
Abstract
A continuous Bromodeoxyuridine (BrdU) labeling approach was used during the whole process of the mice kidney development to explore the best BrdU-labeling time, the distribution of BrdU-retaining cells, and to probe into the niche of stem cells in adult kidney. BrdU was injected intraperitoneally to the mice once daily for 3 consecutive days from day 11.5 of embryonic period (E11.5) to the postnatal day 21.5 (P21.5). The kidneys were harvested 24 h after the last BrdU injection and 6 months of age. A renal injury model of subtotal nephrectomy (Nx) in adult mice treated with BrdU was used to observe the response of BrdU-retaining cells to renal injury. When BrdU labeled at E11.5-13.5, the BrdU-retaining cells were mainly detected in the papilla and inner medulla in adult mice. When BrdU labeled at P0.5-11.5, the BrdU-retaining cells were mainly detected in the inner medulla and outer medulla. When BrdU labeled at P12.5-17.5, the BrdU-retaining cells were mainly detected in the outer medulla. When BrdU labeled at P18.5-21.5, almost no BrdU-positive cells could be found, except the cortex. Seventy-two hours after Nx operation in adult mice by BrdU-labeling at P0.5-2.5 or P15.5-17.5, a significant increase of BrdU-retaining cells was found in many cortical proximal tubules, while a dramatic decrease was detected in medulla near the incision edge. Moreover, most of BrdU-positive cells were not costained with proliferating cell nuclear antigen. The distributions of label-retaining cells in the mice kidney were different if BrdU was administered in different periods of kidney development. Most of BrdU-retaining cells were quiescent, the proximal tubules were the only segments that always contained BrdU positive cells, which may have the niche of stem cells in adult kidney.
Introduction
Adult stem cells (ASCs) are defined as undifferentiated cells capable of proliferation, self-renewal, production of a large number of differentiated progeny and regeneration of tissue in the adult organism [1]. The potential use of stem cells for regenerative medicine to treat kidney diseases represents a critical clinical goal. Exogenous sources of stem cells, including bone marrow-derived cells and mesenchymal stromal cells, have been proved to have few effects on renal diseases [2,3]. Hence, identification of the existence of ASCs in the kidney is an attractive option for renal repair.
Following acute tubular injury, the kidney undergoes a regenerative response in most cases in renal function recovery [4,5]. Our previous study found that some cells expressing nestin (a neuron stem cell marker) in immature proximal tubule near the incision edge of remnant kidneys might promote renal repair [6]. However, the cell sources for generation are poorly understood.
Over the last decade, numerous studies demonstrated that the kidney had intrinsic stem/progenitor cells, which could proliferate and differentiate into new renal cells. Renal stem cells might exist in specific parts of the adult kidney, such as renal papilla [7], tubular epithelial cells [8,9], Bowman's capsule [10], and the S3 segment of the proximal tubules [11,12].However, whether kidneys harbor similar stem cells, where these cells are located, and whether they substantially contribute to renal regeneration still remain unknown.
Traditionally, the expressions of cell specific markers were used for identification of stem cell. However, the specific markers for renal stem/progenitor cells have not yet been confirmed. Most of studies chose the markers expressed in other ASCs (such as hematopoietic stem cells, mesenchymal stem cells) or in developing kidney, while the distributions of renal stem cells in the adult kidney were different and controversial in previous reports [10,13,14]. In addition, the expressions of these markers may change under pathophysiological conditions. We found that the proximal tubule cells might reexpress nestin near the incision edge of remnant kidneys [6]. Therefore, assessment of the cell origin and fate using cell specific markers have been restricted to indirect and retrospective analysis.
The technique of the label-retaining cell (LRC) approach [3] is a more powerful tool to identify ASCs and their distributions in the stem cell niche, which takes advantage of their slow-cycling, quiescent nature. Bromodeoxyuridine (BrdU), as a thymidine analog, is a marker of DNA synthesis, which can be incorporated into the newly synthesized DNA of dividing cells during the S-phase of the cell cycle and detected by using BrdU specific antibodies [15]. Using this method, LRCs have now been reported in many adult tissues and organs, including the lung [16], cornea [17], intestine [18], prostate [19], hair follicles [20].
With BrdU labeling, the existence of LRCs in the adult kidneys has been reported previously. Maeshima et al. [8] found that the LRCs were scattering in the proximal, distal, and collecting tubules of normal rat kidneys. Wang et al. [21] revealed that LRCs existed in the glomeruli expressed undifferentiated podocyte and endothelial markers at high rates, while those in the renal tubules expressed nestin and vascular markers at low rates at 6-week time point. Other studies [22] showed the LRCs presented in the interstitium of kidneys that were capable of reexpressing the mesenchymal cell markers vimentin and e-cadherin after ureteral obstruction. The renal papilla has also been proposed as a niche for adult kidney stem cells because of the presence of large numbers of BrdU-retaining cells in the papillary interstitium in rats and mice [7].
BrdU-retaining cells were identified to be quiescent throughout the life of animals. After ischemia injury, these slow-cycling cells may proliferate and participate in renal regeneration [7]. It is worth noting that although all the above-mentioned studies used the BrdU labeling method, their results were completely different. The reasons might be related to the diverse times of BrdU administration in the different developmental periods of nephron segments in each study, and the different observation times after BrdU injections.
