to a formation of iPSC-like colonies (data not shown). The chromosomal abnormalities observed in this study may indicate that our culture condition is not ideal for gz-iPSCs.
The findings of this study will guide future studies that will be critical to determine improved reprogramming and culture conditions of iPSCs of this species. It also should be noted that the RNA-seq data we reported were generated from a single cell line. Given the potential variations in gene expression between cell lines [60], generation of iPSCs from multiple individuals of a species will be necessary for comprehensive understandings of gz-iPSC gene expression; however, biological materials are often limited in endangered species due to technical and ethical concern. In this regard and to mitigate the decline of genetic diversity in Grevy's zebra [26], cryopreservation of fibroblasts and the subsequent generation of iPSCs from extant individuals will be important for further understandings of gz-iPSCs and future conservation efforts [1].
Conclusions
The iPSCs derived from Grevy's zebra reported in this study have advanced our understanding of mammalian PSCs. The effective reprogramming of gz-fibroblasts by human transcription factors supports the conservation of reprogramming mechanisms. The transcriptome of gz-iPSCs allowed us to further characterize the molecular basis of these newly established iPSCs. gz-iPSCs will provide resources for future functional studies and the conservation management of this endangered species.
Data Availability
RNA-seq data have been deposited under BioProject PRJNA748892 and GEO GSE180619.
Footnotes
Acknowledgments
The authors thank Oliver A. Ryder and Marisa L. Korody for constructive feedback on the article, N. Yoshida for providing technical support for the experiments, Marlys L. Houck and Ann C. Misuraca for assisting with karyotyping.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was supported by Japan Society for the Promotion of Science (JSPS) KAKENHI [grant numbers 17H03624, 20H00420 (M.I.-M.), and 17H02083 (K.K.)], the Kyoto University Supporting Program for Interaction-based Initiative Team Studies (SPIRITS; M.I.-M.), and the Environment Research and Technology Development Fund of the Environmental Restoration and Conservation Agency of Japan [grant number FPMEERF20214001 (M.I.-M.)].
Supplementary Material
Supplementary Data
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.