
Abstract
Alcohol consumption is regarded as one of the leading risk factors for secondary osteopenia. Angiogenesis and osteogenesis coupled by type-H vessels coordinate the biological process of bone homeostasis to prevent osteopenia. This study aimed to determine whether chronic alcohol inhibits type-H vessel-dependent bone formation. Two-month-old mice were fed with 5% (v/v) alcohol liquid diet (28% of calories) or normal liquid diet every day for 2 months. The tibias were isolated and detected with X-ray and microcomputed tomography. Paraffin-embedded or frozen tibial sections were prepared and used for immunohistochemical or immunofluorescence staining, respectively. Human umbilical vein endothelial cells (HUVECs) were treated with different concentrations of alcohol, including 0 mM (0%), 8.7 mM (0.5%), 52 mM (3%), or 87 mM (5%) alcohol for 12 h. The conditioned medium of the above HUVEC cells was collected to culture human bone marrow-mesenchymal stem cells (BM-MSCs), which were induced to differentiate into osteoblasts in vitro. The alcoholic diet retarded the bone growth and led to osteoporosis, impaired bone formation of osteoblasts, and decreased CD31hiEMCNhi type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells in mice. Alcohol treatment obviously increased the expression of p16, while significantly decreased the expression of Bmi-1, CDK6, Cyclin D, E2F1, and bone morphogenetic protein (BMP)2 compared with vehicle. Alcohol inhibited the differentiation of BM-MSCs into osteoblasts through reducing the BMP2 secretion of endothelial cells in type-H vessels. Alcoholic diet impaired CD31hiEMCNhi type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells through Bmi-1/p16 signaling, and inhibited the differentiation of BM-MSCs into osteoblasts through reducing the BMP2 secretion of endothelial cells in type-H vessels. This study provides a basis for developing a new treatment strategy targeting aging endothelial cells of type-H vessel to prevent alcoholic osteopenia.
Introduction
Osteoporosis is a metabolic disease, which is characterized by decreased bone mineral density (BMD), destruction of bone microstructure, and increased bone fragility and risk of fracture [1]. Chronic consumption of excessive alcohol eventually results in an osteopenic skeleton and increased risk for osteoporosis. Alcohol is one of the leading risk factors for male osteoporosis [2]. Compared with individuals without alcoholism, those with chronic alcohol abuse have significantly lower BMD and lower bone volume in the femoral neck, showing thinner trabeculae, reduced range of bone-like surface, and decreased average wall thickness of trabeculae [3,4].
Blood vessels act as structural templates for bone development and also aggregate key elements of bone homeostasis into the osteogenic microenvironment, including minerals, growth factors, and osteogenic progenitor cells [5]. Recent studies revealed that type-H vessels with strong positive expression of CD31 and endomucin (EMCN) proteins coupled angiogenesis and osteogenesis [6]. Type-H vessels are surrounded by the transcription factors Runx2-positive and Osterix-positive osteoprogenitors, which determines that there is a close interaction between type-H vessels and osteogenic progenitor cells. Type-H vessels play an effective role in promoting bone regeneration. The osteogenic progenitor cells around type-H vessels express Runx2 and Osterix at high levels to promote bone formation. Moreover, endothelial cells of type-H vessels secrete many factors that trigger the proliferation and differentiation of osteoprogenitors to promote osteogenesis [7,8]. Alcohol has been reported to have a negative effect on angiogenesis [9,10].
The antiangiogenic effect of alcohol was found associated with excess reactive oxygen species production [11]. However, whether alcohol hampers proliferation and promotes aging of endothelial cells of type-H vessels is still unclear, and whether alcohol inhibits the differentiation of bone marrow-mesenchymal stem cells (BM-MSCs) into osteoblasts through reducing the secretion of bone morphogenetic protein (BMP) of endothelial cells in type-H vessels remains unknown.
B lymphoma Mo-MLV insertion region 1 (Bmi-1) is a member of the polycomb family of transcriptional repressors [12,13]. It could regulate cell cycle progression and prevent cell senescence through inhibiting p16INK4a (hereafter p16)/Rb and p19AFR/p53 pathways [14,15]. Moreover, it also maintains mitochondrial function and redox homeostasis [13,14]. In previous reports, inhibiting the expression of miR-125a could decrease the expression of p16, lead to the proliferation of vascular smooth muscle and endothelial cells, and then promote the remodeling of pulmonary artery [16]. However, whether Bmi-1 promotes proliferation and prevents aging of type-H vessels by inhibiting p16 and maintaining cell cycle progression is unclear.
In this study, we demonstrated the alcoholic diet impaired CD31hiEMCNhi type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells through Bmi-1/p16 signaling, and inhibited the differentiation of BM-MSCs into osteoblasts through reducing the BMP2 secretion of endothelial cells in type-H vessels. This study provides a basis for developing a new treatment strategy targeting aging endothelial cells of type-H vessel to prevent alcoholic osteopenia.
Materials and Methods
Mice and experimental design
Eight-week-old C57BL/6J male mice were obtained from Vital River laboratories in Beijing of China and randomly assigned to two weight-matched groups (n = 6/group), including an alcohol Lieber–DeCarli liquid diet (TROPHIC Animal Feed High-tech Co., Ltd., Nantong, Jiangsu, China) feeding group and a corresponding paired feeding group. By substituting carbohydrate calories with alcohol calories, alcohol was added to the Lieber–DeCarli liquid diet [17]. This alcohol liquid diet provides 28% of the calories and is equivalent to 5% alcohol by volume. The paired feeding group received the Lieber–DeCarli control liquid diet (TROPHIC Animal Feed High-tech Co., Ltd.). Based on the previous day's dietary consumption, the paired feeding group was given equal calorie to its corresponding alcohol group. Alcohol and paired feeding diets were administered for 2 months. According to the previous reports [18,19] and feed instructions, the final blood concentrations of mice can reach 100 to 150 mg%, which can meet the need to simulate osteopenia caused by chronic drinking.