The kidney development of mice starts from embryonic day 11 (E11) to postnatal day 21 (P21) [10].The human kidney continues to develop until no new nephrons appeared after 36 weeks of gestation [10]. Since the different segments of nephron are mature at different stages of kidney development, it may lead to inconsistent findings in previous studies if different BrdU-labeling times were used. In the present study, a continuous BrdU-labeling approach was used during the whole process of the mice kidney development to explore the best BrdU-labeling time, the distribution of BrdU-retaining cells, and to probe into the niche of stem cells in adult kidney.
Materials and Methods
Animals
Twelve-week-old female C57BL/6J mice were mated with the same strain of male, in accordance with the ratio of 1:1. If the vaginal plug was observed next day, it was denoted embryonic day 0.5 (E0.5) [23]. The offsprings were chosen as observed objects and their birth day was denoted as postnatal day 0.5 (P0.5).
The animals used in the study were approved by Animal Welfare and Ethics Group, Department of Experimental Animals, Fudan University.
BrdU labeling
BrdU labeling is based on the premise that stem cells segregate DNA asymmetrically, with the older strand being retained by stem cells, while the new strand is passed on to daughter cells [15]. Since the kidney development of mice starts from day 11.5 of embryonic period (E11.5) to the postnatal day 21.5 (P21.5) [24], we chose the BrdU labeling period covering the whole process of the kidney development (Fig. 1). Fifty mg/kg body weight BrdU (Sigma-Aldrich) were injected intraperitoneally to the mice once daily for 3 consecutive days at E11.5, P0.5, P3.5, P9.5, P12.5, P15.5, and P18.5.

Experimental methods.
In E11.5 group, the BrdU was injected in pregnant female mice from E11.5 to E13.5, then the offspring mice kidneys were harvested at P0.5 and 6 months of age. Other groups of mice were treated with BrdU at different stages after the birth, and their kidneys were harvested in the developmental stage (24-h after the last BrdU injection) and adult stage (6 months of age). There were 5 mice in each observation group.
Subtotal nephrectomy
To observe the response of BrdU-retaining cells to renal injury, we induced a renal injury model of subtotal nephrectomy in adult (6 months old) mice treated with BrdU at P 0.5-2.5 or P15.5-17.5 (Fig. 2). In brief, the upper pole of one side kidney was resected under anesthesia, leaving two-thirds kidney with the renal parenchyma and hilum intact, and resulting in no injury to the inner medulla and papilla. The lower pole of the kidney without injury was used as the control pole. Kidneys were harvested by 72 h after operation.
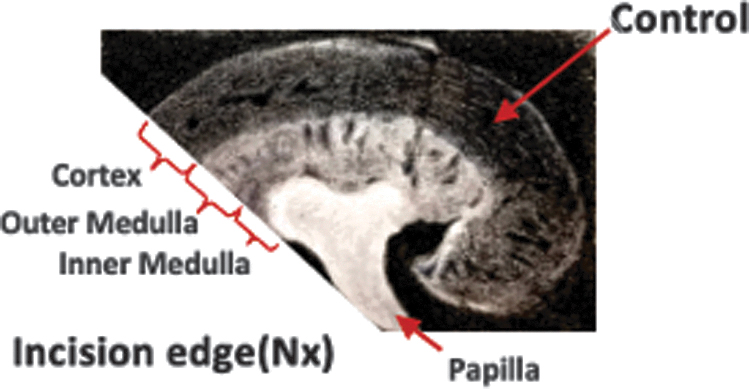
Subtotal nephrectomy.
Immunofluorescence
The kidney tissues were fixed in 4% paraformaldehyde for 6 h, incubated with 30% sucrose overnight, and frozen in optimal cutting temperature compound. The tissue sections (5 μm) were air dried for immunofluorescence assay.
BrdU detection requires a pretreatment of the sections to denature DNA. Sections were incubated with 2 N HCl at 37°C for 30 min, then buffered with 0.1 M sodium borate (pH 8.5), followed by incubation in PBS.
After DNA denaturation, sections were blocked with 10% normal donkey serum for 20 min. A blocking buffer from the M.O.M kit (Vector Laboratories) was used if the primary antibody was anti-mouse. Sections were then incubated with primary antibodies for 60 min. After washing, sections were incubated in Cy2- or Cy3-conjugated anti-IgG secondary antibody (Jackson Immunoresearch Laboratories) for 30 min.
To determine the relationship between intrinsic renal cells and BrdU-positive cells, double immunostaining with BrdU and several markers expressed in different regions of mature kidney were used, such as Lotus tetragonolobus agglutinin (LTA), a marker for both proximal tubule and collecting duct; dolichos biflorus agglutinin (DBA), a marker of collecting ducts; Tamm Horsfall glycoprotein (THP), a marker of thick ascending limb; aquaporin-1 (AQP1), a marker of matured proximal tubule; aquaporin-2 (AQP2), a marker of matured collecting ducts; and thiazide-sensitive cotransporter (TSC), a marker of distal convoluted tubules.
The following antibodies were used: (1) anti-BrdU (1:100; Abcam); (2) FITC-labeled LTA (1:200; Vector Laboratories, Burlingame, CA); (3) FITC-labeled DBA (1:200; Vector Laboratories); (4) anti-AQP1 (1:100; Chemicon International, Temecula, CA); (5) anti-AQP-2 (1:200; Alpha Diagnostic International, San Antonio, TX); (6) anti-THP (1:500; Organon-Technika, Durham, NC); (7) anti- TSC (1:200, Mark Knepper; NIH, Bethesda, MD); and (8) antiproliferating cell nuclear antigen (PCNA) rabbit polyclonal antibody (1:100; Santa Cruz Biotechnology, Inc.).
Sections were viewed and imaged with a Zeiss Axioskop and spot-cam digital camera (Diagnostic Instruments) or confocal microscopy (Zeiss; LSM510). Final images were processed with Adobe Photoshop software.
Quantification of BrdU/PCNA-positive cells
Quantitative analysis of BrdU/PCNA-positive cells was performed by counting the positive nuclei from 10 randomly selected fields ( × 400) under a fluorescence microscope by an experienced pathologist.
Statistical analyses
All values are expressed as mean ± SEM and in box diagrams. Data analysis was performed using GraphPad Prism 7.0 using the unpaired Student's t test or ANOVA as appropriate. Statistical significance was defined as P < 0.05.
Results
Distribution of BrdU-positive cells in the developmental kidney and adult kidney
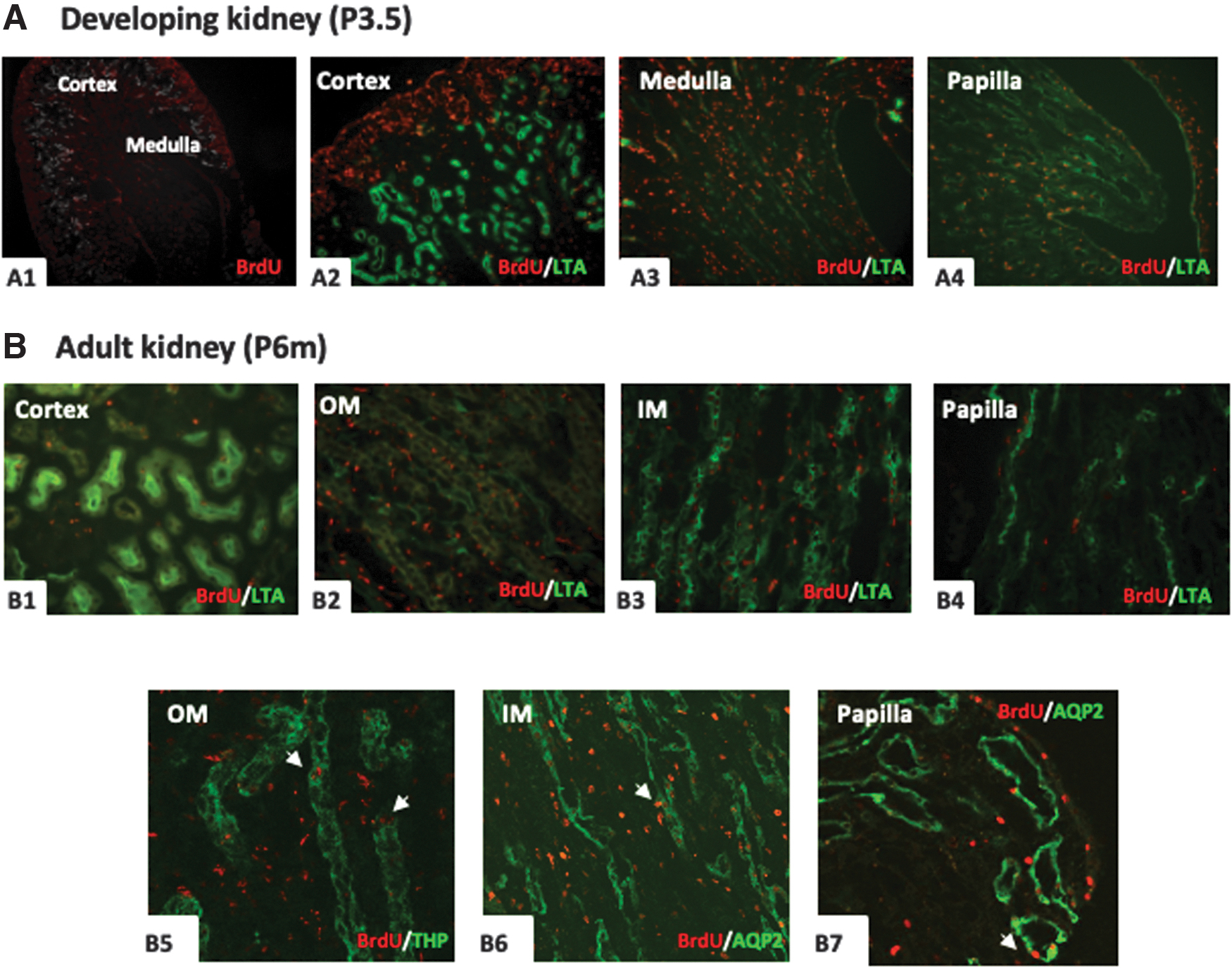
When a pulse of BrdU was administered in pregnant female mice at E11.5-E13.5, lots of BrdU-positive cells were found in the offspring mice kidney of P 0.5, especially in the outer stripe of cortex, medulla and papilla (Fig. 3A1–A4). Double immunostaining results confirmed that some BrdU-positive cells was located in LTA, THP, and DBA-positive tubules, namely proximal tubule, collecting duct, and thick ascending limb, as well as in the interstitial (Fig. 3A5-7).

BrdU immunoreactivity in the P0.5 kidney and adult kidney obtained from mice treated with BrdU at E11.5-13.5.