This study was carried out in strict accordance with the guidelines of the Institute for Laboratory Animal Research of Nanjing Medical University in Nanjing of China. The protocol was approved by the Committee on the Ethics of Animal Experiments of Nanjing Medical University (Permit Number: IACUC-1706001).
Radiography and microcomputed tomography
For the alcohol-treated groups, alcohol was added to the Lieber–DeCarli diet by substituting carbohydrate calories with alcohol calories. The alcohol concentration in the diet was increased slowly in a stepwise manner until 28% total calories were reached, which constituted final alcohol concentrations of 5% (v/v), respectively, and maintained 5% (v/v) for 2 months until sacrifice. Four-month-old alcohol and pair-fed mice were euthanized to obtain the tibial tissue samples. Radiography was performed with a Faxitron machine (Model 805; Faxitron Bioptics, LLC, Tucson, AZ) to observe tibias. A Lunar PIXImus densitometer (GE Lunar, Madison, WI) was used to measure BMD of tibias. A SkyScan 1272 scanner and associated analysis software (Bruker, Kontich, Belgium) were used to conduct microcomputed tomography (micro-CT) of tibias as described [20] at the Nanjing Medical University Medical Laboratory Animal Center.
Histology
Mouse tibias were fixed in 4% periodate-lysine-paraformaldehyde (PLP) [14,15] for 24 h and decalcified in a 10% ethylenediaminetetraacetic acid for 3–4 weeks. Samples were then embedded in paraffin, sectioned at 5 μm in thickness with a rotary microtome (Leica Microsystems Nussloch GmbH, Nubloch, Germany), and processed for histochemical staining of total collagen as previously described [15,20] and Safranin O/Fast Green staining using a kit (No. G1371; Beijing Solarbio Science & Technology Co., Ltd., Beijing, China), or for immunohistochemical staining [20,21]. The samples were embedded in Optimal Cutting Temperature (O.C.T.) compound (No. 4583; SAKURA Finetek USA, Inc., Torrance, CA), and were sliced in 30 μm thickness with a freezing microtome (Thermo Scientific Cryotome FSE Cryostats, Loughborough, Leicestershire) for immunofluorescence staining.
Human BM-MSCs and human umbilical vein endothelial cell culture
Human bone marrow-mesenchymal stem cells (hBM-MSCs) were obtained from the American Type Culture Collection (PTA-1058; ATCC, Manassas, VA). The hBM-MSCs were cultured in a medium, including 85% (v/v) α-MEM, 15% (v/v) fetal bovine serum (FBS) (Gibco, Grand Island, NY), and supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin (Gibco). Human umbilical vein endothelial cells (HUVECs) were obtained from the China Infrastructure of Cell Line Resources (Beijing, China) and cultured in endothelial cell medium (No. 1001; ScienCell Research Laboratories, Inc., Carlsbad, CA) containing 10% FBS, Endothelial Cell Growth Supplement (No. 1052; ScienCell Research Laboratories, Inc.), 100 U/mL penicillin, and 100 μg/mL streptomycin as previously described [22]. Cells from the 3rd passage were used in the experiment, and the culture medium was changed every 3 days. We used the Cell Counting Kit-8 (CCK-8) assay to assess cell viabilities of HUVECs and hBM-MSCs, and found them healthy (Supplementary Fig. S1A, B).
Cell modeling
Some lines of evidence have already used HUVECs as a type-H vessel model as previously described [23 –25]. For the experimental alcohol concentration in vitro, different concentrations of alcohol, including 0 mM (0%), 8.7 mM (0.5%), 52 mM (3%), or 87 mM (5%) alcohol were used to detect the effect of alcohol on endothelial cell proliferation and senescence, and BMP2 secretion in a dose-dependent manner. The selection of these concentrations also refers to the previous study [26], which can simulate the effect of alcohol on vascular endothelium in vitro.
Immunofluorescence staining
Induced HUVECs were fixed with PLP solution [12] for 15 min and preincubated with serum. Primary antibodies against platelet endothelial cell adhesion molecule-1 (PECAM-1) (No. 557355; BD Pharmingen, Franklin Lakes, NJ), EMCN (No. DF13357; Affinity, Melbourne, Australia), Osterix (sc-393325; Santa Cruz Biotechnology, Inc., Dallas, TX), Runx2 (sc-390351; Santa Cruz Biotechnology, Inc.), p16 (ab211542; Abcam, Cambridge, MA), BMP2 (18933-1-ap; ProteinTech Group, Inc., Rosemont, IL), and Bmi-1 (66161-1-lg; ProteinTech Group, Inc.) were used. After primary antibody incubation, bone sections and cells were washed with phosphate-buffered saline (0.01 mM PO4 3−, pH 7.4) for three times and incubated with appropriate Alexa Fluor-coupled secondary antibodies (Life Technologies Corporation, USA). Nuclei were labeled by 4′,6-diamidino-2-phenylindole (Sigma-Aldrich, St. Louis, MO) and mounted with medium, which prevented quenching of fluorescence (Vector Laboratories Inc., Burlingame, CA). No primary antibody or isotype controls was shown in Supplementary Fig. S1D.
Immunocytochemical staining
Bone sections were air dried and endogenous peroxidase was blocked with 3% H2O2 and cells were preincubated with serum. Primary antibody against proliferating cell nuclear antigen (PCNA) (sc-130920; Santa Cruz Biotechnology, Inc.) and type-1 collagen (collagen 1) (ab88147; Abcam) was used. After washing, bone sections were incubated with secondary antibody (biotinylated IgG; Sigma), washed, and processed using the VECTASTAIN ABC-HRP Kits (Vector Laboratories Inc.).