Compared with newborn mice, the numbers of BrdU-positive cells were significantly reduced in the adult offspring mice kidney and mainly detected in the inner medulla and papilla (Fig. 3B2-4). The BrdU-positive cells were rare in the cortex and only distributed in the LTA- and AQP1-positive tubules, but not in THP- and TSC-positive tubules (Fig. 3B1, 5-7). None of BrdU-positive cells was costained with proliferative marker PCNA (Fig. 3B8).
With BrdU administration at P 0.5-P 2.5 of newborn mice, numerous BrdU-positive cells were detected in the subcapsular cortex, medulla, and papilla of developmental kidneys derived at P3.5, which could be identified by LTA costaining (Fig. 4A1-4). When the mice grew to adulthood, the numbers of BrdU-positive cells were significantly decreased in the kidney. The immunoreactivity of BrdU was weak in the cortex and scattered in the glomeruli, proximal tubules, and interstitial (Fig. 4B1). However, many BrdU-positive cells were still detected in the outer medulla and inner medulla, but few in the papilla (Fig. 4B2-4). Some BrdU-positive cells were located in the AQP2-positive tubules of inner medulla and THP-positive tubules of outer medulla (Fig. 4B5-7).
BrdU immunoreactivity in the P3.5 kidney and adult kidney obtained from mice treated with BrdU at P0.5-2.5.
With BrdU administration at P3.5-P5.5 of newborn mice, the distribution of BrdU-positive cells in the developmental kidney of P6.5 was similar with the P3.5 kidney with BrdU treatment at P0.5-2.5, mainly in the subcapsular cortex, outer medulla, and inner medulla (Fig. 5A1a-1c). However, the patterns of BrdU expression were quite different in the adult kidneys of these two groups. The total numbers of BrdU-positive cells were significantly decreased in P3.5-P5.5 group compared to P0.5-2.5 group. Only a few BrdU-retaining cells could be found in the proximal tubules of cortex, interstitium of outer medulla, and relatively more in the collecting ducts and interstitium of inner medullar (Figure 5A2a-2c).
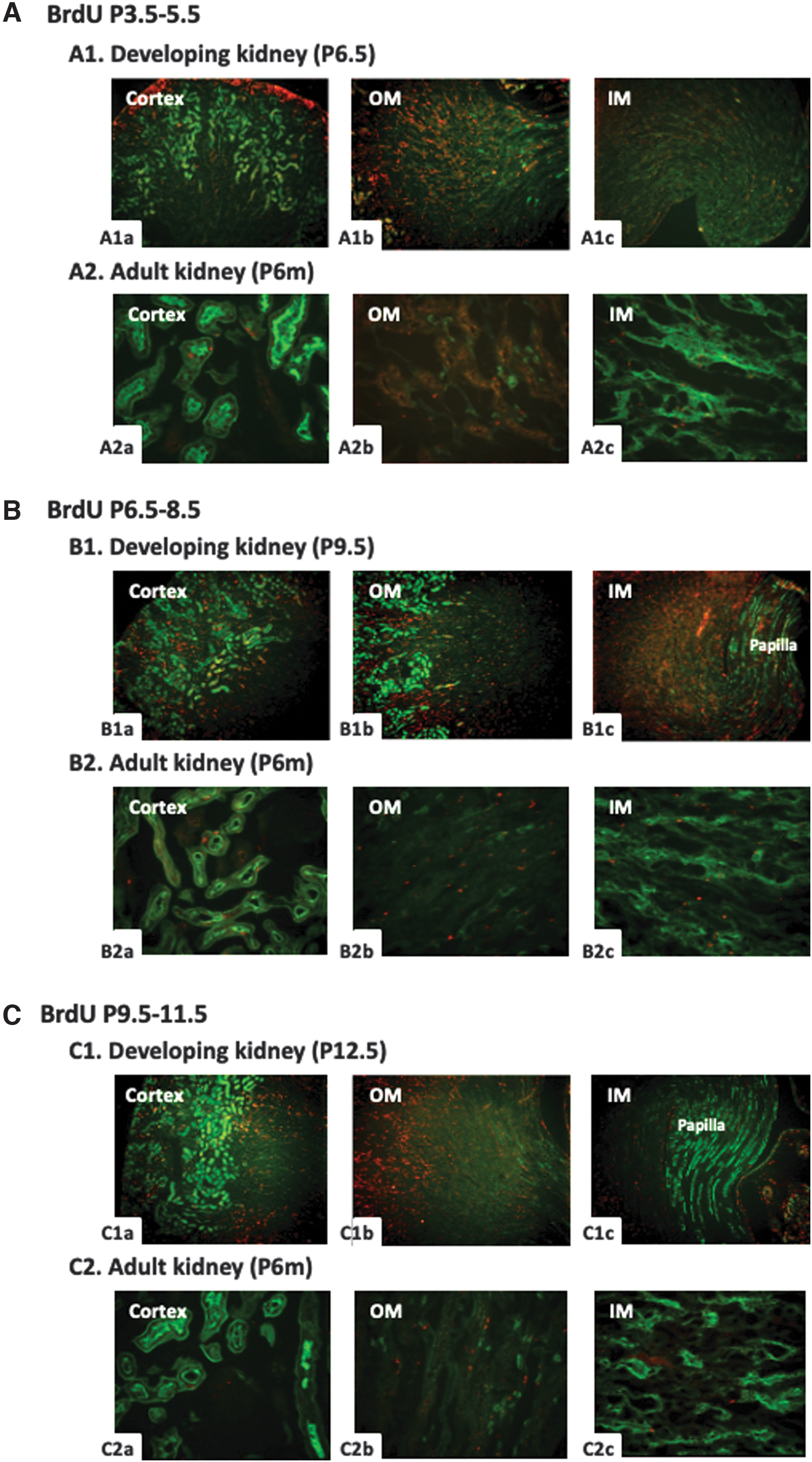
BrdU (red) and Lectin-LTA (green) costaining in the kidney of mice treated with BrdU at P3.5-5.5, P6.5-8.5, and P9.5-11.5.
With BrdU administration at P6.5-P8.5, lots of BrdU-positive cells were observed in the cortex, outer medulla, and inner medulla of developmental kidney at P9.5, too. The BrdU-staining cells were uniformly distributed in the cortex, no longer concentrated in the outer stripe of cortex (Fig. 5B1a-1c). In the adult mice kidney, only a small number of BrdU-positive cells were detected in the proximal tubules of cortex and a little more in the interstitium of outer medulla and inner medulla (Fig. 5B2a-2c), which was similar to the patterns of P3.5-P5.5 group.
With BrdU administration at P9.5-P11.5, a large number of BrdU-positive cells were detected in the outer medulla, and a relatively less in the cortex and inner medulla of developmental kidney at P12.5. No BrdU was found in the papilla (Fig. 5C1a-1c). When the mice grew to 6 months old, their kidney still retained some BrdU-positive cells in the outer medulla, few in the cortical proximal tubules, and almost none in the inner medulla (Fig. 5C2a-2c).
With BrdU administration at P12.5-P14.5 or P15.5-P17.5, the distribution patterns of BrdU-positive cells were similar, both in the developmental kidney and in the adult kidney, but different from other BrdU-treated groups. In the developmental kidney at P15.5 and P18.5, there was a large number of BrdU-positive cells in the outer medulla (Fig. 6A1b, B1b), and some in the cortex (Fig. 6A1a, B1a), but rare in the inner medulla and papilla.
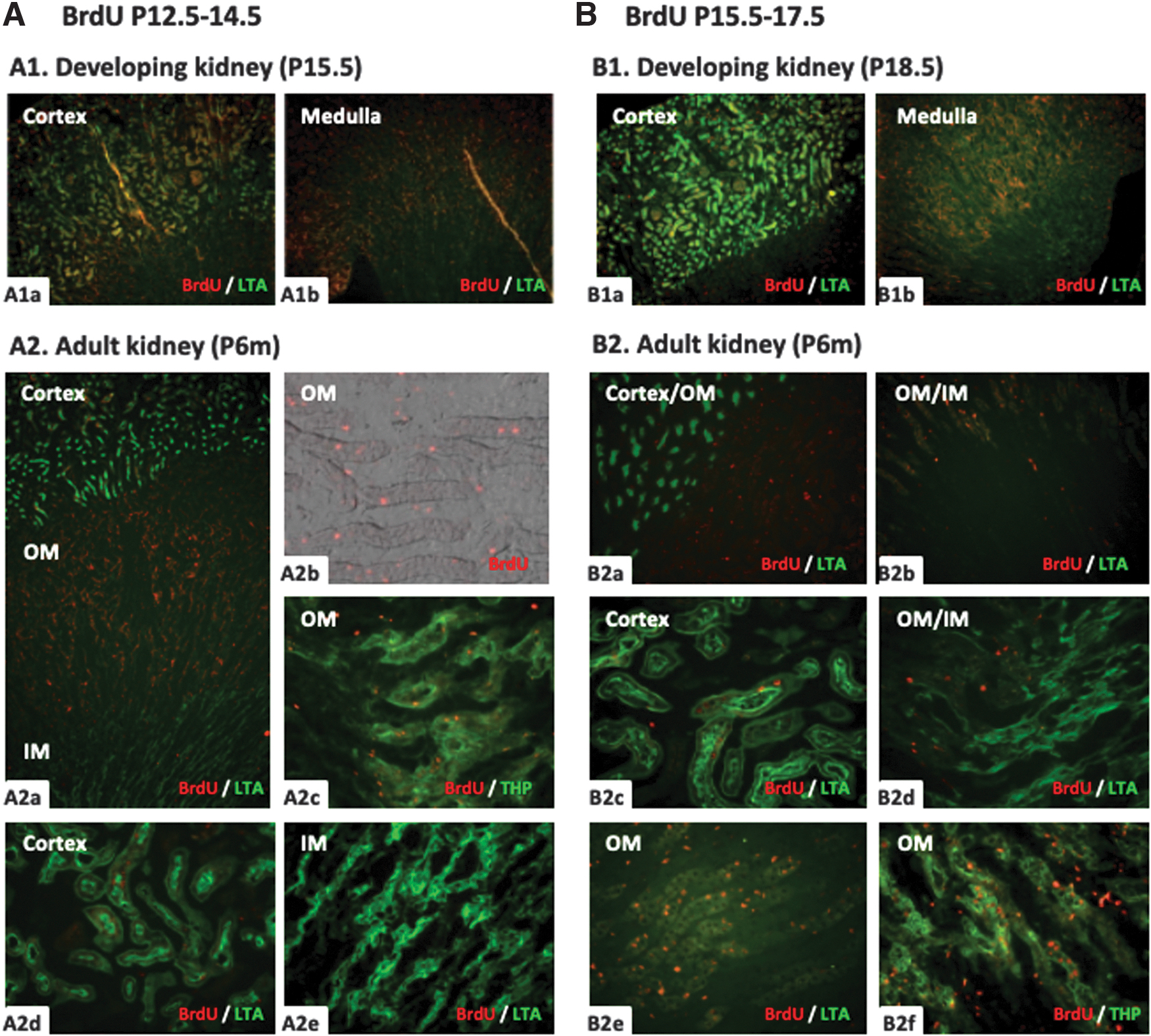
BrdU immunoreactivity in the developing and adult kidney of mice treated with BrdU at P12.5-14.5 and 15.5-17.5. On the next day after BrdU administration at P12.5-14.5 or P15.5-17.5, the distribution of BrdU-positive cells was very similar in the mice kidney of P15.5 and P18.5. Plenty of positive cells (red) were detected in the outer medulla
In the adult kidney of these two groups, the number of BrdU-positive cells in the outer medulla decreased significantly and more than 50% of BrdU-positive cells were located in the thick ascending limb costained with THP, the others located in the interstitium (Fig. 6A2a-2c, B2d-2f). Similarly, only few BrdU-retaining cells were detected in the cortical proximal tubules of adult mice costained with LTA (Fig. 6A2d, B2c). None of the BrdU-positive cells was found in the inner medulla (Fig. 6A2e, B2d).