In vitro proliferation assays of HUVECs and hBM-MSCs
We performed a CCK-8 assay (No. C0038; Beyotime Institute of Biotechnology, Shanghai, China) to assess cell proliferation as previously described [12]. For the CCK-8 assay, 2 × 103 HUVECs or hBM-MSCs were inoculated into 96-well plates to characterize cell health. HUVEC's viability was evaluated at 0 h, 12 h, 24 h, 36 h, 48 h, or 60 h. hBM-MSCs' viability was evaluated at 0 h, 24 h, 48 h, 72 h, or 96 h. Two times 103 HUVECs were inoculated into 96-well plates and incubated with alcohol for 12 h. HUVEC's viability was evaluated at 0, 24, and 48 h after alcohol treatment. We have used the CCK-8 assay to assess cell viabilities of HUVECs and hBM-MSCs, and found them healthy (Supplementary Fig. S1A, B).
In vitro assays of HUVECs tube formation
We assayed the tube formation with HUVECs on Matrigel (BD Bioscience, San Jose, CA). Two times 104 HUVECs were incubated in a precooled 96-well plate and coated with 50 μL Matrigel. Cells were then treated with 8.7 mM (0.5%), 52 mM (3%), or 87 mM (5%) alcohol for 12 h, or with complete medium as the vehicle. After 12 h, endothelial cells linked by tubes were observed and quantified as previously described [27].
Osteogenic differentiation of hBM-MSCs in vitro
To identify osteogenic differentiation, the hBM-MSCs were cultured for 18 days in osteogenic medium of α-MEM from alcohol-treated HUVECs containing 15% (v/v) FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 2 mM L-glutamine, 10−8 M dexamethasone (Sigma-Aldrich), 50 μg/mL ascorbic acid, and 10 mM β-glycerophosphate (Sigma-Aldrich) in a humidified 5% CO2 incubator at 37°C [28,29]. Medium was changed every 3 days. Induced hBM-MSCs were stained sequentially for alkaline phosphatase (ALP) by hydrolysis of naphthol phosphate, calcium nodules with Alizarin Red S [28,29].
Cytology staining
ALP staining
Induced hBM-MSCs were incubated in dark for 30 mins at room temperature in 100 mM Tris-maleate buffer containing naphthol AS-MX phosphate (0.2 mg/mL; Sigma-Aldrich) dissolved in ethylene glycol monomethyl ether (Sigma-Aldrich) as a substrate and Fast Red TR (0.4 mg/mL; Sigma-Aldrich) as a stain for the reaction product [27,30]. The hBM-MSCs were gently washed under running water and left to dry.
Alizarin Red S staining for calcium nodules
Induced hBM-MSCs were exposed to a solution of Alizarin Red S (pH 6.2; 1 mg/mL) for 30 min at room temperature, after which hBM-MSCs were gently washed under running water and left to dry [27,30].
Western blot
Western blot analysis of tibial bone marrow from the metaphyseal and HUVECs were performed following previously described methods [14,31]. Primary antibodies against BMP2 (18933-1-ap; ProteinTech Group, Inc.), Osterix (sc-393325; Santa Cruz Biotechnology, Inc.), Runx2 (sc-390351; Santa Cruz Biotechnology, Inc.), ALP (11187-1-ap; ProteinTech Group, Inc.), PECAM-1 (No. 557355; BD Pharmingen), EMCN (No. DF13357; Affinity), Bmi-1 (66161-1-lg; ProteinTech Group, Inc.), CDK6 (No. 3136S; Cell Signaling Technology, Inc., Boston, MA), Cyclin D (ab134175; Abcam), E2F1 (ab179445; Abcam), and β-actin (AP0714; Bioworld Technology, St. Louis Park, MN) were used.
Enzyme-linked immunosorbent assay
HUVECs were treated with 52 mM (3%) or 0 mM (0%) alcohol for 12 h. The supernatants of them were collected and filtered with MILLEX-GP 0.22-μm filters (Merck Millipore Ltd. Co., Cork, Munster, Ireland) to remove cell debris and concentrated to 10% volume with Amicon Ultra-4 centrifugal ultrafiltration tubes (nominal molecular weight limits 3 kDa) (Merck Millipore Ltd. Co.) as previously described [12,31 –33], and seeing as conditioned medium (CM). Concentrations of human-derived BMP2 (No. H00065) in CM, were detected using Enzyme-Linked Immunosorbent Assay (ELISA) Kits from Yifeixue Biotechnology in Nanjing of China as previously described [34].
Statistical analysis
GraphPad Prism software (version 9.0.0; GraphPad Software Inc., San Diego, CA) was used to statistically analyze as previously described [12,35]. Measurement data were described as mean ± standard error of the mean fold-change over control and analyzed by Student's t-test and one-way analysis of variance to compare differences among groups. Qualitative data were described as percentages and analyzed using chi-square tests as indicated [12,14,31]. P values were two sided and <0.05 was considered statistically significant.
Results
The alcoholic diet retards the bone growth and leads to osteopenia in mice
To determine the effect of alcoholic diet on bone growth, the changes of bone growth and development in normal- and alcohol-diet mice were compared and analyzed by X-ray photography and Safranin O/Fast Green staining. Results showed that the tibial size, length (Fig. 1A, B), and growth plate thickness (Fig. 1C, D) of alcohol-diet mice were decreased when compared with those of normal-diet mice. In addition, PCNA-positive chondrocytes were detected with immunohistochemistry and results showed that the proliferation of chondrocytes in alcohol-diet mice also decreased significantly in comparison with normal-diet mice (Fig. 1E, F). The changes of tibial bone volume and related indexes in mice fed with alcohol were observed by micro-CT (Fig. 1G) and total collagen staining (Fig. 1H). Compared with normal-diet mice, trabecular bone volume/bone tissue volume percentage and BMD were downregulated, while trabecular separation increased in alcohol-diet mice (Fig. 1I).

The alcoholic diet retards the bone growth and leads to osteopenia in mice. The experiments were carried out on the 2-month-old mice fed with normal diet or alcoholic diet for 2 months. The tibias were isolated and analyzed with X-ray and micro-CT. Paraffin-embedded tibial sections from 4-month-old normal- and alcohol-diet mice were prepared.
The alcoholic diet impairs osteogenic potential of osteoblasts in mice
To investigate whether the inhibition of alcoholic diet on bone mass in mice was related to the change of osteogenesis ability, Runx2 immunofluorescence staining of tibial metaphyseal and immunohistochemical staining of type-1-collagen (collagen 1) were conducted. Results showed that the positive staining of Runx2 and collagen 1 in alcohol-diet mice were significantly downregulated when compared with normal diet mice (Fig. 2A–D). At the same time, the relative expression of osteogenesis-related proteins, BMP2, Osterix, Runx2, and ALP, were detected by Western blots in tibialis lysate (Fig. 2E, F) to observe the effect of alcoholic diet on bone formation in mice. Compared with normal-diet mice, the protein levels of BMP2, Osterix, Runx2, and ALP were significantly decreased.
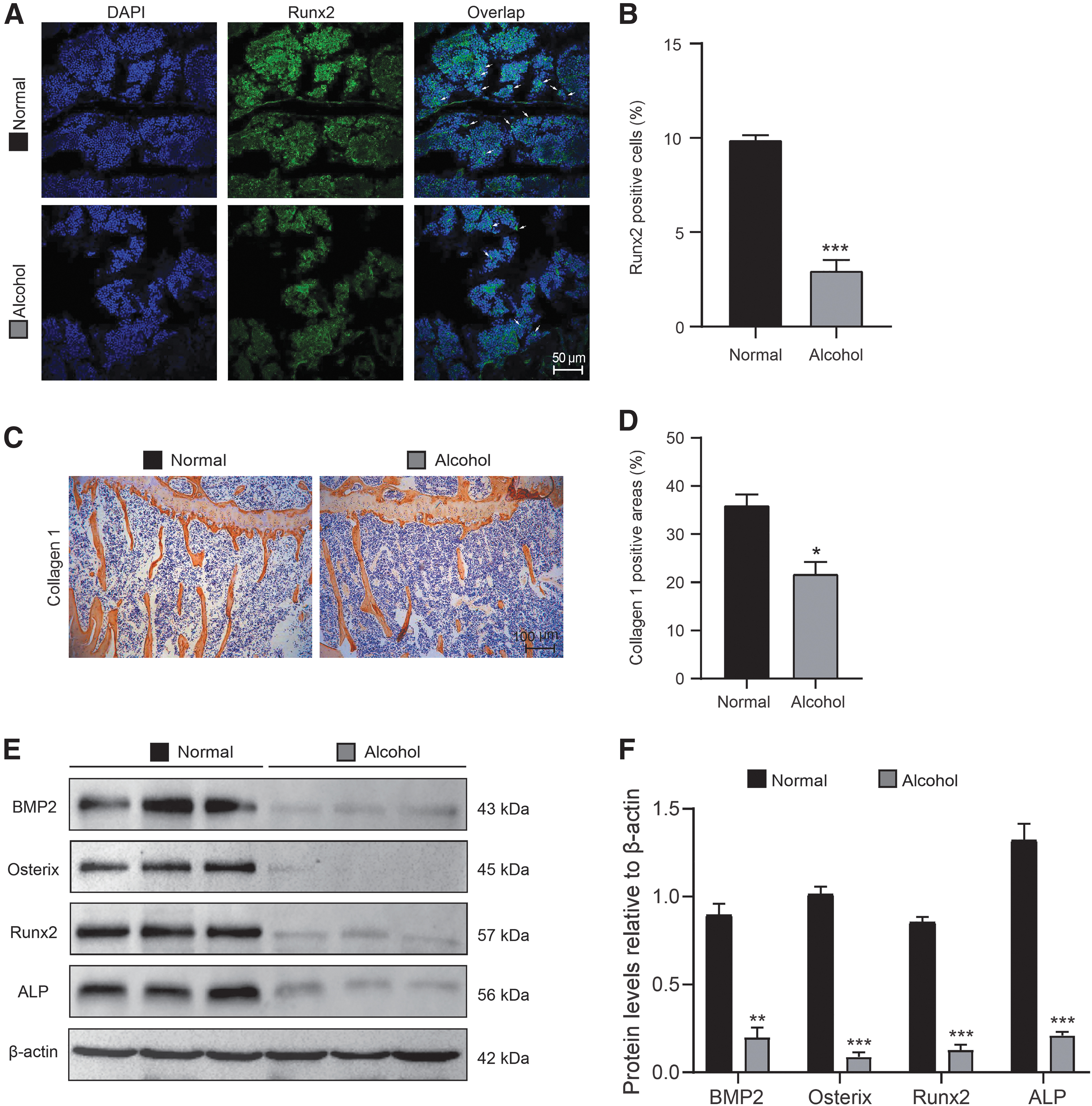
The alcoholic diet impairs osteogenic potential of osteoblasts in mice. The experiments were carried out on the 2-month-old mice fed with normal diet or alcoholic diet for 2 months. Paraffin-embedded or frozen tibial sections from 4-month-old normal- or alcohol-diet mice were prepared and used for immunohistochemical or immunofluorescence staining, respectively.