With BrdU administration at P18.5-P20.5, the last 3 days of mice kidney development, only a small amount of BrdU-positive cells were found in the cortex and outer medulla but not in the inner medulla at P21.5 (Fig. 7A1a-1c). In the 6-month adult kidney, few BrdU-positive cells were still detected in the cortex, and most of them located in the proximal tubules. None of BrdU-retaining cells was found in the outer medulla or inner medulla (Fig. 7A2a-2c).
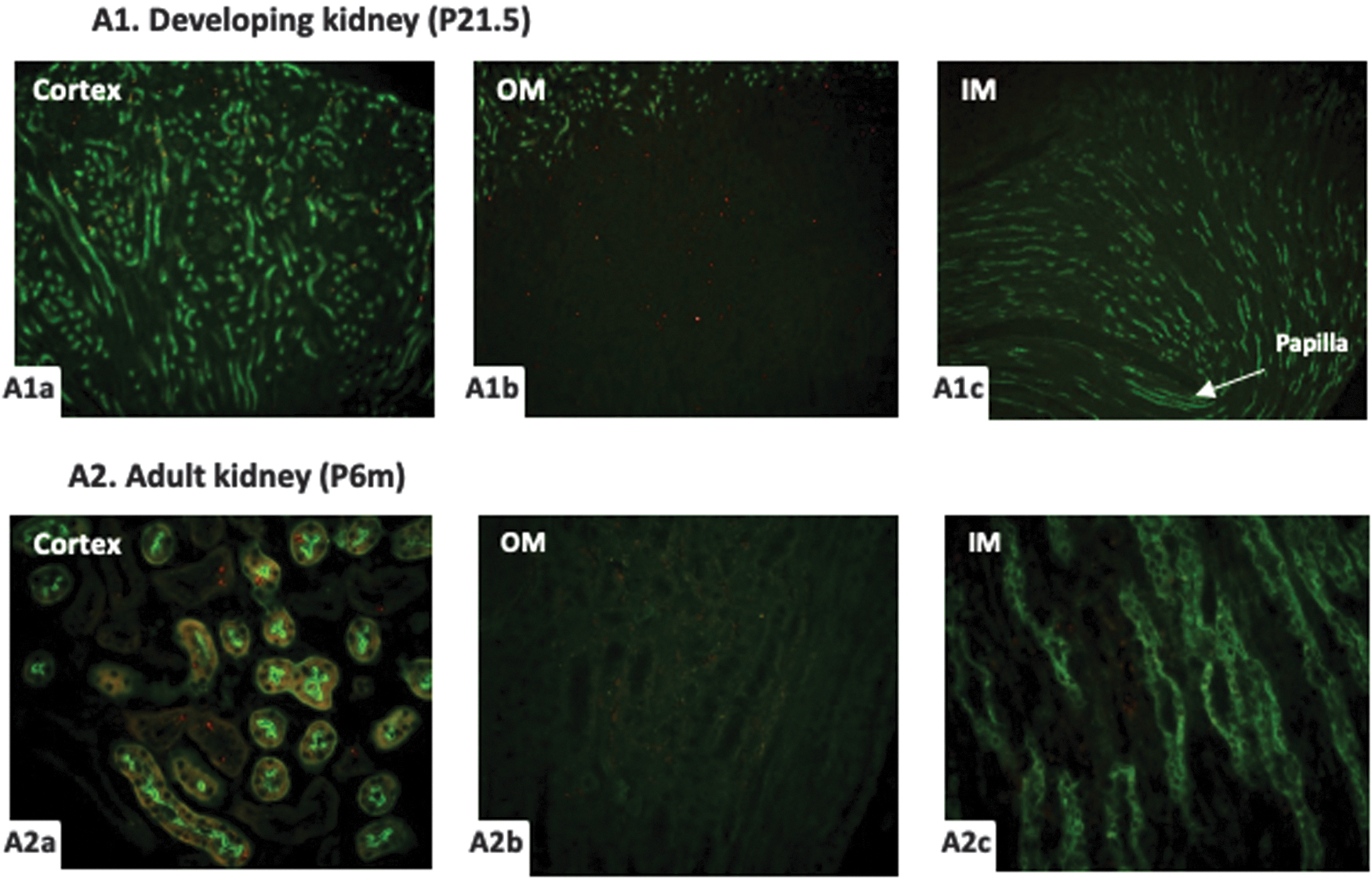
BrdU (red) and Lectin-LTA (green) costaining in the kidney of mice treated with BrdU at P18.5-20.5. A small number of BrdU-positive cells were detected in the cortex (
Heterogeneous distribution of the LRCs in the adult kidney
To further clarify the distribution characteristics of BrdU-retaining cells in the adult mice kidney with BrdU labeling at different developmental periods, we quantitatively analyzed the BrdU-positive cells in the adult kidney. Table 1 shows that the ability to retain the BrdU in different regions of adult kidney was correlated significantly with BrdU-labeling time during kidney development. When BrdU labeled at E11.5-13.5, the BrdU-retaining cells were mainly detected in the papilla and inner medulla. When BrdU labeled at P0.5-11.5, the BrdU-retaining cells were mainly detected in the inner medulla and outer medulla. When BrdU labeled at P12.5-17.5, the BrdU-retaining cells were mainly detected in the outer medulla. When BrdU labeled at P18.5-21.5, almost no BrdU-positive cells could be found in the adult kidney, except the cortex.
The Average Number of Bromodeoxyuridine-Positive Cells Under High-Power ( × 400) Field of Fluorescent Microscopy
PT, proximal tubule; TAL, thick ascending limb of the Henle’ loop; OMCD, outer medullary collecting duct; IMCD, inner medullary collecting duct.
The changes in the distribution of BrdU-positive cells due to different labeling time strongly suggested that such a large number of BrdU-retaining cells detected in the adult kidney might be quiescent cells after full development of these regions. However, the very interesting finding in Table 1 was that a small number of LRCs were always been detected in the renal cortex whenever BrdU was administered at embryo stage or after birth, without numerous cells gathering, suggesting that renal cortex might be always in an exuberant proliferating phase during the whole development stage, in which proximal tubules might be the only nephron segments stably containing a few BrdU-retaining cells after a growing period of 6 months.
BrdU-positive cells response to renal injury
To identify whether the large number of BrdU-positive cells retained in the outer and inner medulla of adult kidney by BrdU-labeling at P0.5-2.5 were quiescent cells, we performed a subtotal nephrectomy (Nx) in these adult mice. Seventy-two hours after operation, the total numbers of BrdU-positive cells were markedly increased in the cortex, and the immunoreactivity near the incision edge enhanced more significantly than non-Nx side of same kidney (Fig. 8A1,2,8). BrdU-staining cells were mainly distributed in the proximal tubules and even presented a string arrangement in some tubules at the site of injury, only a small amount was detected in the glomeruli, vessels, interstitium and other tubular structures. More interestingly, some BrdU-positive cells did not express PCNA, even though numerous PCNA-positive cells were found near the incision edge of kidney (Fig. 8A1,2). In contrast, originally retained abundant BrdU-positive cells in the medulla significantly reduced and disappeared rapidly in the vicinity of incision, while PCNA-staining cells significantly increased, compared to non-Nx side of kidney (Fig. 8A3-6, A8). There was no change of BrdU-staining in the papilla, which was not damaged during nephrectomy (Fig. 8A

BrdU (red) and PCNA (green) costaining in the Nx-kidney of adult mice treated with BrdU at P0.5-2.5 and P15.5-17.5.
In addition, we performed subtotal nephrectomy in another group of 6-month-old mice with a large number of BrdU-positive cells retained in the outer medulla induced by BrdU-labeling at P15.5-17.5. The results were quite similar with the previous one. Although the number of PCNA-positive cells in the tubules and other structures near the incision edge increased significantly compared with those in the control non-Nx pole of the same kidney, the changes of BrdU-positive cells were completely different. A significant increase of BrdU-retaining cells was found in many cortical proximal tubules, while a dramatic decrease was detected in the TAL and interstitium of outer medulla near the incision edge (Fig. 8 B1-4,6). Moreover, most of BrdU-positive cells were not costained with PCNA. None of the BrdU-positive cells was found in the papilla (Fig. 8B5).
Discussion
BrdU labeling, which is to search for slow-cycling cells by labeling their DNA, is one of the most common methods to identify stem cells. A pulse of BrdU mostly labels transit-amplifying (TA) cells, while long-term BrdU labeling is thought to mark stem cells that retain the label for an extended period as a result of slow turnover. Adequate labeling intensity and a suitable washout period for the TA and terminal differentiation compartments will thus result in so-called LRCs, which are believed to represent the stem cell compartment.
The innovation of our study is that a continuous BrdU-labeling approach was used during the whole process of the mice kidney development, and two time-points after BrdU administration were observed to explore the best BrdU-labeling time, the distribution of BrdU-retaining cells, and to probe into the niche of stem cells in adult kidney. One was a short time (24 h) after BrdU injection, to localize the rapidly proliferating TA cells, which indicated the active site of kidney development. Another was a long chase period of 6 months, to identify renal stem cells using the slow-cycling nature of stem cells. In addition, a subtotal nephrectomy was performed to identify whether the large number of BrdU-retaining cells in adult kidney were quiescent cells or stem cells.