The alcoholic diet impairs CD31hiEMCNhi vascularization
To determine whether disturbance of bone formation of osteoblasts after feeding with alcoholic diet was associated with decrease of type-H vessels, immunofluorescence staining of CD31 and EMCN were conducted to observe type-H vessels. Compared with normal diet, alcohol diet decreased the number of CD31hiEMCNhi type-H vessels (Fig. 3A, B; Supplementary Fig. S1C). In addition, alcohol treatment significantly reduced the CD31 and EMCN expression and tube-forming ability of HUVECs. The dose-dependent inhibition of angiogenesis is obvious. After HUVECs were treated with or without alcohol for 12 h, compared with vehicle, 52 mM (3%) or 87 mM (5%) alcohol treatment significantly decreased the total tube length, and the expressions of EMCN and CD31; 87 mM (5%) alcohol treatment further decreased the total tube length and the expression of EMCN in comparison with 52 mM (3%) alcohol treatment (Fig. 3C–F).
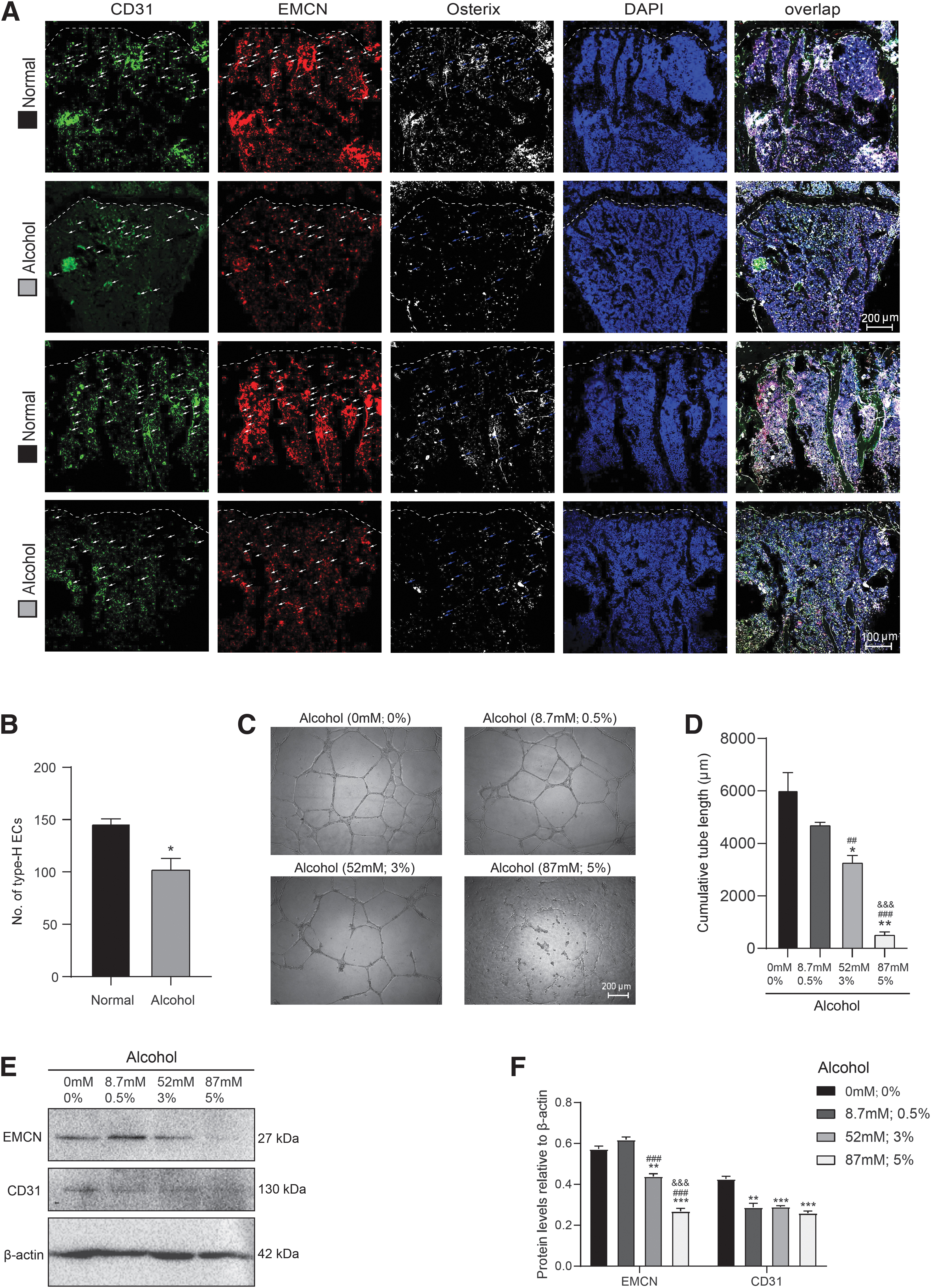
The alcoholic diet impairs CD31hiEMCNhi vascularization. The experiments were carried out on the 2-month-old mice fed with normal diet or alcoholic diet for 2 months. Frozen tibial sections from 4-month-old normal- and alcohol-diet mice were prepared and used for immunofluorescence staining.
The alcoholic diet impairs CD31hiEMCNhi type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells
To further investigate whether the reduced number of type-H vessels caused by alcohol treatment attributed to the decreased proliferation and increased aging of endothelial cells in type-H vessels, cell proliferation with CCK-8 assay, p16 expression with immunofluorescence staining, and proliferation-related proteins with Western blots of HUVECs were detected. After HUVECs were treated with or without alcohol for 12 h and then cultured for 48 h, 52 mM (3%) or 87 mM (5%) alcohol treatment significantly inhibited cell proliferation compared with vehicle; 87 mM (5%) alcohol treatment further inhibited cell proliferation in comparison with 52 mM (3%) alcohol treatment (Fig. 4A).