The adult kidney is given rise from the metanephros, which derives from the inductive interactions between the metanephric mesenchyme (MM) and the ureteric bud (UB) [24,25]. The UB is first visible as an outgrowth at the distal end of the Wolffian duct at ∼5 weeks of gestation in humans or 10.5 days postembryonic mice [24]. The MM becomes histologically distinct from the surrounding mesenchyme and UB invade into the MM at 11.5 days postconception in mice and 5 weeks in humans [10]. Signals from the MM cause the UB to branch into a T-tubule and then to undergo dichotomous branching, giving rise to the urinary collecting system and all of the collecting ducts.
Simultaneously, reciprocal signals from the UB induce the MM to condense along the surface of the bud. Then, the MM undergo mesenchymal-to-epithelial conversion to generate all the different types of nephron epithelia, including Bowman's capsule, podocytes, and the proximal and distal tubule cells [26,27].
Overviewing the results of BrdU labeling in short time, we found that the distribution of BrdU-labeling proliferating site was consistent with the characteristics of kidney development. After the first few rounds of branching of the UB derivatives, and the concomitant induction of nephrons from the mesenchyme, the kidney begins to become divided between an outer cortical region where nephrons are being induced, and an inner medullary region where the collecting system will form [26]. As growth continues, successive groups of nephrons are induced at the peripheral regions of the kidney, known as the nephrogenic zone.
Thus, within the developing kidney, the most mature nephrons are found in the innermost layers of the cortex, and the most immature nephrons in the most peripheral regions [26]. Our results indicated that most of the BrdU-positive cells found at this time point were proliferating cells, but it was possible to cover stem cells.
After a 6-month chase period, the adult kidney still contained large numbers of BrdU-retaining cells. We know that the distributions of LRCs were heterogeneous at different periods of BrdU administration. BrdU incorporates only during the S phase of cell cycle, using labeling the proliferating cells. One possibility for BrdU-retaining cells was that these cells were just during S phase of cell cycle and no mitosis, maintaining the resting state for a long time and retention of the BrdU, because the time point we marked happened to be the time period when the development of the kidney segment at this site was completed. Moreover, cells in the renal medulla and papilla were renewed very slowly under physiological conditions, so they could remain positive.
Another possibility was the existence of stem cells, which had the nature of slow-cycling and asymmetric division that BrdU would not be diluted in offspring cells, and could be labeled whenever BrdU was administered. In our study, we found that the proximal tubule was the only segment stably containing few BrdU-retaining cells. Thus, we speculated that the niche of kidney stem cells was most likely existed in the proximal tubule.
We performed a subtotal nephrectomy to distinguish whether the BrdU-retaining cells in adult kidney were quiescent cells or stem cells. PCNA-labeled nuclei identify a population of cells in late G1 and early S-phase of cell cycle [28], also expressed during DNA repair and the protein can increase the extent of nucleotide excision by patenting the catalytic function of the excision repair enzyme [29]. We found that BrdU-positive cells significantly reduced and disappeared rapidly in the vicinity of incision in the outer medulla and inner medulla, while PCNA-staining cells significantly increased, compared to non-Nx side of kidney. These data suggested most BrdU-retaining cells in medulla were quiescent and rapidly entered into the cell cycle during renal injury resulting in the BrdU dilution.
In the Nx side of the kidney, the cells with BrdU retaining and PCNA expression were mainly distributed in the proximal tubules at the site of injury. It was interesting that BrdU-positive cells increases after nephrectomy, which might be due to the reuse of BrdU of new proliferating cells after the injury or death of old BrdU-positive cells. We noticed that some BrdU-positive cells were without PCNA expression, which might be due to the different expression periods between BrdU and PCNA during the cell cycle, or some BrdU-retaining progenitor cells were not activated during injury. It suggested that renal progenitor cells might exist in the cortex of the kidney especially the proximal tubules, which was consistent with our previous6 and other results8,29.
Following tubular injury, the kidney undergoes a regenerative response leading in most cases to recover renal function. The cell source for regenerating cells is poorly understood. Humphreys et al. [30] performed lineage tracing studies by using Six2-GFPCre transgenic mice and excluded the contribution of stem cells coming from the interstitium or from the bone marrow to tubular regeneration in rodents. To date, several studies have suggested the existence of renal progenitor-like tubular cells [8,31], suggesting the local renal repair process.
Our study also suggested that BrdU-positive cells increased only in the proximal tubules of the injured renal cortex, while BrdU-positive cells decreased in the medulla after injury. It indicated that the kidney repair might be a local process without the migration of precursor cells, or the stem cells might not have time to migrate because our observation time were only 72 h.
Conclusions
Our results suggested that the distributions of LRCs in the mice kidney were different if BrdU was administered in different periods of kidney development. Most of BrdU-retaining cells were quiescent, the proximal tubules were the only segments that always contained BrdU-positive cells, no matter when the BrdU was administrated. These results demonstrated that the proximal tubules may have the niche of stem cells in adult kidney.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Footnotes
Acknowledgment
We thank Dr. Haowen Jiang for drawing the illustration of a mouse kidney receiving subtotal nephrectomy.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by grants from National Key R&D Program of China (2020YFC2005000), Key Project of National Natural Science Foundation of China (No. 81730017), Shanghai Medical leading talents fund (No. 2019LJ03), Innovation action plan project of Shanghai Science and Technology Commission (No. 17411950700, 20Y11904500, and 21ZR1411100).