The alcoholic diet impairs CD31hiEMCNhi type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells. HUVECs were treated with 0 mM (0%), 8.7 mM (0.5%), 52 mM (3%), or 87 mM (5%) alcohol for 12 h.
After HUVECs were treated with or without alcohol for 12 h, in comparison with vehicle, 8.7 mM (0.5%) alcohol treatment increased the expression of p16, whereas decreased the expression of Bmi-1, Cyclin D, and E2F1; 52 mM (3%) or 87 mM (5%) alcohol treatment significantly increased the expression of p16, whereas significantly decreased the expression of Bmi-1, CDK6, Cyclin D, and E2F1; 87 mM (5%) alcohol treatment further increased the expression of p16, whereas decreased the expression of Bmi-1, CDK6, and Cyclin D compared with 52 mM (3%) alcohol treatment (Fig. 4B–E). In summary, alcohol impairs vascularization through inhibiting proliferation and promoting aging of endothelial cells.
Alcohol inhibits the differentiation of BM-MSCs into osteoblasts through impairing the endothelial cells
To investigate the effect of endothelial cells in type-H vessels on osteogenic differentiation of BM-MSCs, the CM of HUVEC cells treated with different concentrations of alcohol was collected to culture hBM-MSCs, which was then induced to differentiate into osteoblasts in vitro. Compared with vehicle, 52 mM (3%) or 87 mM (5%) alcohol treatment significantly decreased ALP and Alizarin Red S-positive calcium nodules; 87 mM (5%) alcohol treatment further decreased ALP and Alizarin Red S-positive calcium nodules (Fig. 5A–D). These results indicated that alcohol resulted in the dysbiosis of HUVECs' function, which hindered osteogenic differentiation of BM-MSCs.

Alcohol inhibits the differentiation of BM-MSCs into osteoblasts through impairing the endothelial cells. HUVECs were, respectively, treated with different concentrations of alcohol, including 0 mM (0%), 8.7 mM (0.5%), 52 mM (3%), or 87 mM (5%) alcohol for 12 h. The CM of the above HUVEC cells was collected to culture hBM-MSCs and induce them to differentiate into osteoblasts in vitro.
Alcohol reduces the BMP2 secretion of endothelial cells in type-H vessels
In vivo, alcohol-diet mice were characterized by fewer Osterix-positive osteoprogenitors and/or osteoblasts compared with normal-diet mice (Fig. 3A, B). To determine whether alcohol reduced the BMP2 secretion of endothelial cells in type-H vessels, the BMP2 expression of aging endothelial cells was observed with immunofluorescence staining and detected with Western blot. Results showed that alcohol-diet mice had decreased number of BMP2- or Bmi-1-positive endothelial cells and increased number of p16-positive endothelial cells in type-H vessels (Fig. 6A–D). After HUVECs were treated with or without alcohol for 12 h, in comparison with vehicle, 8.7 mM (0.5%), 52 mM (3%), or 87 mM (5%) alcohol treatment notably decreased the expression of BMP2; 52 mM (3%) alcohol treatment further obviously decreased the expression of BMP2 compared with 8.7 mM (0.5%) alcohol treatment; 87 mM (5%) alcohol treatment further decreased the expression of BMP2 compared with 52 mM (3%) alcohol treatment (Fig. 6E, F).
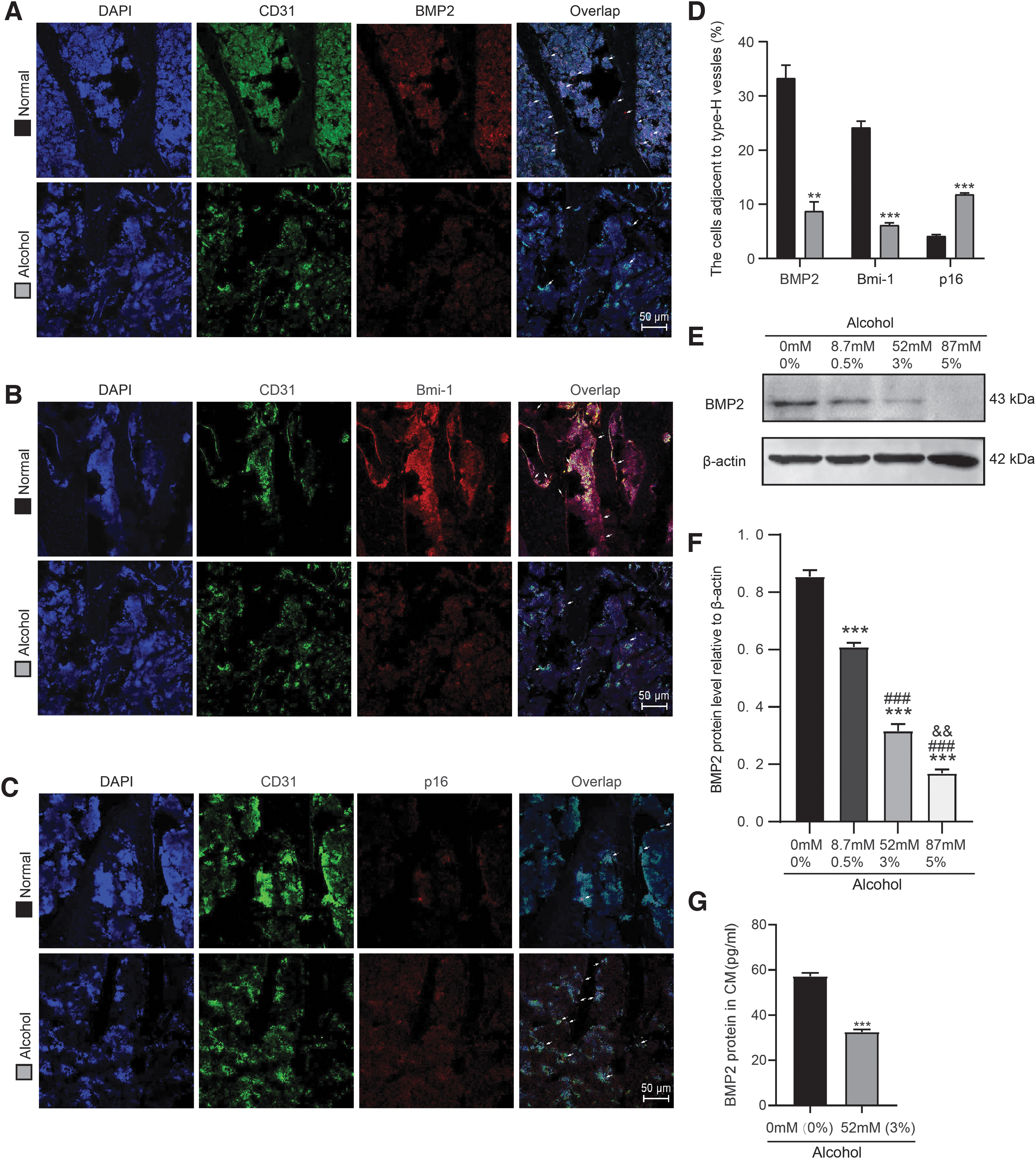
Alcohol reduces the BMP2 secretion of endothelial cells in type-H vessels. The experiments were carried out on the 2-month-old mice fed with normal diet or alcoholic diet for 2 months. Frozen tibial sections from 4-month-old normal- and alcohol-diet mice were prepared and used for immunofluorescence staining.
In addition, after HUVECs were treated with or without alcohol (52 mM; 3%) for 12 h, we collected cell supernatant and concentrated them to CM and analyzed the concentration of BMP2 by ELISA assay. In comparison with vehicle, 52 mM (3%) alcohol treatment decreased the secretion of BMP2 in CM of HUVECs (Fig. 6G). These results suggested that alcohol inhibited cell proliferation, promoted cell aging, and reduced the secretion of BMP2 of endothelial cells in type-H vessels, resulting in the decrease of osteogenic differentiation of BM-MSCs.
Discussion
This study demonstrated that the alcoholic diet retards the bone growth, impairs bone formation of osteoblasts, and injures CD31hiEMCNhi type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells, leading to osteoporosis in mice. Meanwhile, alcohol inhibits the differentiation of BM-MSCs into osteoblasts through reducing the BMP2 secretion of endothelial cells in type-H vessels.
Chronic consumption of excessive alcohol eventually results in an osteopenic skeleton and an increase of risk for osteoporosis [2]. Alcohol-induced bone loss is characterized by damaged bone structure, decreased bone mass, and increased fracture risk after alcohol abuse, which is mainly due to the direct and indirect effects of alcohol [36,37]. Previous study has reported the positive effects of a small amount of alcohol on bones. In detail, light drinking (8 g alcohol per day), mainly wine, was positively correlated with lumbar BMD and whole-body BMD of postmenopausal women [38]. In this study, low-concentrated alcohol had no effect on the proliferation of HUVECs. The CM of low-concentrated alcohol-treated HUVECs had no effect on the osteogenic differentiation of BM-MSCs in vitro. Although the effects of low-concentrated alcohol are controversial, chronic long-term drinking, and binge drinking are harmful to bone tissue [39]. Many studies have confirmed that the animal model of long-term chronic drinking can be established by using Lieber–DeCarli liquid diet, which is reasonable to directly simulate human drinking [40].
It was previously reported that mice showed obvious osteoporotic phenotype under the same feeding conditions as in this article [17]. This modeling method results in much higher ethanol intake than with other approaches and facilitates the comparison with controls by simplifying the pair feeding and is the best procedure available for the study of the toxic effects of alcohol and their interactions with deficiency or excess of various nutrients [41]. In addition, it has been proved that this modeling method can well simulate osteopenia caused by chronic alcohol [17]. Thus, the reasons of the use of this model are to simulate osteopenia caused by chronic alcohol, to maintain higher ethanol intake, and to facilitate the comparison with controls by simplifying the pair feeding. Based on the feed instructions of Lieber–DeCarli liquid alcohol diet, it provides 28% of the calories and is equivalent to 5% alcohol by volume. Consistent with our finding in vivo in this study, we found that when the alcohol concentration in vitro reached 5%, it significantly inhibited the proliferation and BMP2 secretion of endothelial cells and promoted their senescence.
In recent years, the coupling of angiogenesis and osteogenesis has become a new breakthrough in the treatment of osteoporosis [42,43]. Primary osteoporosis is characterized by a decrease in the number of sinuses and arterial capillaries in bone tissue, and the same phenomenon also occurs in postmenopausal osteoporosis [44]. Previous studies found that with aging-related bone loss in mice, the number of type-H vessels decreased, whereas the number of type-L vessels remained unchanged, and type-H vascular endothelial cells also decreased while the whole bone vascular endothelial cell content remained unchanged. In addition, there is a coupling relationship with the content of osteoblasts in mice [6,44]. There is no accumulation of Osterix+ cells around type-L vessels that are not involved in the process of osteogenic differentiation [6], so we have not conducted analysis about type-L vessels.
The number of type-H vessels in the elderly is much less than that in the young, so the type-H vessels can be used as a sensitive indicator of bone aging and bone mass changes [7]. We found that high concentration of alcohol could promote the senescence of endothelial cells, and the aging endothelial cells would further affect the osteogenic differentiation of BM-MSCs, which may provide a new therapeutic target for aging osteoporosis. Previous studies found that through the inhibition of PI3K-Akt signaling pathway, high concentration of alcohol may alter many signaling pathways in HUVECs [45]. This suggests the effects of alcohol on cell proliferation, differentiation, apoptosis, and migration. The reduction of type-H vessels by alcohol plays an important role in alcohol-related bone loss [26]. We found that alcoholic diet inhibited proliferation and promoted aging of endothelial cells.
Cyclin, cyclin-dependent kinases (CDKs), and cyclin-dependent kinase inhibitors play an important role in endothelial cell proliferation and migration [46]. P16 binds to cyclin/CDK complex and causes cell cycle arrest in G1 phase by inhibiting Rb phosphorylation mediated by CDK4/6. Meanwhile, dephosphorylated Rb binds to cell cycle-related transcription factor E2F and blocks its transcription activation domain [47]. In this study, alcohol regulates the proliferation and aging of HUVECs through this signaling pathway. However, further study is needed to explore the crosstalk between vascularization and bone formation in ethanol-induced osteopenia.
Among all the BMPs, BMP2 has a strong osteogenic ability, which significantly increases the expression of osteocalcin. The short-term expression of BMP2 can irreversibly induce bone formation [48]. BMP2 is a key bone growth factor, which can stimulate osteoblast proliferation and induce BM-MSCs to differentiate into osteoblasts by inducing the expression of Runx2, Osterix, and OCN genes. As specific transcription factors, Runx2 and Osterix cooperatively regulate the expression of bone-specific genes, including ALP, type I collagen, and OCN, which induce bone formation [1,48,49]. We found that BMP2 might be a key factor in coupling angiogenesis and osteogenesis. In addition, when bone is injured, the fracture area is surrounded by hematoma. Local hypoxia induces upregulation of BMP2 in endothelial cells to promote osteogenesis. Hypoxia inducible factor-1α (HIF-1α) accumulates in osteoblasts, activating HIF-1α signaling pathway to specifically activate endothelial cells, which leads to vascular endothelial growth factor-A production, further promotes type-H vessel formation and secretion of BMP2, and accelerates bone formation [6,50].
BMPs play an important role in bone formation and fracture healing [51]. Recently, it has been reported that the local and temporary applications of BMP2 stimulate the activation of skeletal stem cells, combined with low level of a colony stimulating factor 1 antagonist inhibits bone resorption, and restores bone regeneration in aged mice [52]. BMP2 is used as an auxiliary drug for fracture healing in clinic [53]. Commercially available BMPs provide a substitute for autologous bone graft for the treatment of tibial nonunion. The recombinant human BMP2 has been widely studied in various preclinical and clinical studies, but the clinical research on nonunion is limited [54]. In addition, some studies have confirmed that alcohol could affect the expression of BMP2 [55]. Our results demonstrated that alcohol inhibited the differentiation of BM-MSCs into osteoblasts through reducing the BMP2 secretion of endothelial cells in type-H vessels. However, whether BMP2 expression is regulated by Bmi-1/p16 signaling pathway in aging endothelial cells of type-H vessels needs further study.
It has been reported that the endothelium can enhance vascular niche function leading to increased hematopoietic stem cell (HSC) numbers [56]. This suggests the effect of type-H vessels on HSCs. Interestingly, there may be a link between osteoblast activity and hematopoietic niches in mouse long bones. Dysfunctional vascular niches induce skeletal aging, bone diseases, and hematological disorders [57,58]. This indicates that the effect of alcohol on bone formation reduction may be determined by alcohol damage to type-H vessels, and then reduce the number of HSCs, resulting in osteopenia. In addition, alcohol exposure can reduce the physiological proliferation rate of bone marrow hematopoietic progenitor cells [59,60]. Thus, alcohol-induced altered hematopoiesis may be a possible determinant in the observed osteopenia. However, the effect of changes in HSCs on type-H vessels has not been reported.
Although BMP2 has been used as an adjuvant in fracture healing [53], this study provides a new idea of targeting BMP2 for preventing alcoholic osteopenia through coupling with angiogenesis and bone formation. Furthermore, this study first reported that Bmi-1/p16 signaling pathway could promote the proliferation and the secretion of BMP2, and delay the senescence of endothelial cells for maintaining bone mass. It provides a new treatment strategy targeting accumulated p16-positive aging endothelial cells in type-H vessel to prevent alcoholic osteopenia.
In conclusion, the alcoholic diet impaired type-H vessel formation through inhibiting proliferation and promoting aging of endothelial cells through Bmi-1/p16 signaling, and curbed the osteogenic differentiation of BM-MSCs through reducing the BMP2 secretion of endothelial cells in type-H vessels. It provides a basis for developing a new treatment strategy targeting aging endothelial cells of type-H vessel to prevent alcoholic osteopenia.
Data Availability Statement
All data and materials used in the analysis are available to any researcher for purposes of reproducing or extending the analysis.
Footnotes
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (81871097 and 81571371 to J.J., 81602839 to X.L. and 81730066 to D.M.), the “Qinglan project” funded by Jiangsu Universities (2020-10 to J.J.), the Natural Science Foundation of Jiangsu Province (BK20151554 to J.J.), the Scientific Research Project of Jiangsu Health Commission (H2019023 to J.L.), and the Xuzhou Science and Technology Project (KC19152 to J.L.).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.